Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва)
Эритрон при длительном алиментарном голодании и последующем питании людей
Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва)
Голодание, как частный раздел науки о питании, за последние годы вновь стало привлекать к себе пристальное внимание в связи со все более широким его использованием для изучения многих биохимических, патофизиологических, биологических и других проблем (обмен веществ и энергии, регенерация, компенсация функций, выживаемость, регуляция кроветворения, в частности, исследование гемоноэтинов и проч.). С полным алиментарным голоданием нередко приходится встречаться в медицинской практике, при стихийных и социальных бедствиях. Особый интерес представляет опыт применения полного алиментарного голодания с лечебной целью как в качестве самостоятельного метода, так и в комбинации с другими терапевтическими средствами воздействия (1, 11) и др.
Изучение системы крови, в том числе и красной, было начато еще на заре научной разработки голодания (12, 13). Большинство наиболее обстоятельных работ по изучению системы крови при полном голодании были выполнены в эксперименте (14—18). Сообщения о подобных исследованиях у полностью голодавших людей до сих носят казуистический характер (19—28).
Согласно данным большинства авторов при полном алиментарном голодании самых разнообразных животных и людей концентрация эритроцитов и содержание гемоглобина остаются весьма стабильными в течение довольно длительных его сроков (29—33, 37—39) по крайней мере до периодов, характеризующихся потерями веса тела на 10—15 % к исходному (34), а по данным некоторых авторов и до 26—30% (16, 35). Нередко в различные сроки голодания наблюдали и гем о концентрацию (26, 27, 29, 33 36—41). В более отдаленные сроки голодания, как правило отмечали умеренную гипонормохромную анемию (15, 16, 34, 35 и др.). После прекращения голодания относительное содержание эритроцитов и гемоглобина обычно кратковременно снижается, а затем довольно быстро достигает исходного уровня (19, 25, 42, 61) причем рост числа эритроцитов, как правило, опережает процесс гемоглобинизации (28, 42). Данные о распределении гемоглобина в эритроцитах неопределенны: чаще при голодании наблюдали нерезко выраженные колебания цветного показателя (15, 17, 23, 29). Наряду с указаниями на устойчивость объемного числа эритроцитов и гемоглобина было показано, что во время полного голодания общая масса крови снижается примерно параллельно падению «живой массы тела» (13, 43—45). Существуют предположения и доказательства о перераспределении циркулирующей и депонированной крови в пользу депо (21, 44). Одновременно следует отметить почти полное отсутствие каких-либо достоверных введений относительно изменения сроков жизни, резистентности (осмотической, кислотной, механической), тонкого строения и других морфофункциональных свойств эритроцитов при полном голодании. Данные авторов, находивших на основании исследований эритроцитов в жидкой среде изменения формы типа «шиповатости» и т.п., по-видимому, относятся к хорошо известному феномену «тутовой ягоды», характеризующему предлитическую сферуляцию эритроцитов (46—48). В более современных работах указания на существенные изменения формы эритроцитов при голодании отсутствуют. Наиболее часто отмечают тенденцию к микроцитозу, особенно отчетливо проявляющуюся в отдаленные сроки голодания, а иногда легкий анизоцитоз (12, 15, 17). На основании отдельных исследований можно полагать, что напряженность эритродиеретического процесса во время голодания остается сохранной[8], на что в частности, указывает непрекращающееся желчеобразование, а также гемосидероз селезенки, печени и костного мозга (14, 19, 49—57).
Не вызывает сомнения, что интенсивность эритропоэза при голодании (определяемая ретикулоцитарным, радиоизотопным и другими методами) во время полного алиментарного голодания отчетливо снижается, причем снижение идет в целом пропорционально длительности голодания (17, 19, 26, 59, 60). Среди причин, вызывающих депрессию эритропоэза во время голодания, особое значение придают снижению эритролоэтической активности крови. Широко известно, что экспериментальное голодание является одной из адекватных моделей при изучении гемолоэтинов (в первую очередь эритропоэтинов) (61—71). Интересно отметить, что снижение Эритропоэтической активности при голодании обратимо, а сам эритропоэз чрезвычайно чувствителен к эритропоэтическому фактору (72—80), что может свидетельствовать о функциональном характере этих изменений. Снижение эритропоэтической активности подтверждается также и немногочисленными исследованиями цитоморфологии костного мозга при голодании. Здесь наряду с устойчивым митотическим коэффициентом эритробластов находят уменьшение фронта эритробластичекого кроветворения (16, 17, 79, 81). В парциальных эритробластотраммах отмечают сдвиг вправо. Соотношение ретикулоциты костного мозга — ретикулоциты периферической крови смещается в пользу костномозговых ретикулоцитов (16, 17, 60). По-видимому, все эритроциты при голодании проходят стадию ретикулоцитов, а снижение активности эритропоэза зависит также от задержки элиминации эритроцитов (ретикулоцитов) из костного мозга на периферию (59, 60).
Спустя некоторый латентный период после прекращения голодания наблюдается отчетливое усиление репарации: увеличивается число митозов, расширяется фронт эритробластического кроветворения, отмечаются сдвиги влево в парциальных эритробластограммах и отчетливый ретикулоцитоз в периферической крови, предшествующий или совпадающий с подъемом числа эритроцитов (14, 16, 17/59, 60, 79).
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследование было проведено у 24 человек, полностью голодавших от 9 до 40 дней включительно. Из их числа мужчин было — 22, женщин — 2. Возраст колебался от 17 до 59 лет, причем до 40 лет включительно было 19 человек. Среди обследованных 3 человека были практически здоровы и голодали добровольно с целью эксперимента, а 21 — находились в психиатрическом стационаре, где они проходили курс лечения полным голоданием. Из числа последних у 17 была диагностирована шизофрения и у 4 — другие нервно-психические заболевания (хронический алкоголизм, ипохондрический синдром, травматическое поражение мозга, депрессивное состояние в климактерическом периоде). Двое из обследованных голодали повторно.
У обследованных лиц до голодания не было выявлено каких-либо грубых нарушений со стороны внутренних органов и ЦНС.
Методика голодания (по Ю. С. Николаеву) (5) в общих чертах сводилась к полному добровольному прекращению приема пищи без ограничения употребления воды, количество которой в процессе голодания индивидуально колебалось от 800 до 3500 мл в сутки. Во время голодания проводились различные общегигиенические физиотерапевтические мероприятия: прогулки на воздухе, массаж, субаквальные ванны и т, д. После прекращения голодания назначался специальный «щадящий» пищевой режим, заключавшийся в постепенном наращивании калорийности пищи с последовательным включением в пищевой рацион углеводов, белков и жиров.
Проведение полного лечебного голодания хорошо контролировалось персоналом, прошедшим специальную медицинскую подготовку; возможность самовольного приема пищи во время голодания полностью исключалась.
При исследовании показателей крови мы в основном пользовались гематологическими методами, достаточно известными в лабораторной практике и подробно описанными в специальных руководствах (82, 83).
Из показателей, характеризующих эритрон, были изучены следующие: количество эритроцитов в 1 мм3, содержание гемоглобина, показатели гематокрита крови, количество ретикулоцитов в 1 мм3, ретикулоцитограммы по Гейльмейеру, время созревания ретикулоцитов in vitro (84), средний-диаметр эритроцитов и эритроцитометрические кривые. На основании, этих данных рассчитывались средний объем и средняя толщина (высота), эритроцитов, условная форма эритроцитов, цветной показатель, средняя концентрация гемоглобина в одном эритроците (СКЭ), средняя площадь поверхности эритроцитов, интенсивность эритропоэза. Помимо этого определялась также концентрация билирубина в крови.
Кровь бралась утром (9 час. 20 мин.) до начала дополнительных лечебных процедур. Взятие крови из пальца, обработка пальца, транспортировка крови и ее обработка проводились с необходимыми предосторожностями, направленными на уменьшение лабораторных ошибок.
Кроме исследования периферической крови было также проведено изучение костномозговых пунктатов. Всего было получено 7 пунктатов (на 4, 9, 11 и 26 дни полного голодания и 7, 17 и 43 дни последующего питания) у трех обследованных лиц. Причем у одного из них, практически здорового добровольца-голодальщика, было получено в динамике 5 пунктатов. Костномозговые пункции проводили иглой Аринкина-Кассирского, в шприц набирали минимальное количество костномозгового содержимого (до появления первой капли над иглой в шприце). Окраска проводилась по Паппепгейму. В костном мозгу помимо общего описания препарата, сосчитывали парциальные эритробластограммы (на 200 элементов эритробластического ряда), определяли количество ретикулоцитов и составляли ретикулоцнтограммы.
Данные относительно всех параметров красной крови группировались по периодам голодания и питания, выделяемых при этом состоянии у людей, после чего обрабатывались статистически.
При голодании людей различают три периода самого голодания и три периода последующего питания. Первый период охватывает 1—4 дня голодания (потери веса тела равны примерно 5%); второй период — с 5 по 12 дни голодания (потери веса тела до 10%) и третий период голодания — остальной срок после 13 дня. В наших наблюдениях ни в одном из случаев потери веса тела к концу голодания не превышали 25% от первоначального. Таким образом весь 30—40-дневный срок голодания оказывается равным примерно 1—1,5 периодам голодания мелких лабораторных животных. Сроки периодов питания равны срокам периодов голодания. В наших наблюдениях мы вынуждены были объединить I и II периоды питания из-за недостатка данных, полученных в эти сроки восстановления.
СОБСТВЕННЫЕ ДАННЫЕ
Согласно полученным данным, исходные (до голодания) цифры эритроцитарного состава периферической крови у всех обследованных лиц колебались в пределах нормальных величин.
В течение всего срока как полного голодания, так и последующего литания, среднее содержание эритроцитов в 1 мм3 периферической крови не отличалось от фонового уровня (Р>0,1). В первые 4 дня голодания в 9 случаях наблюдалось увеличение числа эритроцитов на 180000—1800000 b 1 мм3. Такое же увеличение было отмечено в 3 других случаях в сроки с 5 по 12 дни и в 5 случаях в сроки с 13 по 40 дни голодания. И лишь в 3 наблюдениях к 25—40 дням голодания число эритроцитов снизилось на 420000—800000 по сравнению с исходными данными. В первые 2—4 дня после прекращения голодания в 15 наблюдениях отмечалась кратковременная эритропения по сравнению с последними днями голодания (на 39000—870000 в 1 мм3), которая исчезала уже в течение двух первых недель восстановления.
Столь же стабильным оказалось и среднее содержание-гемоглобина во все исследованные нами сроки голодания. ±0,8-3,4 г %.
Подобное соотношение привело к неопределенной вар-лабильности цветного показателя. Так, при монотонных средних данных (табл. 1) цветной показатель в отдельных случаях варьировал в пределах от +0,36 до —0,29 при наиболее частых колебаниях ±0,08—0,09. Проведенное нами математическое вычисление характера и плотности корреляционных отношений между содержанием эритроцитов в 1 мм3 крови и гемоглобином показало, что достоверная прямая, достаточно высокая корреляция между этими двумя параметрами обнаруживалась лишь в 11 наблюдениях (=+0,630±0,250; + 0,960±0,025). В 12 случаях корреляционные отношения были статистически неопределенными и в 1 — отрицательными.
Одной из наиболее устойчивых величин при голодании оказалась средняя концентрация гемоглобина в эритроцитах (СКЭ), которая не только не снижалась во все обследованные сроки голодания и питания (варьируя в пределах нормы от 32% до 36%), но и обнаруживала вероятную тенденцию к увеличению в III период голодания (0,05>Р>0,02).
Постоянство эритроцитарного состава периферической крови во время голодания вполне подтверждалось и изучением гематокрита крови, средние показатели которого не уменьшались ниже исходных цифр во все обследованные сроки. В I период голодания в 13 случаях отмечалось умеренное увеличение гематокрита (Р<Д05). Индивидуальные показатели гематокрита в период голодания колебались от 35% до 50% с максимальным варьированием ±7,0%. Умеренное снижение средней величины гематокрита отмечалось лишь в III периоде голодания (0,001<Р<0,01) (табл- 1).
Таблица 1
Эритроцитарный состав периферической крови при полном алиментарном голодании и последующем питании людей
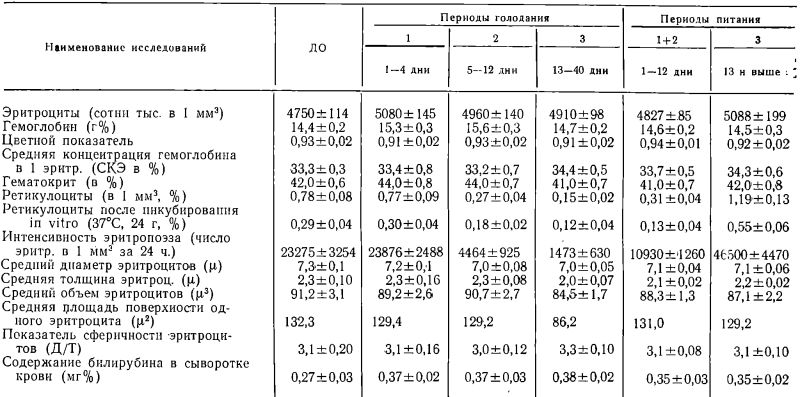
Данные по гематоириту позволили рассчитать средний объем эритроцитов. Последний не изменялся в течение двух первых периодов голодания и лишь в III его период статистически достоверно уменьшился (Р<0,05). После прекращения голодания средний объем эритроцитов вновь начал увеличиваться. Сопоставление динамики колебания среднего объема эритроцитов с числом эритроцитов в 1 мм3 крови показало, что в большинстве случаев между этими двумя величинами наблюдается при голодании Обратная зависимость: увеличение числа эритроцитов сопровождается уменьшением среднего объема эритроцитов, и, наоборот, что дает основание полагать, что стабильность гематокритного показателя во время голодания зависит как от концентрации эритроцитов, так и от их объема (см. рис. 1).
Геометрические параметры эритроцитов во все обследованные сроки также колебались в пределах физиологической нормы. Так, средний диаметр эритроцитов несколько уменьшился лишь во II и III периоды голодания (0,05>Р>0,01). Линейные размеры эритроцитов в большинстве случаев колебались в диапазоне от 5,5?. до 10?. Тенденция к микроцитозу сопровождалась и легким аиизоцитозом, наиболее отчетливым в I периоде голодания и уменьшавшимся по мере продолжения сроков голодания. В I период голодания на кривых Прайс-Джонса чаще всего обнаруживалось расширение основания, уплощение верхушки, иногда появление двугорбых вершин. В отдаленные сроки голодания (20—40 дни) основания кривых сужались. В восстановительный период отмечалось увеличение среднего диаметра эритроцитов и сдвиг вершин кривых Прайс-Джонса вправо (см. рис. 2).
Проведенные эритроцитометрические измерения позволили рассчитать как толщину (высоту) эритроцитов, так и определить их условную форму. Как можно видеть из табл. 1, средняя высота эритроцитов уменьшалась лишь в III периоде голодания и с началом питания вновь обнаруживала тенденцию к увеличению. Из сопоставления данных относительно диаметра и высоты эритроцитов можно видеть, что для III периода голодания чаще всего была характерна тенденция к микропланацитозу. В восстановительный период форма эритроцитов постепенно возвращалась к исходной. Тенденция к микропланоцитозу в III период голодания нашла свое отражение и в уменьшении площади поверхности одного эритроцита, которая вновь начала увеличиваться лишь в восстановительном периоде (табл. 1)- Поскольку изменению формы эритроцитов придают большое значение гори определении степени их «готовности» к деструкции, нами была предпринята попытка математически определить существование зависимости между изменением условной формы эритроцитов и их концентрацией в периферической крови. С этой целью был рассчитан коэффициент корреляции между показателями сферичности эритроцитов и их содержанием в 1 мм3 периферической крови. Расчеты показали, что прямая достоверная или вероятная корреляция отмечалась всего лишь в 9 наблюдениях (r=от +0,42±0,24 до +0,90±0,04), в одном случае она была достоверно отрицательной (r=—0,94 ±0,02) и в остальных неопределенной.
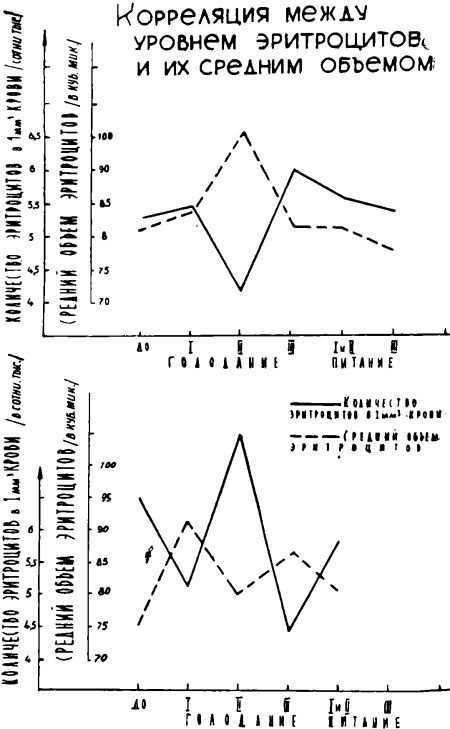
Рисунок 1.
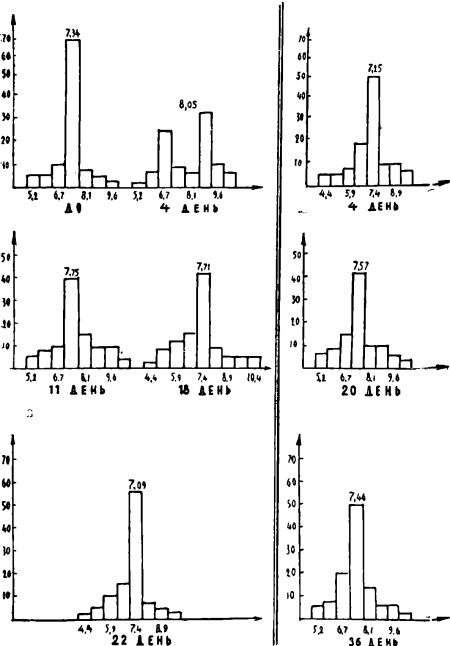
Рисунок 2.
Наряду с отмеченной во время голодания «устойчивостью» эритроцитарного состава в единице объема периферической крови отмечалось и неуклонное уменьшение числа ретикулоцитов. В отдаленные сроки голодания ретикулоцитопения становилась чрезвычайно выраженной (1—2 ретикулоцита в нескольких мазках крови). Однако полностью в сроки наших наблюдений ни в одном случае ретикулоциты из периферической крови не исчезали. Интересно также отметить, что в случаях с гемаконцентрацией ретикулоцитопения носила более выраженный характер. После прекращения голодания число ретикулоцитов начинало постепенно нарастать, а в ретикулоцитограммах отмечался сдвиг в сторону макроцитарных, грубозернистосетчатых форм. Максимальное повышение числа ретикулоцитов отмечалось не сразу же после превращения голодания, но спустя примерно 10—20 дней после начала питания, когда их число в 1,5—3 раза превосходило исходный уровень. Увеличение числа ретикулоцитов закономерно предшествовало или совпадало с подъемом эритроцитов. Нормализация показателей красной крови наступала спустя примерно 30—60 дней после прекращения голодания.
Интенсивность эритропоэза, высчитанная по формуле, учитывающей как уровень эритроцитов и ретикулоцитов, так и скорость превращения ретикулоцитов в эритроциты in vitro в целом изменялась параллельно динамике числа ретикулоцитов в периферической крови: резко снижалась во II и III периоды голодания и увеличивалась в восстановительном периоде (см. рис. 3).
Падение числа ретикулоцитов после инкубирования in vitro было менее выражено, чем до инкубирования крови (на 28% и 59% и соответственно на 68% и 81% во второй и третий периоды голодания), что дает основание полагать, что
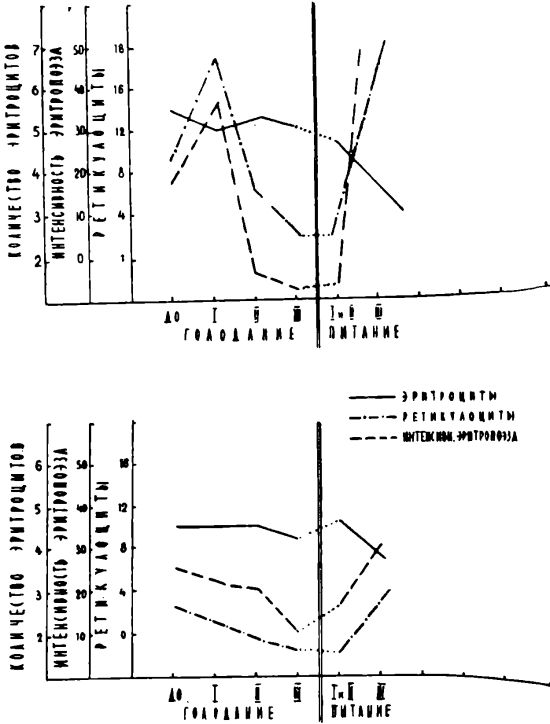
Рисунок 3.
В костном мозге при голодании не было отмечено учащения дегенеративных изменений в клетках эритробластического ростка. В парциальных эритробластограммах отмечался сдвиг вправо, процентное число ортохромных нормобластов превалировало над полихроматофильными (таблица 2).
Таблица 2
Парциальные эритробластограммы
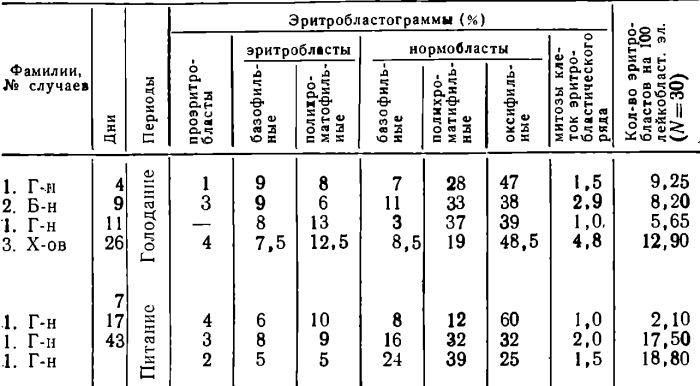
Обнаруживалось повышенное число ортохромных нормобластов с ядрами в различных стадиях пикноза. Митотический индекс для клеток эритробластического ряда оставался высоким, в то время как отношение числа эритфобл готических элементов к лейкобластическим существенно снижалось. Таким образом, при голодании наряду с устойчивым митотическим индексом наблюдалось существенное сужение фронта эритропоэза. Отношение ретикулоциты костного мозга—ретикулоциты периферической крови увеличивалось. После прекращения голодания указанные сдвиги продолжали наблюдаться и в исследовании на 7 день питания и лишь в более отдаленные сроки (17 и 43 дни восстановления) наблюдалось расширение фронта эритробластического кроветворения, сдвиг влево в парциальных эритробластограммах, увеличение числа митозов и другие признаки усиления эритропоэза примерно совпадающие по срокам с ретикуладитозом в периферической крови.
ОБСУЖДЕНИЕ
Наибольший интерес в исследовании эритрона при голодании представляет вопрос о механизмах поддержания относительного постоянства эритроцитарного состава периферической крови на фоне одновременного снижения интенсивности эритропоэза.
Большинство авторов решающее значение в этом отводят сгущению крови, полагая, что именно плазморреей гори голодании регулируется уровень эритроцитов в периферической крови.
Полученные нами данные при их сопоставлении с литературными источниками позволили предположить, что регуляция относительного постоянства эритроцитарного состава периферической крови обусловливается, во-первых, возрастающей депонацией крови и, во-вторых, относительной стабильностью процесса эритродиереза (т. е. либо сохранением его на уровне, имевшем место до голодания, либо меньшей, по сравнению с процессом эритропоэза, падением его интенсивности).
Как уже говорилось, в экспериментальных работах имеются указания на перераспределение массы крови в пользу депо (побледнение кожных покровов, стойкая гиперемия внутренних паренхиматозных органов и т. д.) (15, 19, 44, 45), более постоянно наблюдаемая, начиная со средних сроков голодания. Известно также, что во время полного (голодания не прекращается процесс желчеобразования (14, 19, 49—54), а органы ретикулогистиоцитарной системы содержат большое количество гемосидарина (15).
Зная, что при полном голодании масса крови изменяется примерно параллельно падению веса тела, и учитывая, что в норме в течение суток разрушается около 0,6—0,8% всех эритроцитов, можно легко рассчитать, что, например, за 40 дней полного голодания (при допущении сохранности обычного для физиологических условий объема и темпа эритродиереза) должно разрушиться около 24—32% всей эритроцитарной массы. При этом должно сохраниться нормальное (или близкое к нему) соотношение между объемом эритроцитов и плазмы, как это и имело место в наших наблюдениях. Если учесть, что голодающий организм человека теряет за этот же период примерно 20—26% веса тела к исходному, то мы получаем хорошее соотношение между массой эритроцитов и «живой массой тела». При этом следует напомнить, что эритропоэз в наших наблюдениях, постепенно ослабевая, не угасал окончательно даже в отдаленные сроки голодания и, следовательно, некоторая часть эритроцитов (ретикулоцитов) поступала из костного мозга в периферическое русло.
Таким образом, можно полагать, что основная направленность регуляции эритроцитарной системы при полном голодании состоит в предотвращении возможности возникновения вторичной полицитемии (устранение эритроцитарных «излишков»), что и достигается снижением уровня эритропоэза, перемещением («выведением») части крови во «внутренний цилиндр» организма с последующим разрушением эритроцитов в соответствующих органах и тканях. Подобное предположение ни в коей мере не исключает участия в регуляции относительного постоянства красной крови таких факторов как плазморрея и пр., которые, однако, в данном случае должны рассматриваться как второстепенные.
По-видимому, функциональная депрессия эритропоэза (одной из причин которой, как уже говорилось, является снижение эритроггоэтической активности крови) затрагивает вначале процесс элиминации эритроцитов из костного мозга в периферическое русло, что, видимо, и приводит к увеличению отношения ретикулоциты костного мозга — ретикулоциты периферической крови. Можно также полагать, что при полном голодании имеет место замедление сроков созревания нормобластов, о чем в наших наблюдениях свидетельствовало преобладание оксифильных форм с пикнотизированными ядрами. Этим же феноменом можно, вероятно, объяснить и тенденцию к микроцитозу эритроцитов при голодании. Сужение «фронта эритропоэза» также является одним из отражений снижения эритропоэтической активности.
С другой стороны, есть основания полагать, что описанные механизмы являются факторами, способствующими сохранности функциональных качеств регенерируемых эритроцитов (например, стабильная концентрация гемоглобина в эритроцитах — СКЭ, высокие митотаческие коэффициенты).
Несомненно, что все вышеуказанные предположения подлежат проверке (изучение функциональной активности РГС при голодании, исследование сроков жизни эритроцитов, их резистентности по отношению к различным гемолитикам и т. п.).
Следует здесь же подчеркнуть, что отчетливые нарушения эритропоэза наблюдаются лишь при более значительных сроках голодания, чем это имело место в наших наблюдениях (потери веса тела на 30% и более к исходному) и описаны в основном при экспериментальном голодании животных (14, 16, 17, 34, 35).
Восстановительный период характеризуется усилением активности эритропоэтической функции с последующей нормализацией всех показателей периферической красной крови.
Рассматривая все изменения в полностью голодающем организме как проявление приспособления (84—87), можно полагать, что, изучая эритрон, мы имеем дело с частным случаем адаптационной реакции организма к голоданию. Причем для эритрона в условиях и в сроки наших наблюдений следует принять сохранность компенсаторных механизмов, обеспечивающих эффект его адаптации. С точки зрения биокибернетики при рассмотрении поведения системы эритрон — «целью» описанных регуляторных механизмов является как поддержание достаточного количества и сохранение функциональной активности (дыхательной, транспортной, выделительной и т. п.) универсальной физиологической системы красной крови на уровне, необходимом для поддержания жизни, так и энергичная репарация (переадаптация) этой системы при возобновлении питания.
ЛИТЕРАТУРА
1. Спасский И. Г. Военно-мед., журнал, Ш34, 23, № 2, 457.
2. Buchinger О. Das Heilfasten, Stuttgart—Leipzig, 1941.
3. Бурштейн M. Д. Невропсихология и пспхиатрия, 1947, 16, Хя 6, 73.
4. Н е u n Е. Das Fasten als Erlebnis und Geschehnis. Frankfurt am Main, 1953.
5. Николаев Ю. С. Разгрузочно-диетическая терапия шизофрении и ее физиологическое обоснование. Дисс. докт. М., 1959.
6. Бакулев А. Н. и Колесникова Р. С. Клинич. медицина, 1962, № 2, 14.
7. Kraufi Н. u. Hartmann К. Archiv f. physik. Therapie, 1964, Marz/April, N. 2, 109.
8. S a b e h G., В о n e s s i J. V., S а г v e г M. E. et al. Metabolism, 1965, 14, N. 5, 603.
9. LecocqR, McPhaul J. J. Metabolism, 1965, 14, N. 2, 186.
10. R a t h R., M a s e k J., В u 11 n e г о v a H. O. Ceskosl. gastro-erferol. a. vyziva, 1966, 20, N. 2, 175.
11. Drenick E. J, Blahd W. H., Singer F. R., Lederer M, Amer. J. Clin. Nutr., 1966, 18, N. 4, 278.
12. Schultz С. H. Ueber den Zustand des Blutes in einem verhun-geiten Proteus, sowie in verhungerten Katzen und Kaninchen. In: Beitrage zur physiologischen und pathologischen Chemie und Mikroskopie in ihrer Anwendung und die praktische Medizin. Berlin, Von Dr. Franz Simon, 1844, 1, 820.
13. Panum P. Z, V i г с h. Arch, pathol. Anat., Physiol., Klin. Med., 1864, 29, 241.
14. Каган Я. А. Кровь и кровяное давление у голодающих. Дисс. СПб, 1884.
15. Сольц О. С. Об анатомических изменениях костного мозга при полном голодании и последующем откармливании. Дисс. Вильна, 1894.
16. Агамова К. А. Костномозговое кроветворение при голодании и влияние на него внутривенных вливаний видовонеспецифической сыворотки. Дисс. канд., М., 1954.
17. М а ч а в а р и а н и Э. Н. Морфологические изменения периферической крови и кроветворного аппарата при экспериментальной алиментарной дистрофии. Дисс. канд., Тбилиси, 1952.
18. Зайцева Р. А. Регенерационные процессы в костном мозгу у молодого организма в норме, при голодании и парентеральном питании. Дисс. канд., М., 1956.
19. Benedict F. A study of prolonged fasting. W., 1915.
20. Leumann Mueller, Munk, Senator a. Zuntz. Virch. Arch. f. pathol. Anat. u. Physiol., 1893, 131, Beiheft 1, 1.
21. L u с i а п i L. Der Hunger. 1890.
22. Архангельский С. В. Врачебное дело, 1926, 8, 729.
23. П р у с с к и ft В. Ф. и Ш у с т е р о в Г. А. Мед журнал. М., 192а, Но 5.
24. Гефтер Ю. М. и Юделович Р. Я. Ж. Физиологии СССР, 1934, 17, И 2, 287.
25. S с h е n k Е. G. u. М eye г Н. Е. Das Fasten. Stuttgart-Leipzig, 1938.
26. Egoroff A., Gall W. I., L a p t e w a - P о p о w a M. S„ Wassiljewa E. S. Folia Haematologica, Leipzig, 1935, 53, H. 2, 155.
27. Kanzow U. Dtsch. Arch. klin. Med., 1951, 198, No. 6, 698.
28. Шапиро Ю. Л. Ж. Бюлл. эксперим. биологии и мед., 1963, 5, 40.
29. 3 о р и н Е. Н. В кн.: Сб. работ по физиологии, М., 1939. с. 198.
30. Р я б у ш и и с к а я Н. П., Ежова Е. Н., Зорин Е. Т., Ильин-Кукуев Б. И., Молчанова О. П., Неумолот о-в а Е. А., Редина Л. В., Алексеев М. В. В кн.: Труды 3-го Всесоюзного съезда физиологов. Л., 1928, т. 1, с. 227.
31. Кадыров И. Г. Научные труды Башкирского мединститута, 1940, 3; 56.
32. Касимов В. Б. Влияние полного голодания на реактивность организма в зависимости от его различных функциональных состояний. Дисс. канд., Баку, 1962.
33. W h i t е R. R., С h г i s t i a n k К. В., W i 11 i a m s V. J. New Zealand J. Sci. a. Technol., 1956, A-38, No. 4, 440.
34. Любомудров П. В. Изменение крови и некоторых органов крови при голодании. Дисс. СПБ, 1893.
35. Полетаев П. И. Морфологический состав крови при полном и неполном (с водой) голодании у собак. Дисс. ОПБ, 1894.
36. Раrеirа М., Sic he г N.. Lang S. Arch. Surg., 1958, 77, No. 2, 191.
37. H u r s с h f e 1 d W. J., Gordon A. S. Anatom. Rec., 1965, 153, No. 3, 317.
38. Смирнова Л. И. Докл. АН СССР, 1965, 160, № 1, 240.
39. Hansen P., S с a n d. J. Clin. a. Lab. Invest., 1963, 15, No. 4,
40. D о n a f i R. M., G h a p m a n C. N.t W а г n e с k e M. A., Gallagher N. J. Proc. Soc. Exptl. Biol. a. Med.f 1964, 117, No. 1, 50.
41. P a natt a G. В., Gaeta M, Fiorentini M, L e о 11 a R. Boll. Soc. ital. biol. sperim., 1966, 42, No. 11, 681.
42. Солодюк H. Ф. Восстановление белкового и морфологического состава крови после кровопотери и голодания (экспериментальное исследование). Дисс. докт., Киев, 1958.
43. Voit С. Zeitschr. f. Biologie, 1866, 2, No. 3, 309.
44. Пашутин В. В. Курс общей и экспериментальной патологии, 1902, СПБ, т. 2, ч. 1.
45. Перельман Р. Л. Руководство по патологической физиологии. Под редакцией А. А. Богомольца, Киев, 1947, т. 2, ч. 2, гл. XII, с. 89.
46. С a s а 1 P. Erythrocytes et erythropathies. Paris, 1957.
47. Ponder E. Hemolysis and related phenomena, N. Y., 1948.
48. Ч и ж e в с к и й А. Л. Структурный анализ движущейся крови. М., 1959.
49. П о б е р и й И. А. Процесс регенерации в селезенке и лимфатических узлах при парентеральном введении белка в условиях голодания. Дисс. канд., М., 1956.
50. Л а н д у а Л., Р о з е м а н Р. Учебник физиологии человека. М., 1913, 94, 382, 389.
51. Лукьянов С. М. Изв. Варшавск. ун-та, 1891, 7, 1—16, 17.
52. М а г t i g n i С. de u. N a s s e. Ueber den Einflup der Nahrung auf das Blut. Mtrburg u. Leipzig, 1850.
53. О x о т и н И. А. Патологоанатомические изменения и газовый обмен у голодающих кроликов. Дисс, GFIB, 1885.
54. О к и н ч и ц Е. С. К вопросу об изменении морфологического состава крови у кроликов при полном голодании и последующем откармливании. Варшава, 1893.
55. Lambiotte М, Bull. Soc. chim. biol., 1958, 40, No. 11, 1579.
56. К i ni К- S., Bollman J. L, G r i n d 1 а у J. H. Amer. J. Physiol., 1956, 184, No. 3, 445.
57. G г u п e г P., Heni F, M a s t H. Acta Histochem, 1959, 7, No. 5—8, 199.
58. Ярошевский А. Я. Ж. высш. нервн. деятельности им. И. П. Павлова, 1954, 4, вып. 5, 674.
59. Поберий И. А. Цитология, 1964, 6, JNfe 3, 330.
60. Ш а п и р о Ю. Л. Состояние системы крови при длительном полном алиментарном голодании и последующем питании людей. Дисс. канд., М. 1964.
61. Hodgson G., Yudilevich D., Perretta M., Esku-che I., Tohai J Ann. N. Y Acad. Sci., 1955, 77, No. 3, 703.
62. С arm en a A., Garcia N. E., Aggio M. C. Rev. Soc. argent, biol., 1963, 39, No. 1—4, 104.
63. A b b г e с h t P. H., M a 1 v i n R. L. Amer. J. Physiol., 1966, 210, No. 2, 237.
64. Y u n i s S. L., В e г с о v i t с h D. Danny, S t i e n R. M. J. Clin. Investig., 1964, 43, No. 8, 1668.
65. Hodgson G., JudilevichD., Eskuche J., Perret-t a M. Proc. Exptl. Biol, and Med., 1960, 104, No. 3, 438.
66. N a k а о Kiku, Нихон кэцуэки гаккай дзасси, Acta haemathol. japon., 1962, 25, No. 3—4, 253.
67. E s k u с h e I., H о d g s о n G. Proc. Soc. Exptl. Biol. and Med., 1966, 122, No. 2, 437.
68. Mann D. L., Donati R. M., G a 11 a g h e r N. J. Proc. Soc. Exptl. Biol. and Med, 1966, 121, No. 4, 1152.
69. Арутюнян P. А. Ж. эксперим. и клииич. мед., 1964, 4, № 3, 19.
70. Z i 1 ke г s m i t D. В., Bollman J. L. Arch. Boichem. and Biophys, 1956, 63, No. 1, 64.
71. Cordon A. S., W i n k e r t J, Dornfest B. S, S i e g e 1 C. D. Ann. N. Y. Acad. Sci, 1959, 77, No. 3, 650.
72. G о 1 d w a s s e r E, Jacobson L. O., Fried W, Plzak L. Mechanism of the erythropoietic effect of cobalt, Science, 1957, 125, No. 3257, 1085.
73. H о d g s о n G, Perretta M, Yudilevich D., Eskuche I. Proc. Soc. Exptl. Biol. and Med, 1958, 99, No. I, 137.
74. Mc Carthy G. M, Gallagher N.J, Lange R. D. Metabolism, 1959, 8, No. 4, Part 1, 429.
75. Gurney C. N, Pan С h a o. J. Lab. and Clin. Med, 1960, 55, No. 1, 67.
76. Sсаrо J. L, К f i g h I e у G, L о w у P. H. Acta physiol.
77. Танака Ходэо. Нихон кэцуэки гаккай дзасси, lation-amer., 1963, 13, No. 4, 362.
Acta haematol. japon, 1964, 27, No. 4, 505.
78. Thorling E. B. Scand. J. Haematol, 1965, 2, No. 1, 36.
79. Scaro S. L. Acta physiol. latino-amer, 1965, 15, No. 2, 200.
80. Gurney С W., F г i e d W. J. Lab. and Clin. Med, 1965, 65, No. 5, 775.
61. Ш а п и р о Ю. Л. В кн.:- Материалы конференции по патологии клетки. М., 1967, с. 253.
82. Т о д о р о в И. Клинические лабораторные исследования в педиатрии. София, 1963.
83. Кост Е. А. В кн.: Руководство по внутренним болезням. М., 1962, т. 6, с. 56.
84. Попов Ю. П. К вопросу изучения скорости созревания ретикуляцитов и продукции эритроцитов костного мозга, Дисс. канд., Казань, 1961.
85. Селье Г. Бюллетень эксперим. биол. и мед. 1909, в, 5, 11, 339.
86. Федоров Н. А. Предисловие к книге: «Лечение шизофрении дозированным голоданием». Труды Гос. НИИ психиатрии МЗ РСФСР, М., 1963, т. 93, с. 5.
87. Давыдовский И. В. Проблемы причинности в медицине (этиология). М., 1962.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва)
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва) Лечение ожирения остается одной из наиболее актуальных проблем современной медицины. Число больных с избыточным весом, согласно данным многих
К характеристике изменения ЭЭГ у больных, находящихся на длительном лечебном голодании В. В. АРШАВСКИЙ, Б. В. КРАЙЦЕРОВ (Москва)
К характеристике изменения ЭЭГ у больных, находящихся на длительном лечебном голодании В. В. АРШАВСКИЙ, Б. В. КРАЙЦЕРОВ (Москва) Из многочисленных данных литературы известно, что центральный отдел висцерального анализатора как у животных, так и у человека проецируется в
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва)
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва) Изучению реакции организма на длительное голодание посвящено много работ. Большинство из них касается обменных процессов, биохимии синтеза и распада
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва)
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва) Как известно, при полном алиментарном голодании в течение довольно длительного времени (по крайней мере до сроков, характеризующихся потерями
Сывороточное и эритроцитарное железо при длительном голодании людей Ю. Л. ШАПИРО, Л. М. ДОНДЫШ, Л. М. ЛЕЙБИН, Э. А. ЛЕЙЗЕРОВИЧ, Л. Ф. ЛЕВИНА (Москва)
Сывороточное и эритроцитарное железо при длительном голодании людей Ю. Л. ШАПИРО, Л. М. ДОНДЫШ, Л. М. ЛЕЙБИН, Э. А. ЛЕЙЗЕРОВИЧ, Л. Ф. ЛЕВИНА (Москва) Данные о содержании железа в жидкой фракции крови при полном алиментарном голодании и последующем питании немногочисленны и
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва)
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва) В литературе имеется значительное число работ, посвященных изучению изменения степени насыщения крови кислородом при разнообразных условиях и
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва)
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва) Изучению фагоцитарной активности лейкоцитов при длительном полном алиментарном голодании посвящены
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва)
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва) Известно, что так называемые «жесткие константы» сохраняются неизменными в организме В течение длительного срока, несмотря на воздействие каких-либо «отклоняющих» факторов.
К вопросу о реакции различных форм ожирения на голодание Д. Д. ФЕДОТОВ, Ю. Л. ШАПИРО, Ф. А. ВАЙНДРУХ (Москва)
К вопросу о реакции различных форм ожирения на голодание Д. Д. ФЕДОТОВ, Ю. Л. ШАПИРО, Ф. А. ВАЙНДРУХ (Москва) Ожирение, как и вообще проблема неправильного питания, становится в последнее время все более актуальной.В то время, как большая часть человечества (по данным ВОЗа —
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва)
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва) Половой хроматин — внутриядерное тельце, дающее Фельген-положительную реакцию и
О динамике основного обмена при лечебном голодании больных с ипохондрическим синдромом В. Б. ГУРВИЧ (Москва)
О динамике основного обмена при лечебном голодании больных с ипохондрическим синдромом В. Б. ГУРВИЧ (Москва) Вопрос об обмене веществ, в частности, основном обмене, является одним из ведущих при объяснении патофизиологических механизмов голодания (1, 5, 9, 11, 12, 14).В. В.
СОСТОЯНИЕ ИММУНОБИОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ ОРГАНИЗМА ЧЕЛОВЕКА ПРИ ПОЛНОМ ДЛИТЕЛЬНОМ ГОЛОДАНИИ Ю. С. НИКОЛАЕВ, В. А. СКОРИК-СКВОРЦОВА (Москва)
СОСТОЯНИЕ ИММУНОБИОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ ОРГАНИЗМА ЧЕЛОВЕКА ПРИ ПОЛНОМ ДЛИТЕЛЬНОМ ГОЛОДАНИИ Ю. С. НИКОЛАЕВ, В. А. СКОРИК-СКВОРЦОВА (Москва) Изучение состояния иммунобиологической реактивности организма в период длительного полного голодания и последующего
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва)
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва) В течение последних лет в нашей стране и за рубежом появилось большое число сторонников применения голодания с лечебной целью при
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва)
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва) Проблема влияния голодания на разные показатели обменных процессов в организме животных и человека продолжает привлекать внимание
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва)
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва) Голодание как метод лечения успешно зарекомендовал себя при некоторых психических и соматических заболеваниях (3, 7, 10—13). Особенно перспективно его применение при обменных, аллергических