Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва)
Активность нейронов коры головного мозга у голодных животных
Ю. А. ФАДЕЕВ (Москва)
Успехи, современной нейрофизиологии, связанные с использованием новых тонких методов электрофизиологического исследования мозга, позволили подойти к изучению центральных механизмов мотивационных состояний животного и человека. Эти состояния организма являются необходимым компонентом афферентного синтеза при формировании целенаправленной поведенческой реакции животного (П. К. Анохин) (13). Известно, например, что такая поведенческая реакция как пищедобывательная, полноценно развертывается только в том случае, если животное длительное время было лишено пищи, т. е. голодало. Следовательно, голодное состояние организма является важным побуждающим моментом в формировании целенаправленного пищевого поведения. Выяснение механизмов, по которым на основе этого внутреннего состояния животного складывается тот нервный аппарат, в дальнейшем приводящий животного к конечной цели (получение пищи), имеет несомненно практическое значение.
В настоящее время установлено, что пищевое возбуждение у голодных животных на основе первичного возбуждения пищевых центров гипоталамуса и его восходящих активирующих, влияний избирательно распространяется на кору головного мозга, в частности адресуется к ее передним отделам (К. В. Судаков) (6). Однако остается невыясненным вопрос о том, как восходящие активирующие влияния гипоталамуса у голодных животных изменяют деятельность корковых нейронов. Ответив на этот вопрос, можно ближе подойти к пониманию конкретных механизмов афферентного синтеза в пищедобывательной деятельности.
Исходя из всего вышеизложенного, в настоящем исследовании мы изучали активность нейронов коры головного мозга у голодных животных.
Опыты проводились под уретановым наркозом. Голодное состояние животного, находящегося под уретановым наркозом, характеризовалось повышенной биоэлектрической активностью передних отделов коры головного мозга, что выражалось внешне высокочастотными низкоамплитудными колебаниями биопотенциалов. Такая избирательная ЭЭГ активация передних отделов коры головного мозга исчезала при искусственном насыщении животного, т. е. после введения молока в желудок и инъекции глюкозы в кровь (5). Применив микроэлектродную технику регистрации активности одиночных нейронов коры мозга, мы решили установить, как отражается состояние голода и насыщения в деятельности нейронов коры головного мозга.
Активность нейронов регистрировалась в передних отделах коры мозга внеклеточно с помощью стеклянных микроэлектродов. Экспериментальные данные были получены на 31 кошке.
В первой серии опытов у голодных и накормленных животных сравнивалась активность нейронов передних отделов коры головного мозга. При погружении микроэлектрода в передние отделы коры мозга голодной кошки (2-суточное голодание) мы обнаружили значительное количество нейронов, имеющих фоновую импульсную активность. Рисунок фоновой активности нейронов очень разнообразен, а частота возникновения импульсаций в нейронах лежала в пределах от 8 до 25 в секунду. В некоторых случаях частота разрядов нейронов была очень низкой (3—4 импульса в секунду) или очень высокой (40—50 импульсов в секунду). Нейроны с исходной фоновой активностью обычно встречались по всей глубине коры головного мозга. Их активность продолжалась 5—10 минут, иногда до 20 минут. У накормленных до опыта животных под уретановым наркозом в передних отделах коры мозга нейроны с исходной фоновой активностью встречались в значительно меньшем количестве, чем у голодного животного. Их активность длилась очень короткое время (0,5—1 минута). Все это указывало на то, что у голодных животных исходная фоновая активность нейронов передних отделов коры мозга выражена более отчетливо, чем у накормленных.
Для сравнения степени активации нейронов передних отделов коры головного мозга у голодных -животных мы подсчитывали (количество нейронов с фоновой активностью, встречающихся по мере прохождения микроэлектрода -через всю толщину коры. В результате нами было установлено, что у голодных животных в передних отделах коры -мозга встречалось в среднем 7—8 нейронов с исходной фоновой активностью на одно прохождение микроэлектрода через все слои коры. У накормленных же животных в передних отделах коры мозга встречалось значительно меньше таких нейронов, в среднем 3 нейрона на одно прохождение микроэлектрода через все слои коры.
Найденное нами различие в уровне активности нейронов передних отделов коры головного мозга у голодных и накормленных животных позволило нам предположить, что у первых выраженная фоновая активность нейронов была связана с исходным голодным состоянием. С целью проверки этого предположения мы исследовали реакции нейронов передних отделов коры мозга при определенном воздействии на животное, приводящем к снижению уровня его «голодного» возбуждения. В качестве такого воздействия мы применили введение в кровь голодному животному раствора глюкозы. Обычно применялся 40% раствор, который вводился в количестве 2 см3 как внутриартериально (a. carotis), так и внутривенно (v. femoralis).
Всего в 2-8 опытах на голодных животных мы исследовали 121 нейрон. Было выделено 5 типов реакций нейронов на введение в кровь глюкозы (табл. 1).
Таблица 1
Соотношение типов нейронов коры мозга по ответной реакции на введение глюкозы
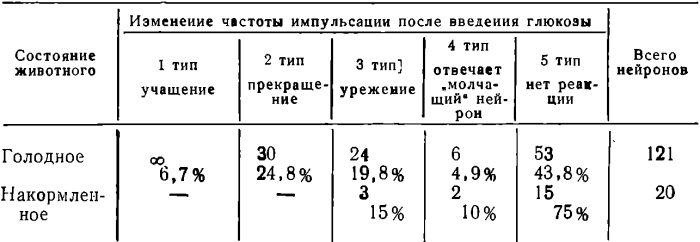
Как видно из таблицы, большая труппа нейронов передних отделов коры головного мозга в ответ на введение глюкозы снижала частоту своих разрядов, иногда вплоть до прекращения активности (2 и 3 типы, 44,6%). Некоторые нейроны после введения глюкозы наоборот увеличивали частоту разрядов (6,7%).
Кроме того, в передних отделах коры головного мозга голодных животных мы обнаружили так называемые «молчащие» нейроны, т. е. нейроны, не имеющие исходной фоновой активности, которые, однако, активировались во время или после введения глюкозы (4,9%). Наконец, имеется также и большая труппа нейронов, которая в ответ на введение в кровь глюкозы не изменяла своей исходной фоновой активности (43,8%). Подобное распределение типов реакций нейронов на введение глюкозы является статистически достоверным (Р = 0,04).

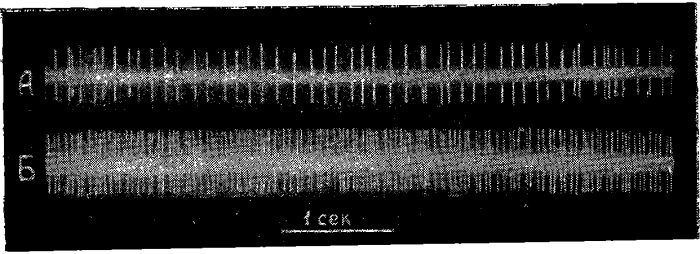
Рис. 1. Нейрон с исходной фоновой активностью из передних отделов коры мозга голодной кошки (уретановый наркоз).
А — активность нейрона до введения глюкозы; Б — снижение активности через 20 секунд после введения глюкозы; В — прекращение активности через 30 сек. после введения глюкозы.
Pис. 2. Нейрон с исходной фоновой активностью из передних отделов коры мозга голодной кошки (уретановый наркоз).
А — активность до введения глюкозы; Б — увеличение активности через 47 сек. после введения в кровь глюкозы.
Таким образом, можно видеть, что у голодного животного в ответ на введение в кровь глюкозы наиболее типичной реакцией нейронов является снижение их исходной фоновой активности, нередко до полного се прекращения.
С целью контроля специфичности этих реакций в некоторых случаях мы вводили в кровь голодному животному физиологический или гипертонический растворы хлористого натрия. Как правило, при введении физиологического раствора нейроны не изменяли частоту своих разрядов или несколько ее увеличивали в случаях введения гипертонического раствора.
На основании полученных данных мы пришли к выводу, что у кашек после двухсуточного голодания повышенная исходная фоновая активность кормовых нейронов, реагирующих на введение в кровь глюкозы, отражает «голодную» активацию передних отделов коры мозга, которая в ЭЭГ выражается высокочастотными, низкоамплитудными колебаниями биопотенциалов.
Этот вывод подтвердили результаты несколько экспериментов (3 опыта), проведенных на предварительно накормленных кошках, ЭЭГ которых под уретановым наркозом характеризовалась наличием высокоамплитудных медленных колебаний биопотенциалов во всех отделах коры головного мозга. При исследовании у этих животных реакций нейронов передних отделов коры головного мозга на введение в кровь глюкозы оказалось, что при этом большинство нейронов не меняет частоты своей исходной импульсации (75%) (табл. 4). Только несколько нейронов у этих животных снижало частоту разрядов после введения глюкозы. Несколько «молчащих» нейронов при этом, наоборот, активировалось (табл. 1). Эти контрольные эксперименты показали, что функциональное состояние нейронов передних отделов коры больших полушарий голодного животного существенно отличается от функционального состояния нейронов этих же отделов кары у накормленных животных.
Следует также отметить тот факт, что реакция корковых нейронов у голодных животных на введение в кровь глюкозы зависела и от способа ее введения (внутриартериальный или внутривенный) (табл. 2).
Таблица 2
Распределение нейронов по типам в зависимости от ответной реакции на введение глюкозы различными способами

Из таблицы видно, что при внутриартериальном способе ведения глюкозы нейроны более четко изменяют свою активность, чем при внутривенном. При последнем способе введения большое количество нейронов (51,2%) не дает ответных реакций. В наших опытах физиологический эффект изменения клеточной активности обычно наступал через 40—50 сек после введения глюкозы. В ряде случаев он наступал несколько быстрее (через 20—30 сек), а в некоторых — активность нейронов изменялась через 5—10 секунд после введения глюкозы. Такие латентные периоды одинаковы для всех типов реакций нейронов, но следует отметить, что при внутривенном введении глюкозы по сравнению с внутри артериальным латентные периоды реакций были более короткими.
При анализе полученных нами экспериментальных данных перед нами прежде всего возникли следующие вопросы: чем обусловлена высокая активность корковых нейронов у голодного животного и каков механизм ответных реакций корковых нейронов на введение в кровь глюкозы?
В свое время еще И. П. Павлов (4) указывал на то, что одним из главных факторов в возникновении голодного состояния животного является снижение уровня питательных веществ в крови («голодная кровь»). Работами Майера (14, 15, 16) было довольно точно доказано существование в гипоталамусе рецепторов, специфически чувствительных к уровню сахара крови, на основании чего им была выдвинута «глюкостатическая» теория регуляции пищевого поведения. Проводимые многими авторами исследования по изучению нейрофизиологических механизмов пищевых реакций убедительно показали наличие в гипоталамических отделах мозга центров «голода» и «насыщения» (8—10, 13, 15). Наличие двусторонних морфологических связей гипоталамических пищевых центров с передними отделами коры больших полушарий указывает на их тесную функциональную связь (11, 19). Эта функциональная связь у голодного животного проявляется в активирующем влиянии со стороны гипоталамуса на передние отделы коры головного мозга (1, 2, 5, 6, 7). Исходя из этого, можно думать, что установленная в наших экспериментах высокая активность нейронов передних отделов коры больших полушарий отражала пищевое возбуждение, формируемое восходящими влияниями пищевых центров гипоталамуса.
Это означает, что у голодных животных синаптические организации коркового нейрона вовлечены в доминирующее пищевое возбуждение. Показателем того, что данный нейрон находится под влиянием именно «голодного» восходящего возбуждения со стороны гипоталамуса для нас является реакция этого нейрона на введение в кровь глюкозы. По мере воздействия ее на специфические глкжорецепторы снижалась возбудимость гипоталамичеокого центра «голода», а это приводило к уменьшению восходящих влияний на кору головного мозга со стороны этого центра, результатом чего являлось прекращение нейронной активности коры. Однако наряду с центральными рецепторами имеется большое поле периферических рецепторов, сигнализирующих об уровне глюкозы а крови.
Существует точка зрения, согласно которой в печени имеются рецепторы, информирующие центральную нервную? систему о внутриклеточной концентрации глюкозы и близких к ней метаболитов (17). В последнее- время стали известны факты, говорящие о том, что уровень пищевой возбудимости зависит от артериавенозной разницы в концентрации глюкозы в крови (9, 18). Аналогичный вывод можно сделать и на основании наших экспериментов. Как указывалось выше, нами было установлено, что латентные периоды реакций нейронов на введение в кровь глюкозы были значительно меньше при внутривенном способе введения, чем при артериальном. Этот, казалось бы, парадоксальный факт, на наш взгляд, говорит о наличии периферических рецепторов, чувствительных к изменению уровня питательных веществ в крови. Кроме того, при внутривенном способе введения глюкозы быстро уменьшалась артериовенозная разница в концентрации сахара крови, что приводило к снижению пищевой возбудимости. Однако, все изменения уровня сахара крови, которые воспринимаются периферическими рецепторными зонами в конечном счете достигают в первую очередь гипоталамических пищевых центров, а затем уже по принципу восходящих влияний адресуются к коре головного мозга. Мы, конечно, не можем полностью отрицать возможность непосредственного воздействия глюкозы на нейроны передних отделов коры головного мозга, однако данных, подтверждающих это, в литературе нам найти не удалось.
Как показали наши эксперименты, наряду с нейронами,, прекращающими свою активность в ответ на введение глюкозы в кровь, в передних отделах коры мозга голодного животного встречались также нейроны, которые при этом увеличивали частоту своих разрядов. Вероятно, эти нейроны находились под влиянием гипоталамического центра «насыщения», так как по данным некоторых авторов, этот центр повышает свою возбудимость после введения глюкозы в кровь животному (8, 9, 20). Под влиянием того -же центра, на наш взгляд, находились и те «молчащие» нейроны, которые после введения глюкозы активировались.
Наличие в передних отделах коры головного мозга голодного животного большого количества нейронов, имеющих исходную фоновую активность и не реагирующих на введение глюкозы, в некоторых случаях, очевидно, было обусловлено способом ее введения (табл. 2). С другой стороны, возможно, что исходная фоновая активность этих нейронов отражает какие-то другие корковые активации, которые мы не можем выявить в наших экспериментах.
На основании вышеизложенных экспериментальных данных можно сделать вывод, что при введении в кровь голодному животному глюкозы, в передних отделах коры головного мозга избирательно вовлекаются в реакцию клеточные системы, которые как-то в прошлом были связаны с пищевым поведением животного. Эти нейронные системы мобилизуются теми микропроцессами, которые разыгрываются в гипоталамусе на границе: чувствительная клетка — кровь, и которые являются стимулом для развертывания организмом вполне определенной, биологически целенаправленной пищевой деятельности.
ЛИТЕРАТУРА
1. Анохин П. К. Журн. в. н. д., 1959, 9, 4, 489.
2. Анохин П. К. Журн. в. н. д., 1962, 12, 3, 379.
3. Анохин П. К. Физиологическая архитектура поведенческих актов разной сложности. Конф. «Физиологические основы сложных форм поведения», реф. докл., M-J1., 1963.
4. Павлов И. П. О пищевом центре. «Тр. об-ва русских врачей», 1911, т. 78, 31.
5. Судаков К. В. Бюлл. эксперим. биол. и мед., 1962, 8, 3.
6. Судаков К- В. Физиол. журн. СССР, 1963, 49, 8, 901.
7. Судаков К- В., Фадеев Ю. А. Физиол. журн. СССР, 1963, 49, 11, 1310.
8. A nan d В. К. Physiol. Revs., 1961, 41, 4, 677.
9. An and В. К. Amer. J. Physiol., 1964, 207, 5, 1146.
10. Brobeck J. R. Gastroenterology, 1957, 32, 2, 169.
11. С 1 a r k Le G г о s. Lancet, 1948, 254r 6497, 353.
12. Hess W. R. Hypothalamus und Thalamus. Stuttgart, 1956.
13. Larsson S. Acta physiol. Scandinav. (supp. 115), 1954, 32, 1.
14. Mayer J. Ann. N. J. Acad. Sci., 1955, 63, 15.
15. Mayer J. Clin. Research. Proc., 1957, 5, 123.
16. Mayer J. New England. J. Med., 1953, 249, 13.
17. Russek M. Nature (Engl), 1963, 197, 4862.
18. V a n S t a 1 i e Т. B. Diabetes, 1959, 8, 226.
19. Ward A. A., Me Culloch W. S.J. Neurophysiol., 1947, 10,
20. Yulaka Oocuura. Science, 1964, 143, 3605, 484, etal.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
§ 28. Возникновение отделов головного мозга
§ 28. Возникновение отделов головного мозга Ранний период истории возникновения предков позвоночных, до формирования хорошо структурированного скелета, довольно туманен. Если допустить, что предковые формы хордовых были мягкотелыми существами размером около 10–15 см,
Увеличение головного мозга
Увеличение головного мозга Итак, чтобы объяснить, откуда возникло название этой главы, следует прежде всего подчеркнуть, что мы понимаем под поведением, и затем показать особую связь поведения с мозгом. Термин «поведение» имеет самые разнообразные значения; мы же будем
8.2. Паттерны системной специализации нейронов разных структур мозга
8.2. Паттерны системной специализации нейронов разных структур мозга Для того чтобы получить данные, непосредственно характеризующие проекцию тех или иных элементов опыта на структуры мозга, необходимо сопоставить паттерны системной специализации нейронов этих
ЛИМБИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА
ЛИМБИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА Лимбическая система в мозге человека выполняет очень важную функцию, которая называется мотивационно-эмоциональной. Чтобы было ясно, что это за функция, вспомним: каждый организм, включая организм человека, имеет целый набор
Травмы головного и спинного мозга
Травмы головного и спинного мозга Несомненно, что всех животных с травматическим повреждением любого вида, в том числе и с травмой головного и спинного мозга, прежде всего должен осмотреть ветеринар — это необходимо для уточнения диагноза и определения показаний для
Болезни головного мозга
Болезни головного мозга При заболеваниях головного мозга развиваются следующие расстройства: судороги, параличи, которые возникают в результате поражения двигательной области коры головного мозга или двигательных путей от коры в боковой столб противоположной
4.1.1. Органическое поражение головного мозга
4.1.1. Органическое поражение головного мозга Животное пытается достать лакомство, но быстро прекращает попытки (истощаемость), либо не пытается достать лакомство (депрессия, апатия), либо постоянно отвлекается на другие внешние раздражители, вплоть до переключения на
Травмы головного мозга
Травмы головного мозга Встречаются относительно редко.¦ ЭТИОЛОГИЯ И ПАТОГЕНЕЗТравмы головного мозга регистрируются после ударов или падения с высоты. Сопровождаются сотрясением мозга и кровоизлияниями различной степени.¦ СИМПТОМЫПосле удара или падения собака
5. Анализаторная и синтетическая деятельность коры головного мозга
5. Анализаторная и синтетическая деятельность коры головного мозга Высшая нервная деятельность животного дает возможность выделять ему из сложной массы всевозможных раздражений, идущих из внешнего мира, те, которые имеют для животного то или иное значение. Собака при
Восходящие активирующие влияния на кору головного мозга у голодных животных К. В. СУДАКОВ (Москва)
Восходящие активирующие влияния на кору головного мозга у голодных животных К. В. СУДАКОВ (Москва) В последние годы благодаря применению новых методов физиологического эксперимента (электронные усилители, стереотаксическая техника) проблема голода, пищевого поведения
Анализ физиологических механизмов восходящих активирующих влияний на кору мозга у голодных животных методом аппликации различных фармакологических веществ А. И. ТУРЕНКО (Москва)
Анализ физиологических механизмов восходящих активирующих влияний на кору мозга у голодных животных методом аппликации различных фармакологических веществ А. И. ТУРЕНКО (Москва) Многочисленные исследования показали, что основным регулятором потребления пищи является
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва)
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва) Изучению реакции организма на длительное голодание посвящено много работ. Большинство из них касается обменных процессов, биохимии синтеза и распада
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва)
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва) В связи с тем вниманием, которое в настоящее время уделяется роли лизосом в развитии повреждения клеток тканей различных
Влияние голодания на активность ферментов пентозофосфатного пути в печени и мозге крыс Ю. Л. ЗАХАРЬИН (Москва)
Влияние голодания на активность ферментов пентозофосфатного пути в печени и мозге крыс Ю. Л. ЗАХАРЬИН (Москва) В последние годы в клинике часто применяется с лечебными целями, в частности, для лечения психических заболеваний, полное голодание. Не вызывает сомнения, что