Влияние голодания на активность ферментов пентозофосфатного пути в печени и мозге крыс Ю. Л. ЗАХАРЬИН (Москва)
Влияние голодания на активность ферментов пентозофосфатного пути в печени и мозге крыс
Ю. Л. ЗАХАРЬИН (Москва)
В последние годы в клинике часто применяется с лечебными целями, в частности, для лечения психических заболеваний, полное голодание. Не вызывает сомнения, что применение такого сильного воздействия требует тщательного изучения последствий голодания для организма.
Как известно, пентозофосфатный путь окисления глюкозы является одним из наиболее важных путей углеводного обмена. Начальные реакции этого пути — окисление глюкозо-6-фосфата и 6-фосфоглюконата — приводит к образованию рибозофосфата, необходимого для синтеза нуклеотидов и нуклеиновых кислот и НАДФ-Н2, являющегося кофактором в целом ряде синтетических процессов, в частности, в синтезе жирных кислот, кортикостероидов, эстрогенов, аскорбиновой кислоты, о восстановлении рибонуклеотидов в дезоксирибонуклеотиды, в процессе органификации йода в щитовидной железе и т. п.
Ферменты, осуществляющие окисление глюкозо-6-фосфата и 6-фосфоглюконата — глюкозо-6-фосфат-дегидрогеназа (Г-6-Ф-Д; КФ 1.1.1.49) и 6-фосфоглюконат-дегидрогеназа (6-Ф-ГЛ-Д; Кф 1-1-1.44) — являются адаптивными. Было показано, что активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени зависит от характера диеты. Неоднократно исследовалось также влияние голодания на активность этих ферментов. Однако в большинстве работ (2, 5, 6, 10, И, 14—17, 19—22, 29, 30) были исследованы только короткие сроки голодания — 1—3 дня. Лишь в некоторых работах (12, 24— 26. 31) исследовались сравнительно длительные (для крыс) сроки голодания (5—6 дней). Влияние же голодания на активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в ткани мозга вообще не изучалось.
В настоящей работе исследовалось влияние голодания и возобновления кормления на активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени и двух функционально различающихся областях мозга (полушария и стволовая часть).
Методика
Работа проведена на крысах линии Вистер исходным весом 130—170 грамм. Перед началом опыта крысы в течение 10 дней содержались на синтетической сбалансированной, высокоуглеводной или малобелковой диете. Сбалансированная диета содержала (по калорийности) 18,1% казеина, 26,9% лярда, 55% крахмала; высокоуглеводная диета содержала 18,1% казеина, 5% лярда и 76,9% крахмала; малобелковая диета содержала 8% казеина, 30% лярда и 62% крахмала. К этому добавлялось также 5% (по весу) сухих дрожжей, 4% солевой смеси и необходимое количество витаминов А, Д и Е.
В процессе опыта часть крыс была лишена пищи в течение 5—10 дней и получала только воду; контрольные крысы получали ту же пищу вволю. Сроки голодания были предельно допустимыми и определялись выживаемостью крыс в каждом отдельном опыте. Обычно крысы голодали до тех пор, пока не начинали гибнуть; тогда оставшихся крыс забивали и определяли активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в тканях. Исследования, проведенные с более короткими сроками голодания (1—3 дня), не показали заметных изменений ферментной активности, поэтому эти результаты не приводятся. За время голодания крысы теряли 30—40% веса (по сравнению с контрольными крысами того же возраста). В опытах с возобновлением кормления крысы голодали 5 дней и затем вновь получали ту же пищу в течение 3 дней.
Ткани брались сразу после забоя на лед и гомогенизировались в охлажденном стеклянном гомогенизаторе с охлажденным 0,15 М раствором КС1 в соотношении 1 : 10. Гомогенат центрифугировали 20 минут при 4000 об/мин. и центрифугат разбавляли водой до нужной концентрации.
Активность Г-6-Ф-Д и 6-Ф-ГЛ-Д определялась опектрофотометрическим методом в нашей модификации (1).
Реакционная смесь имела следующий состав: 1 мл 0,3 М буфера+0,2 мл 1 М MgSO4 + 0,2 мл 0,002 М НАДФ + 0,2 мл гомогената+1 мл воды (в пустые пробы — 1,4 мл). Реакцию начинали добавлением 0,4 мл 0,002 М раствора субстрата (глюкозо-6-фосфата или 6-фосфоглюконата). В качестве контрольных к пробам с глюкозо-6-фосфатом были взяты пробы, в которые добавлялся 6-фосфоглюконат в концентрации 0,001 М (объяснение см. 1). Инкубация проводилась в течение 15 минут при 37°. Реакцию останавливали добавлением 1 мл 1,2 н NaOH. Выпадающий осадок Mg(OH)2, увлекающий за собой все взвешенные частицы, удалялся центрифугированием, и в прозрачном центрифугате определялась оптическая плотность при 340 м?.
Для определения активность Г-6-Ф-Д использовался глицил-глициновый буфер рН — 9,5, а для 6-Ф-ГЛ-Д — рН — 8,5.
Калибровочная кривая строилась по раствору НАДФ — Н2 в 0.1 н NaOH. Активность ферментов выражалась в наномолях НАДФ, восстановленного за 1 мин., на 1 мг растворимого белка.
РЕЗУЛЬТАТЫ
Как видно из таблицы 1, активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени заметно снижается при голодании. Это снижение более выражено у крыс, находившихся перед голоданием на высокоуглеводной диете. Активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в мозге также имеет тенденцию к снижению, но разница в этом случае статистически недостоверна.
Возобновление кормления после голодания сопровождалось выраженными морфологическими изменениями в печени. Вес печени значительно увеличивался (5,5% от веса тела против 3,8% в норме). Макроскопически и микроскопически у таких крыс найдена выраженная жировая инфильтрация печени.
Таблица 1
Активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени и мозге при голодании
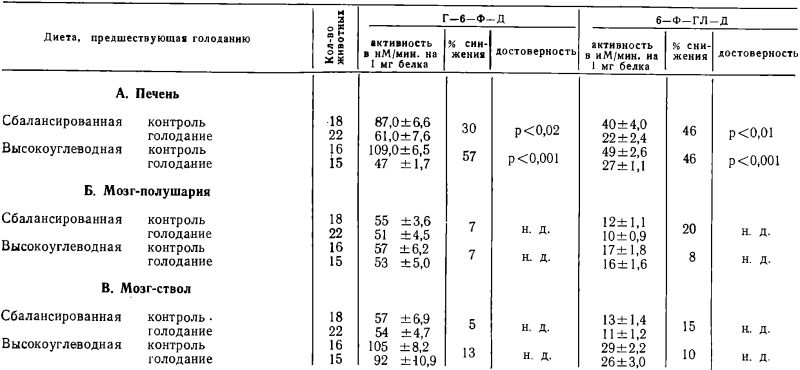
Таблица 2
Активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени и мозге при возобновлении кормления
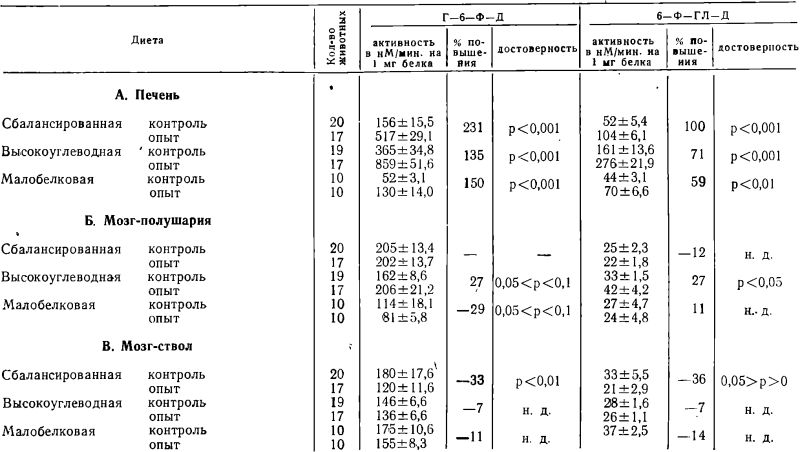
Таблица 3
Активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени и мозге крыс, находившихся на разных диетах

В таблице 2 показаны изменения активности Г-6-Ф-Д и 6-Ф-ГЛ-Д при возобновлении кормления крыс после голодания. Активность ферментов-в печени значительно возрастает после возобновления кормления в 2—3 раза, превосходя их активность в печени контрольных (не голодавших) крыс. Это возрастание наблюдается не только на высокоуглеводной и сбалансированной, но и на малобелковой диете. Активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в мозге после возобновления кормления находится в общем в пределах нормы. Небольшие отклонения в полушариях мозга на высокоуглеводной и малобелковой диете и в стволовой части мозга на сбалансированной диете, незакономерны и по-видимому случайны.
В таблице 3 сравнивается активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени и мозге не голодавших крыс, находившихся на различных диетах. Активность обоих ферментов в печени крыс, находившихся на высокоуглеводной диете, была выше, чем на сбалансированной. В печени крыс, находившихся на малобелковой диете, активность Г-6-Ф-Д была ниже, чем на сбалансированной; для 6-Ф-ГЛ-Д это снижение не было выражено.
Обсуждение
Большинство авторов (2, 7, 10-12, 20, 28) нашло при голодании небольшое снижение активности Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени или даже не нашло снижения (5, 23, 29, 30). Однако, в некоторых работах (4, 6, 8, 16, 24, 25, 31, 32) было обнаружено очень сильное уменьшение активности этих ферментов в печени голодавших крыс. Вероятно, эти разногласия можно объяснить различием предшествовавшей голоданию диеты. Как было показано (5), голодание после сбалансированной диеты не влияет на активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени, тогда как голодание после предшествующей диеты, богатой глюкозой, сопровождается снижением активности этих ферментов. К такому же выводу можно прийти и на основании наших опытов, в которых голодание после сбалансированной диеты вызывало менее выраженное снижение активности Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени, чем голодание после высокоуглеводной диеты.
Многочисленными работами (2, 8, 9, 12, 13, 19, 20, 22-2, 3) было показано, что активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени значительно увеличивается при возобновлении кормления после голодания. Величина этого повышения зависит от характера диеты в период возобновления кормления (2, 8, 9, 12, 13, 18); оно, как правило, более выражено на высокоуглеводной диете. В то же время возобновление кормления малобелковой диетой не вызывает увеличения активности этих ферментов в печени (2).
В наших исследованиях, возобновление кормления сопровождалось значительным повышением активности Г-6-Ф-Д и 6-Ф-ГЛ-Д не только на сбалансированной и высокоуглеводной, но и на малобелковой диете. Это можно объяснить сравнительно мягким характером белкового дефицита (8% белка в диете вместо 18% в норме), тогда как в опубликованных-ранее работах (2 и др.) был применен более жесткий малобелковый режим (3—5% белка в диете).
Полученные нами результаты показывают, что абсолютная величина повышения активности Г-6-Ф-Д и 6-Ф-ГЛ-Д после возобновления кормления оказывается большей на высокоуглеводной и меньшей на малобелковой диете, чем на сбалансированной, но это повышение (выраженное в процентах к контрольной активности) больше на сбалансированной диете (вследствие более низкой активности у контрольных крыс на сбалансированной диете, чем на высокоуглеводной).
Результаты сравнения активности Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени не голодавших крыс на различной диете соответствуют литературным данным (5, 6, 20, 21, 23).
Активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в обеих исследованных частях мозга не зависит от диеты и существенно не изменяется при голодании и возобновлении кормления.
Многочисленные исследователи, изучавшие углеводный обмен с помощью неравномерно меченной С14-глюкозы, единодушно утверждают, что пентозофосфатный путь окисления глюкозы в мозговой ткани полностью отсутствует. Однако найденная нами в ткани мозга выраженная активность Г-6-Ф-Д и 6-Ф-ГЛ-Д заставляет предположить, что и в углеводном обмене мозга пентозофосфатному пути принадлежит определенное место.
Полученные в настоящей работе результаты показывают, что даже при предельно допустимых сроках голодания активность исследованных ферментов в печени изменяется не очень резко и полностью нормализуется при возобновлении кормления. Активность же этих ферментов в мозге при голодании практически не изменяется. Вместе с тем, учитывая все сказанное выше, можно прийти к выводу, что при терапевтическом применении полного голодания необходимо учитывать характер питания больных как в период, предшествующий голоданию, так и в период возобновления питания после голодания.
Выводы
Активность Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени снижается при голодании; это снижение более выражено у крыс, получавших перед голоданием высокоуглеводную диету. Возобновление кормления после голодания вызывает значительное увеличение активности Г-6-Ф-Д и 6-Ф-ГЛ-Д в печени на сбалансированной, высокоуглеводной и малобелковой диете. Активность этих ферментов у неголодавших крыс на высокоуглеводной диете выше, а на малобелковой — ниже, чем на сбалансированной.
ЛИТЕРАТУРА
1. Захарьин Ю. Л. Лабор. дело, 1967, стр. 327.
2. Поттер В. Р., Оно Г. «Регуляторные механизмы клетки», М., 1964, стр. 429.
3. Вlumen thai М. D., Abraham S., Chaikoff I. L. Arch. Biocem. a. Biophys., 1964, v. 104, N 2, p. 215.
4. Carlson J. R., Suttie J. W. Amer. J. Physiol., 1966, v. 210, N 1, p. 79.
5. Fitсh W. M., Chaikoff I. L. Arch. Biochem. a. Biophys., 1961, v. 94, N 3, p. 380.
6. Freedland R. A., Barnes J. K. J. Biol. Chem., 1963, v 238, N 6, p. 1915.
7. Glосk G, E, Mc Lean P. Biochem. J., 1955, v. 61, N 3, p. 390.
8. Jоhnsоn В. C., Moser K-, S a s s о о n H. F. Proc. Soc. Exp. Biol. a. Med., 1966, v. 121, N 1, p. 30.
9. Кagawa Y., Kagawa A., Shimazono N. J. Biochem., 1Я34, v. 56, N 4, p. 364.
10. КnocheI., Hartmann F. Bioch. Z., 1961, B. 334, H. 3, S. 269.
11. Lee M., Debrо J. R., Lucia S. P. Arch. Biochem. a. Biophys., 1962, v. 98, N 1, p. 49.
12. Mangiarоtti G., Сalabria G. A. Boll, soc, ital. biol. sierim., 1964, t. 40, N 7, p. 265.
13. Niemeyer H, Сlаrk-Turri L., Garcez E., Vаrgara F. E. Arch. Biochem. a. Biophys., 1962, v. 98, N 1, p. 77.
14. Niemeyer H., Perez N., Garcez E., Vergara F. E. Bioch. Bioph. Acta, 1962, v. 62, N 2, p. 411.
15. Оno Т., Potter V. R. Feder. Proc., 1961, v. 20, N 1, part 1, p. 224.
16. Pande S. V., Khan R. P., Venkitasubramanian T. A. Bioch. Bioph. Acta, 1964, v. 84, N 3, p. 239.
17. Potter V. R., Ono T. Cold Spring Symposia in Quantitative Biology, 1961, v. 26, p. 355.
18. Shimazono N. U. S. Departm. Com. Office Techn. Serv., -962—63, AD 432391, p. 21.
19. Tepperman H. M., Террегшап J. Diabetes, 1958, v. 7, N 6, p. 478.
20. Tepperman H. M., Tepperman J. Amer. J. Physiol., 1962, v. 202, N 3, p. 401.
21. Tepperman H. M., Tepperman J. Amer J. Physiol., 1964, v. 206, N 2, p. 357.
22. Tepperman H. M., Tepperman J. Amer J. Physiol., 1958, v. 193, N I, p. 55.
23. Vaughan D. A., Winders R. L. Amer. J. Physiol., 1964, v. 206, N 5, p. 1081.
24. Weber G., Ash mo re J., В a n e г j e e G. Feder. Proc., 1960, v. 19, N 1, part 1, p. 50.
25. Weber G., Banerjee G., Bixler D., Ash mo re J. J. Nutrition, 1961, v. 74, N 2, p. 157.
26. Weber G., Banerjee G., В г о n s t e i n S. B. Bioch. Bioph. Res. Comm., 1961, v. 4, N 5, p. 332.
27. Weber G., Banerjee G., Bronstein S. B. Amer. J. Physiol., 1962, v. 202, N 1, p. 137.
28. Weber G., Cantero A. Rev. Canad. Biol., 1956, t. 15, N 3, p. 290.
29. Weber G., Cantero A. Cancer Res., 1957, v. 17, N 10, p. 995.
30. Weber G., Cantero A. Amer J. Physiol., 1957, v. 190, N 2, p. 229.
31. Weber G., Mc Donald H. Exp: cell. Res., 1961, v. 22, p, 292.
32. Willmer J. S. Canad. J. Biochem a. Physiol., 1960, v. 38, N 12, p. 1449.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Влияние сенсорной стимуляции на двигательную активность эмбриона
Влияние сенсорной стимуляции на двигательную активность эмбриона Выше приводились примеры рефлекторных движений зародышей, производимых преимущественно в ответ на тактильные раздражения. Сенсомоторная активность составляет единый процесс на всех этапах жизни
Развитие идей лечебного голодания Ю. С. НИКОЛАЕВ (Москва)
Развитие идей лечебного голодания Ю. С. НИКОЛАЕВ (Москва) Можно считать, что люди еще с доисторических времен применяли голодание с лечебной целью; это подтверждается наблюдениями за животными, которые при заболевании отказываются от приема пищи, этому же инстинктивно
Клинико-электроэнцефалографичские корреляции у больных шизофренией с вялым течением в процессе лечебного голодания и последующего питания Б. В. КРАЙЦЕРОВ (Москва)
Клинико-электроэнцефалографичские корреляции у больных шизофренией с вялым течением в процессе лечебного голодания и последующего питания Б. В. КРАЙЦЕРОВ (Москва) Резистентность некоторых групп больных шизофренией с вялым течением ко многим терапевтическим
О динамике основного обмена жак отражении общей реактивности в процессе лечебного голодания психически больных В. Б. ГУРВИЧ, Г. И. БАБЕНКОВ (Москва)
О динамике основного обмена жак отражении общей реактивности в процессе лечебного голодания психически больных В. Б. ГУРВИЧ, Г. И. БАБЕНКОВ (Москва) В настоящее время в литературе встречаются указания «а расстройства обмена веществ при различных психических заболеваниях
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва)
Опыт лечения тучности методом полного голодания Д. Д. ФЕДОТОВ, Ю. С. НИКОЛАЕВ, Ю. Л. ШАПИРО, Г. И. БАБЕНКОВ, В. Б. ГУРВИЧ (Москва) Лечение ожирения остается одной из наиболее актуальных проблем современной медицины. Число больных с избыточным весом, согласно данным многих
Случай применения лечебного голодания у тучного больного, перенесшего инфаркт миокарда М.И. МИНЯЙЛЕНКО (Москва)
Случай применения лечебного голодания у тучного больного, перенесшего инфаркт миокарда М.И. МИНЯЙЛЕНКО (Москва) В литературе нам встретились ряд работ, указывающих на эффективность кратковременного воздержания от приема пищи при целом ряде соматических заболеваний.Так
О влиянии дозированного голодания на состояние слизистой желудка М. И. МИНЯЙЛЕНКО, А. И. БУХМАН, И. И. ЛУЧИНСКИЙ (Москва)
О влиянии дозированного голодания на состояние слизистой желудка М. И. МИНЯЙЛЕНКО, А. И. БУХМАН, И. И. ЛУЧИНСКИЙ (Москва) Голодание нередко приводит к изменениям со стороны желудочно-кишечного тракта. По данным Ю. С. Николаева, вторая стадия голодания сопровождается
К вопросу о роли дозированного голодания в комплексной терапии больных язвенной болезнью желудка и 12-перстной кишки М. И. МИНЯЙЛЕНКО (Москва)
К вопросу о роли дозированного голодания в комплексной терапии больных язвенной болезнью желудка и 12-перстной кишки М. И. МИНЯЙЛЕНКО (Москва) Язвенная болезнь представляет собой страдание, при терапии которого необходимы совместные усилия терапевтов, хирургов,
Влияние дозированного голодания на показатели проб Торна у больных шизофренией О. Я. СИЛЕЦКИЙ (Ростов-на-Дону)
Влияние дозированного голодания на показатели проб Торна у больных шизофренией О. Я. СИЛЕЦКИЙ (Ростов-на-Дону) Давно замечено, что эффект от лечения больных шизофренией с затяжным течением наступает тогда, когда меняется реактивность организма (2, 13 и др.). Применяя лечение
Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва)
Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва) Успехи, современной нейрофизиологии, связанные с использованием новых тонких методов электрофизиологического исследования мозга, позволили подойти к изучению центральных механизмов
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва)
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва) Изучению фагоцитарной активности лейкоцитов при длительном полном алиментарном голодании посвящены
Условнорефлекторные лейкоцитарные реакции в динамике лечебного голодания при некоторых нервно-психических заболеваниях С. М. ЛУРЬЕ (Москва)
Условнорефлекторные лейкоцитарные реакции в динамике лечебного голодания при некоторых нервно-психических заболеваниях С. М. ЛУРЬЕ (Москва) Учение И. П. Павлова «Об организме, как о целостной системе, как о функциональном единстве всех органов и тканей, психического и
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва)
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва) Проблема влияния голодания на разные показатели обменных процессов в организме животных и человека продолжает привлекать внимание
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва)
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва) В связи с тем вниманием, которое в настоящее время уделяется роли лизосом в развитии повреждения клеток тканей различных
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва)
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва) Голодание как метод лечения успешно зарекомендовал себя при некоторых психических и соматических заболеваниях (3, 7, 10—13). Особенно перспективно его применение при обменных, аллергических