Сравнительное изучение действия полного длительного голодания И белковой недостаточности на состав периферической крови мышей СС57 Вr И. Л. ПОВЕРИЙ и В. И. ПРИЛЯЦКИЙ (Москва)
Сравнительное изучение действия полного длительного голодания И белковой недостаточности на состав периферической крови мышей СС57Вr
И. Л. ПОВЕРИЙ и В. И. ПРИЛЯЦКИЙ (Москва)
Полное или частичное голодание является удобной экспериментальной моделью для изучения влияния обменных процессов на разнообразные формы жизнедеятельности организма, в том числе на клеточный состав кроветворных органов.
Как показали исследования XIX века (9, 12, 15, 18) и многочисленные материалы сравнительно последних лет (2, 8, 11, 13, 23 и др.) в разные стадии полного голодания в периферической крови различных млекопитающих наблюдаются определенные изменения концентрации и состава лейкоцитов. При этом концентрация лимфоцитов в отдаленные периоды голодания, как правило, уменьшается, а изменения лейкоцитарной формулы зависят, по-видимому, как от видовых особенностей животного, так и от фазы процесса.
Г. Селье (19, 25) полагал, что полное голодание является «стрессовым» фактором, воздействие которого на животных сопровождается нейтрофилезом, лимфопенией и эозинопенией.
Лимфатическая ткань у млекопитающих тонко реагирует на голодание. Так, при полном лишении животного пищи в тимусе, селезенке и лимфатических узлах возникают (особенно отчетливо заметные в терминальные периоды голодания) атрофические и дегенеративные явления (6, 16, 17, 25 и др.).
В то же время имеются данные об относительной устойчивости к голоданию циркулирующих лимфоцитов (1, 4, 21). Так, было отмечено повышение относительной концентрации лимфоцитов в крови голодающих крыс и людей (1, 23). У отдельных животных и людей доля лимфоцитов в формуле крови достигла 98—98,5%. Однако, у голодающих молодых кроликов подобных сдвигов не отмечалось (7).
Целью настоящей работы являлось изучение морфологических изменений в периферической крови мышей СС57Вr при содержании животных на различных диетах и при полном голодании.
Выяснялись следующие вопросы:
а) зависимость морфологических показателей периферической крови мышей от содержания белка в диете и сравнение этих особенностей у животных, находящихся на безбелковой диете, с изменениями, происходящими при полном голодании;
б) в какой степени проявляются «стрессовые» изменения в периферической крови мышей при разных степенях белковой недостаточности и при полном голодании.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперименты проведены на 91 мышах-самцах линии СС57Вr, возраст животных 2—3 и 4—6 месяцев. Мыши данной линии устойчивы к различным воздействиям (14).
Опыты состояли из нескольких серий: в серии 1—животные лишались всякой пищи, кроме воды (полное голодание). Часть опытов этой серии была проделана в июле (6 мышей — подсерия 1а), часть — в ноябре-декабре (40 мышей — подсерия 16); в серии 2 мыши содержались на стандартизированной безбелковой диете, включавшей жиры и углеводы в количестве достаточном для стабилизации энергетического баланса в организме мышей. Эта серия также имела подсерии — 2а (июль, 5 животных) и 26 (ноябрь, декабрь — 5 животных); в серии 3 (июль) 5 мышей содержались в течение 4 дней на стандартизированной диете с 5% белка; в серии 4, часть которой проводилась в течение марта—апреля—мая (подсерия 4а — 30 мышей), а часть в течение ноября (подсерия 46 — 5 животных) мыши, в течение месяца, находились на стандартизированной диете, с 5% белка; в серии 5 (20 животных в марте—апреле—мае—подсерия 5а, и 5 мышей в ноябре — подсерия 56) животные получали в течение месяца стандартизированную диету с 20% белка[11]. Мыши более старшего возраста были использованы в опытах летних серий в связи с тем, что эти животные в силу большей упитанности и оптимальных температурных условий лета, позволяли выяснить предельную устойчивость мышей к голоданию.
Во всех сериях опытов у мышей производились регулярные исследования концентраций гемоглобина, эритроцитов и лейкоцитов в периферической крови, а также определялись лейкоцитарные формулы, по данным которых вычислялись абсолютные концентрации нейтрофилов (палочко-ядерные + сегментоядерные), лимфоцитов и эозинофилов.
Так как в сериях 1, 2 и 3 наблюдения продолжались не более 6 дней, то за контроль были приняты значения соответствующих показателей до качала опыта, а также данные специального исследования сезонной периодичности гематологических показателей у здоровых мышей СС57Вr (данные от 83 мышей для ноября—декабря и данные от 20 мышей для июля). Эти же показатели (для ноября—декабря) приняты за контрольные для подсерий 46 и 56.
Контролем для подсерий 4а и 5а служила специальная группа из 55 мышей, получавших обычную полноценную диету, содержавшую зерно, овощи, молочные продукты и т. п, наблюдения над которыми проводились синхронно, с опытом.
Количественные данные подвергались биометрической обработке. При большом количестве наблюдений (п>10) вычислялась стандартная ошибка средней арифметической, при анализе малых выборок использовались непараметрические критерии. Производился коррелятивный анализ.
РЕЗУЛЬТАТЫ
Среднестатистические показатели периферической крови у мышей СС57Вr в июле были следующими: концентрация гемоглобина 15,4±0,09 г% (вариации 14,8—16,4 г%), концентрация эритроцитов — 11,1±0,21 млн. на мм3 (вариации 10,0 • 106 — 12,7 • 106 на мм3), концентрация лейкоцитов — 15,4 + 0,10 тыс. на мм3 (14,5 • 103 — 16,1 • 103 на мм3), концентрация нейтрофилов (суммарно палочкоядерные и сегментоядерные) 6,3±0,5 тыс. на мм3 (1,9 • 103 — 9,0 • 103), концентрация лимфоцитов — 8,3±0,6 тыс. на мм3 (4,9 • 103 — 13,6 • 103), концентрация эозинофилов — 0,55 тыс. на мм3 (0,15-103— 0,95-103).
Аналогичные показатели в ноябре-декабре были следующими: гемоглобин — 14,7±0,05 г% (13,8—15,0 г%), эритроциты 12,1 ±0,14 млн. на мм3 (10 • 10 — 13 • 10 на мм3), лейкоциты 15,2±0,18 тыс. на мм3 (12,8 • 103 — 16,9 • 103 на мм3), нейтрофилы 6,0±0,48 тыс. на мм3 (4,1 • 103 — 8,7 • 103 на мм3), лимфоциты 7,5±0,43 (5,0 • 103 — 10,2 • 103 на мм3, эозинофилы — 0,3 тыс. на мм3 (0,14 • 103 — 0,55 • 103 на мм3).
В первой серии опытов голодающие животные в группе 1а (июль) жили не более 5 суток, а в группе 16 (ноябрь-декабрь) не более 2 суток. Потеря веса у мышей при полном голодании достигала 20—30% от исходного.
В сериях 2 и 3 опыт был прекращен на 4—6 день и животные были забиты для гистологического исследования. Потеря веса в этих сериях колебалась от 7% до 21% от исходного.
Изменения гематологических показателей у мышей в 1, 2 и 3 сериях опытов представлены в таблицах 1 и 2.
Как следует из таблиц 1 и 2, в отдаленные сроки полного голодания у мышей наблюдается тенденция к уменьшению концентраций гемоглобина и эритроцитов в периферической крови, причем у животных, погибших от истощения на 6 день (июль, упитанные мыши 4—6 месяцев), анемичность к концу опыта была выражена в большей степени, чем у животных, погибших на 2 день голодания (ноябрь-декабрь, мыши 2—3 месяцев).
Таблица 1
Изменения концентраций гемоглобина, эритроцитов и лейкоцитов в периферической крови у мышей СС57Вr при полном голодании, безбелковой диете и при диете 5% белка (показатели в конце опыта)
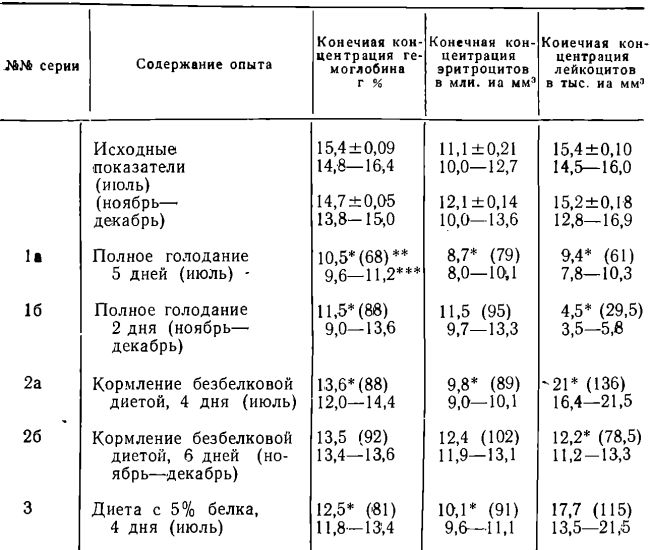
* Достоверное отличие от исходного показателя;
** В % к исходному показателю;
*** Индивидуальные вариации.
Общая концентрация лейкоцитов в те же сроки голодания значительно снижается, причем в осенней подгруппе животных этот показатель уменьшается быстрее, чем у голодающих мышей из летней подгруппы. Концентрация нейтрофилов у мышей, голодавших в июле, уменьшается к концу опыта в той же степени, что и у животных, голодавших в ноябре-декабре, но лимфопения при голодании выражена значительно больше у- животных из осенней подгруппы. Изменения концентрации эозинофилов у голодающих животных подсерии 1а (июль) малодостоверны, однако у животных подсерии 16 (ноябрь-декабрь) к концу опыта отмечается значительная эозинопения (в среднем 0,05 • 103 эозинофилов на мм3).
Таблица 2
Изменения концентраций нейтрофилов и лимфоцитов в периферической крови у мышей СС57Вr при полном голодании, при безбелковой диете и при диете с 5% белка (показатели по дням опыта)

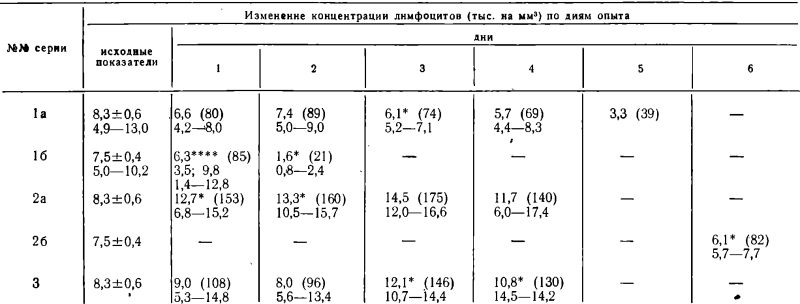
Примечания: Форма таблицы и обозначения аналогичны табл. 1.
**** «3,5; 9,8» — выборочные средние арифметические для групп животных, на которых опыт был поставлен в ноябре или декабре соответственно.
6.3 — общая средняя арифметическая.
Корреляционный анализ показал, что в течение голодания в серии 1 (а, б) отмечается положительная корреляция концентраций эозинофилов и лимфоцитов (р>0) и отрицательная корреляция концентраций эозинофилов r<0).
При кормлении мышей безбелковой диетой в течение 4—6 дней (серия 2 (а, б)) отличия в показателях гемоглобина и эритроцитов были незначительны. У мышей, содержавшихся на диете без белка в июле, наблюдалось увеличение общей концентрации лейкоцитов, которое связано с лимфоцитозом, в то время как концентрация нейтрофилов у них или не изменяется или уменьшается. У мышей, содержавшихся на безбелковой диете в ноябре-декабре, отмечается незначительная лейкопения за счет некоторой лимфопении. Концентрация эозинофилов у животных серии 2 (а, б) существенно не меняется. Коррелятивный анализ не дал определенных результатов.
При содержании мышей в течение 4 дней на диете с 5% белка (серия 3, июль) у них возникает незначительная анемия, сочетающаяся с тенденцией к лимфоцитозу; концентрация эозинофилов достоверно не менялась. Коррелятивный анализ позволил обнаружить в серии 3 отрицательную зависимость между относительными величинами лимфоцитов и нейтрофилов, и эозинофилов и нейтрофилов, и положительную зависимость концентраций эозинофилов и лимфоцитов.
Изменения гематологических показателей у мышей в 4 и 5 сериях опытов представлены в таблице 3.
Месячное содержание мышей на стандартизированных диетах с 5% и 20% белка в течение весеннего периода не привело к существенным изменениям состава периферической крови животных. В серии 4а обнаружена достоверная эозинофилия (6% эозинофилов в формуле и 0,7 тыс. на мм3).
Кормление мышей стандартизированными диетами в осенний период вызывает у них тенденцию к лимфоцитозу, которая лучше выражена при содержании животных на диете (5% белка). В серии 56 наблюдается также нейтропения. Кроме того, в сериях 46 и 56 была достоверная эозинофилия (0,8 тыс. на мм3 и 0,65 тыс. на мм3 соответственно).
Таблица 3
Изменение гематологических показателей периферической крови мышей СС57Вr при месячном кормлении их стандартизованными диетами с различным содержанием белка
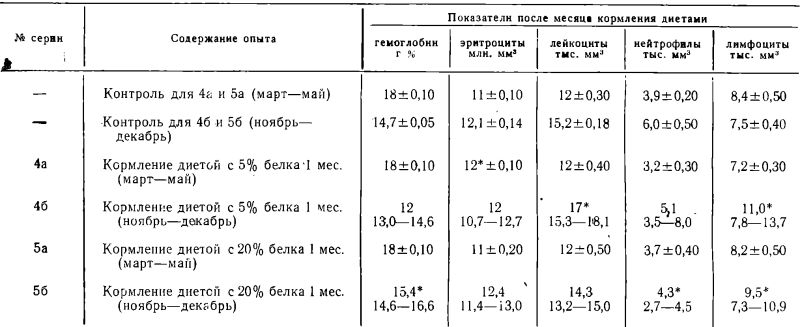
* Отличия от контроля достоверны.
Обсуждение результатов
Как следует из полученных данных, изменение состава периферической крови в отдаленные сроки полного голодания (с водой) у мышей СС57Вr заключается в тенденции к анемии и в существенной лейкопении. При этом концентрация лимфоцитов уменьшается в большей степени, чем концентрация нейтрофилов. Изменения концентрации эозинофилов при полном голодании находятся в положительной корреляции с изменениями концентрации лимфоцитов.
Таким образом, для состава лейкоцитов в периферической крови у голодающих мышей характерно: снижение абсолютных количеств нейтрофилов, лимфоцитов и эозинофилов (в 1 мм3 крови) и относительный нейтрофилез. Эти данные соответствуют в качественном и количественном отношении динамике показателей периферической крови при других острых «стрессовых» состояниях у мышеи. В то же время сведения об относительной устойчивости к голоданию концентрации лимфоцитов в периферической крови (полеченные в эксперименте на других животных: собаки, кролики, крысы, а также при голодании людей) не подтверждаются нашими данными, полученными при голодании мышей.
Устойчивость мышей СС57Вr к голоданию зависит от состояния животных. В летнее время животные старшего возраста (4—6 месяцев) более устойчивы к голоданию. Они могут жить при полном отсутствии пищи (но с водой) до 6 суток. В конце осени мыши линии СС57Вr (2—3 месяца) погибают после 2 дней полного голодания, что соответствует распространенным для этих животных литературным данным, где, очевидно, авторы изучали продолжительность жизни голодающих белых мышей в осенне-зимний период (10, 24).
Не исключено, что различное отношение к голоданию мышей в связи с сезоном находится в известной прямой зависимости изменения показателей крови и других параметров физиологических процессов грызунов от времени года (3, 20 и др.).
У животных из летней и осенней подгрупп в терминальный период полного голодания лимфопения и эозинопения в серии 16 достигают максимальной степени. Таким образом, эти показатели отражают степень тяжести состояния подопытных животных.
При содержании мышей в течение 4—6 дней на безбелковой диете или на рационе, содержащем 5% белка, показатели концентраций гемоглобина и эритроцитов существенно не меняются, концентрации нейтрофилов и эозинофилов в целом остаются стабильными, концентрация лимфоцитов или возрастает (до 175%) или падает (до 82%). Различия в изменениях числа лимфоцитов также зависят от сезонного фактора.
При длительном содержании мышей на диете с 5% белка (до 1 мес.) концентрация лимфоцитов повышается к концу опыта до 147% у осенней подгруппы животных, но остается стабильной у животных весенней подгруппы. Напротив, у мышей, длительно получавших дефицитную по белку пищу в весенний период, отмечается эозинофилия. Прочие гематологические показатели в таких случаях были относительно стабильны.
У мышей, получавших в течение месяца диету с 20% белка в весенний период, исследованные показатели существенно не изменились, однако, у животных из осенней подгруппы месячное содержание на данной диете вызывает нейтропению и лимфоцитоз, при стабилизационной суммарной концентрации лейкоцитов. Таким образом, лимфоцитоз у мышей является не только реакцией на белковую недостаточность, но и на изменения качественного состава пищи.
Из серии 2, 3, 4 следует, что мыши СС57Вr устойчивы к белковой недостаточности.
Основные морфологические показатели периферической крови при диете с низким содержанием белка, как правило, поддерживаются на уровне не ниже «нормальных» цифр данного показателя.
Выводы
1. Устойчивость мышей СС57Вr к полному голоданию зависит от состояния животных, в частности, от возраста и сезонного фактора.
2. Изменение состава лейкоцитов в периферической крови мышей СС57Вr при полном голодании напоминает сдвиги, полученные при острых «стрессовых» реакциях: снижение абсолютных концентраций нейтрофилов, лимфоцитов и эозинофилов и относительный нейтрофилез.
3. При значительном снижении количества белка в диете, организм мышей СС57Вr способен стабилизировать основные морфологические показатели периферической крови.
4. При содержании мышей СС57Вr на стандартизованных диетах с 5% и 20% белка (полное покрытие суточного количества требуемого белка) в периферической крови животных часто отмечается лимфоцитоз. В одной из серий опытов с кормлением животных белководефицитной диетой наблюдалась статистически значимая эозинофилия.
5. Лимфоцитоз и эозинофилия находятся в некоторой зависимости от сезонного фактора и отражают или реакцию организма на белковую недостаточность или изменившийся качественный состав пищи.
6. Отсутствие белка в диете не препятствует возникновению лимфоцитарной реакции.
ЛИТЕРАТУРА
1. Агамова К. А. Костномозговое кроветворение при голодании и влияние на него внутривенных вливаний видовонеспецифической сыворотки. Экспериментальное исследование. Автореферат дисс. канд., М., 1955.
2. А г а м о в а К. А. Ученые записки 2 Моск. мед. ин-та, 1958, 16, 7.
3. Б а ш е н и и а Н. В. Экология обыкновенной полевки. М., 1962.
4. Бекенская А. И. Сб. научн. труд. Гос. ин-та усоверш. врачей им. Кирова, за 2-ой год Отеч. войны. Л,, 1944, 212.
5. В о л г а р е в Н. М. Бюлл. эксп. биол. и мед., 1957, 99, 12. 108.
6. Воронцова М. А. и Лиознер Л. Д. Физиологическая регенерация. М., 1955.
7. Зайцева Р. А. Регенерационные процессы в костном мозгу у молодого организма в норме, при голодании и парентеральном питании. Автореферат дисс. канд. М., 1956.
8. Зайцева Р. А. Ученые записки 2 Моск. мед. ин-та, 1958, 16, 30.
9. Каган Я. А. Кровь и кровяное давление у голодающих. СПБ. 1884.
10. К о н с т а н т и н о в В. А. В кн.: Механизмы патологических реакций. Л., 1950, 16, 180.
11. Кочарова Е. А. В кн.: Труды V Всесоюзного съезда анатомов, гистологов, и эмбриологов. Л., 1951, 502.
12. Любомудров П. В. Изменение крови и некоторых органов при голодании. Дисс. СПБ, 1893.
13. М а ч а в а р и а и и Э. Н. Морфологич. изменения периферической крови и кроветворного аппарата при экспериментальной алиментарной дистрофии. Автореферат дисс. канд., Тбилиси, 1952.
14. Медведев Н. Н. Вопросы онкологии, 1962, 8, 7, 120.
15. Окончиц Е. С. К вопросу об изменении морфолог, состава крови у кроликов при полном голодании и послед, откармливании. Варшава, 1893. Отуиск из «Архива лаборатории общей патологии» при Варшавском университете.
16. Поберий И. А. Ученые записки 2 Мск. мед. ин-та, 1958, 16, 63.
17. Полетаев П. И. Морфологический состав крови при полном и неполном (с водой) голодании у собак. Дисс. Сиб., 1894.
18. Селье Г. Бюлл. эксп. биол. и мед., 1939, 8, 9, 11, 339.
19. С л о н и м А. Д. Частная экологическая физиология млекопитающих. М.,—Л., 1962.
20. Чельцова О. Н. Журн. теор. и практ. мед., 1925, 1, 3—4, 363.
21. Ч е р к е с Л. А., С т р у к о в А. Й*., В о л г а р е в М. Н., Смирно в В. П. Вопросы питания, 1960, 19, 1, 3.
22. Шапиро Ю. Л. Патол. физиология и эксперимент, терап. 1963, 7 1 39.
24. D i 11 е г J. Anat. Rec., 1946, 96, 4, 562.
25. S е 1 у е Н. Physiolegy and pathology of the expesure to stress. Montreal, 1948.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Развитие идей лечебного голодания Ю. С. НИКОЛАЕВ (Москва)
Развитие идей лечебного голодания Ю. С. НИКОЛАЕВ (Москва) Можно считать, что люди еще с доисторических времен применяли голодание с лечебной целью; это подтверждается наблюдениями за животными, которые при заболевании отказываются от приема пищи, этому же инстинктивно
О механизме терапевтического действия лечебного голодания
О механизме терапевтического действия лечебного голодания Несмотря на значительную давность применения лечебного голодания, до настоящего времени остается много неясного в вопросах терапевтического действия этого метода лечения. Этим в большой степени объясняется
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва)
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва) Лечение ожирения остается одной из наиболее актуальных проблем современной медицины. Число больных с избыточным весом, согласно данным многих
Случай применения лечебного голодания у тучного больного, перенесшего инфаркт миокарда М.И. МИНЯЙЛЕНКО (Москва)
Случай применения лечебного голодания у тучного больного, перенесшего инфаркт миокарда М.И. МИНЯЙЛЕНКО (Москва) В литературе нам встретились ряд работ, указывающих на эффективность кратковременного воздержания от приема пищи при целом ряде соматических заболеваний.Так
О влиянии дозированного голодания на состояние слизистой желудка М. И. МИНЯЙЛЕНКО, А. И. БУХМАН, И. И. ЛУЧИНСКИЙ (Москва)
О влиянии дозированного голодания на состояние слизистой желудка М. И. МИНЯЙЛЕНКО, А. И. БУХМАН, И. И. ЛУЧИНСКИЙ (Москва) Голодание нередко приводит к изменениям со стороны желудочно-кишечного тракта. По данным Ю. С. Николаева, вторая стадия голодания сопровождается
Динамика показателей периферической крови при лечебном голодании у больных гипертонической болезнью и ожирением Г. Н. БЖИШКЯН-БОРОДИНА (Москва)
Динамика показателей периферической крови при лечебном голодании у больных гипертонической болезнью и ожирением Г. Н. БЖИШКЯН-БОРОДИНА (Москва) Литературные данные относительно исследования состава периферической крови при лечебном голодании представляют большую
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва)
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва) Изучению фагоцитарной активности лейкоцитов при длительном полном алиментарном голодании посвящены
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва)
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва) Известно, что так называемые «жесткие константы» сохраняются неизменными в организме В течение длительного срока, несмотря на воздействие каких-либо «отклоняющих» факторов.
О действии полного длительного алиментарного голодания на хромосомный аппарат лимфоцитов периферической крови К. Н. ГРИНБЕРГ, Ю, Л. ШАПИРО, Е. А. КИРИЛОВА, Р. С. КУШНИР (Москва)
О действии полного длительного алиментарного голодания на хромосомный аппарат лимфоцитов периферической крови К. Н. ГРИНБЕРГ, Ю, Л. ШАПИРО, Е. А. КИРИЛОВА, Р. С. КУШНИР (Москва) Полное алиментарное голодание успешно применяется при лечении некоторых психических и
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва)
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва) Половой хроматин — внутриядерное тельце, дающее Фельген-положительную реакцию и
Условнорефлекторные лейкоцитарные реакции в динамике лечебного голодания при некоторых нервно-психических заболеваниях С. М. ЛУРЬЕ (Москва)
Условнорефлекторные лейкоцитарные реакции в динамике лечебного голодания при некоторых нервно-психических заболеваниях С. М. ЛУРЬЕ (Москва) Учение И. П. Павлова «Об организме, как о целостной системе, как о функциональном единстве всех органов и тканей, психического и
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва)
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва) В связи с тем вниманием, которое в настоящее время уделяется роли лизосом в развитии повреждения клеток тканей различных
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва)
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва) Голодание как метод лечения успешно зарекомендовал себя при некоторых психических и соматических заболеваниях (3, 7, 10—13). Особенно перспективно его применение при обменных, аллергических
Влияние голодания на активность ферментов пентозофосфатного пути в печени и мозге крыс Ю. Л. ЗАХАРЬИН (Москва)
Влияние голодания на активность ферментов пентозофосфатного пути в печени и мозге крыс Ю. Л. ЗАХАРЬИН (Москва) В последние годы в клинике часто применяется с лечебными целями, в частности, для лечения психических заболеваний, полное голодание. Не вызывает сомнения, что