Восходящие активирующие влияния на кору головного мозга у голодных животных К. В. СУДАКОВ (Москва)
Восходящие активирующие влияния на кору головного мозга у голодных животных
К. В. СУДАКОВ (Москва)
В последние годы благодаря применению новых методов физиологического эксперимента (электронные усилители, стереотаксическая техника) проблема голода, пищевого поведения и насыщения прочно заняла свое место среди основных проблем современной нейрофизиологии.
Установлено, что центральным пунктом формирования пищевых реакций животных и человека является гипоталамическая область. Так обнаружено, что при разрушении области вентромедиальных ядер гипоталамуса у животных возникает гиперфагия и на этой основе — ожирение. Наоборот, разрушение области латеральных гипоталамических областей вызывает афагию и гибель экспериментальных животных от истощения.
На основании этих и подобных им экспериментов было сформулировано представление о наличии в области латерального гипоталамуса так называемого «центра питания», а в области вентромедиальных ядер гипоталамуса— «центра насыщения» (17).
Вместе с тем в формировании пищедобывательного поведения первостепенная роль безусловно принадлежит клеткам коры головного мозга. Именно клетки коры головного мозга осуществляют синтез возбуждений, обусловленных воздействием на организм внутренней и внешней среды, производят их оценку и выработку своеобразного «решения» для совершения целенаправленного пищедобывательного акта. В связи с этим вопрос о том, на основе каких механизмов у голодных животных строятся функциональные взаимоотношения подкорковых пищевых центров с корой головного мозга, является наиболее важным для понимания природы пищедобывательного поведения. Однако прямого ответа на этот вопрос в современной научной литературе не имеется.
Открытие восходящих активирующих влияний ретикулярной формации на кору головного мозга (20, 24, 25 и др.) позволило исследовать эту проблему в новом аспекте. Стало ясным, что возбуждение может распространяться к коре не только локально в определенные пункты по специфическим лемнисковым путям, но и генерализованно ко всем отделам через активирующие аппараты ретикулярной формации. Эти восходящие активирующие влияния ретикулярной формации на кору мозга многими авторами рассматривались как неспецифические. Такое представление возникло на основе факторов, показывающих, что раздражение любой сензорной модальности вызывает однотипные электроэнцефалографические изменения, проявляющиеся в форме реакции, известной под названием «реакции десинхронизации», «активации», «блокады альфа-ритма» и т. п.
Вместе с тем, исследования лаборатории П. К. Анохина показали, что каждая биологически важная реакция организма строится на основе своих специфических механизмов восходящих активирующих влиянии ретикулярной формации на кору мозга.
Так было установлено, что активация коры мозга при болевой п пищевой реакции определяется различными в химическом отношении активирующими элементами ретикулярной формации. Оказалось, что болевая реакция» а также оборонительное состояние животного избирательно подавлялись введением аминазина (хлор-промазина), который согласно общераспространенному взгляду, блокирует преимущественно метаболизм ростральной части ретикулярной формации (2, 7). Тем не менее введение аминазина не оказывало заметного действия на проявление пищевой реакции (1, 5, 6, 11, 16).
Дальнейшее подтверждение эта точка зрения нашла в исследованиях, показавших, что различные наркотические вещества также избирательно действуют на системы корково-подкорковых взаимоотношений, вовлекаемых в формирование реакции того или иного биологического качества.
Так было показано, что такое наркотическое вещество как нембутал почти полностью блокирует все формы восходящих активирующих влияний со стороны подкорковых образований на кору мозга. В то же время другое наркотическое вещество — уретан, блокируя активацию коры, связанную с состоянием бодрствования, допускал развитие корковой активации вслед за нанесением болевого раздражения (I).
Все эти наблюдения позволили взамен представления о неспецифическом механизме восходящих активирующих влияний ретикулярной формации на кору мезга сформулировать представление о том, что подкорковые образования оказывают на кору мозга множественные восходящие активирующие влияния, каждое из которых соответствует определенной реакции того или иного биологического качества (П. К. Анохин, 1962) (2).
Такое представление позволило с новых позиций приступить к изучению корково-подкорковых механизмов «основных влечений» организма, одним из которых является состояние голода. Мы предположили, что пищевое возбуждение у голодных животных строится на основе своих специфических механизмов активации коры головного мозга со стороны подкорковых и пищевых центров.
В связи с этим мы поставили перед собой следующие основные задачи:
1. Изучить, каким образом пищевое возбуждение у голодных животных проявляется в электрической активности коры головного мозга?
2. Какие подкорковые образования определяют пищевое возбуждение животных?
3. Каковы функциональные взаимоотношения подкорковых центров с корой головного мозга у голодных животных?
4. Каким образом подкорковые образования вовлекают в свою деятельность элементы коры головного мозга, т. е. каковы те нейрофизиологические механизмы, на основе которых строится пищедобывательное поведение животных и человека?
МЕТОДИКА ОПЫТОВ
Опыты были проведены на 283 кошках под уретановым наркозом. Уретан вводили внутрибрюшинно в 7,5% или 15% растворе из расчета 1,5 гр. на кг веса животного.
В процессе наших экспериментов было установлено, что уретан, как наркотическое вещество, избирательно блокируя состояние бодрствования, в то же время оставлял относительно интактным нейрофизиологические механизмы пищевых возбуждений. Все это позволяло исследовать центральные механизмы «голодных» возбуждений в «чистом виде», не осложненные возбуждениями других биологических модальностей.
Под уретановым наркозом у животных регистрировалась ЭЭГ. Отведение ЭЭГ осуществлялось при помощи стальных игольчатых электродов, которые вкалывали непосредственно в кость черепа.
В большинстве опытов ЭЭГ отводили униполярным способом. Индифферентный электрод помещали по средней линии над лобной пазухой.
Для записи электрической активности подкорковых образований мозга так же как и для воздействия на них использовали изолированные за исключением кончика специальные погружные нихромовые электроды. Ориентацию погружения электродов в подкорковые образования проводили согласно координатам стереотаксического атласа мозга кошки, составленного Джаспером и Айжмон-Марсаном (20).
ЭЭГ записывали на чернилопишущем 10-канальном электроэнцефалографе фирмы «Альвар-Электроник».
В ряде опытов в коре мозга регистрировали вызванные потенциалы, возникающие в ответ на электрическое раздражение седалищного нерва и на раздражение подкорковых гипоталамических структур.
Регистрация вызванных ответов в коре мозга осуществлялась па 4-канальном катодно-лучевом - осциллографе «Биофаз» фирмы «Альвар-Электроник» с полосой пропускания 0,5 гц до 10 кгц.
Электрическое раздражение осуществляли стимулятором «физиовар» с трансформаторным выходом.
В специальной серии опытов производили коагуляцию или поляризацию различных подкорковых образований. Это достигалось действием на них анода постоянного тока. Локализацию кончиков электродов в мозге определяли проекционным методом (26). В ряде опытов препараты окрашивались по методу Ниселя.
РЕЗУЛЬТАТЫ ЭКСПЕРИМЕНТОВ
Электрическая активность коры головного мозга у животных в состоянии голода и после приема пищи
Для того, чтобы характеризовать, каким образом возбуждения подкорковых отделов пищевого центра выходят на «кору мозга, в первой серии экспериментов нами изучался характер электрической активности коры мозга у животных в различные сроки голодания и после приема пищи.
Опыты-показали, что у подавляющего числа животных под уретановым наркозом после 1—2-суточного голодания в передних отделах коры головного мозга регистрировалась высокочастотная низкоамплитудная электрическая активность (амплитуда колебаний 6—10 мкв, частота 20—25 кол/сек), которая резко отличалась от высокоамллитудной (50—80 мкв), относительно медленной (6—8 кол/сек) активности теменно-затылочных областей.
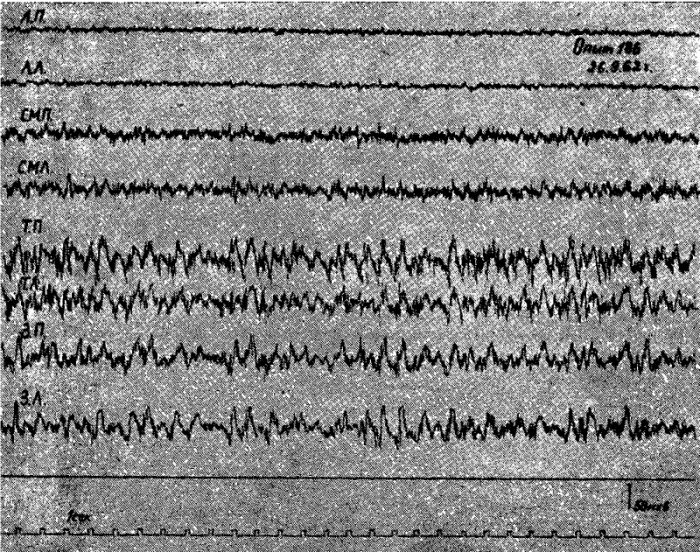
Рис. 1. Электрическая активность коры мозга кошки под уретановым наркозом после 2-суточного голодания.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сейсомоторная правая, CMЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.
Граница распространения но жоре головного мозга высокочастотной активности у животных после 1—й-суточного голодания была определена в специальных опытах. Такая активность в наиболее отчетливой форме наблюдалась над отделами мозга, расположенными впереди венечной борозды. Однако высокочастотная активность регистрировалась и в каудальных частях передних отделов мозга, в которых по мере продвижения в направлении теменных отделов постепенно нарастала высокоамплитудная активность. Задней границей распространения зоны высокочастотной низкоамплитудной ЭЭГ активности у животных после кратковременного голодания была область сильвиевых борозд.
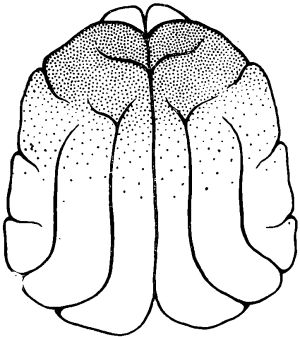
Рис. 2. Схема распространения под уретановым наркозом реакции, десинхронизации по коре мозга у кошки после 2-суточного голодания.
У накормленных перед опытом животных характер электрической активности коры мозга значительно отличался от активности голодных животных. У таких животных со всех отделах коры мозга регистрировалась медленная высокоамплитудная электрическая активность (амплитуда 50—80 мчсв, частота 3—5 кол/сек)
Все эти эксперименты указывали на то, что у животных в состоянии физиологического голода наблюдается ЭЭГ активация коры мозга. Эта активация под уретановым наркозом наиболее отчетливо проявляется в передних отделах коры мозга.
Наличие реакции десинхронизации корковой электрической активности передних отделов мозга у голодных животных указывало ка то, что эти отделы мозга находились у них в -состоянии возбуждения. При насыщении животного это возбуждение не наблюдалось. Естественно было предположить, что ЭЭГ активация передних отделов шоры мозга у голодных животных определялась «голодным» возбуждением пищевого центра.
Для выяснения того, в какой степени избирательная ЭЭГ активация передних отделов коры мозга у голодных животных была связана с деятельностью пищевого центра, мы провели специальные эксперименты.
Прежде всего мы исследовали, как скажется на ЭЭГ у голодных животных их искусственное насыщение.
Такое «насыщение» осуществлялось двояким путем: в одних случаях вводили в кровь голодного животного, находящегося под уретановым наркозом, раствор глюкозы или >кровь накормленного животного; в других опытах — животному под уретановым наркозом вводили жидкую пищу в ротовую полость, а затем в желудок.
При наличии у голодного животного реакции ЭЭГ активации только в передних отделах коры мозга и медленной высокоамллитудной активности в теменных и затылочных отделах, (введение в кровь глюкозы вызывало через 0,5—2 минуты значительное возрастание амплитуды и снижение частоты колебаний в передних отделах коры мозга. При этом иногда также увеличивалась амплитуда и снижалась частота колебаний в теменных и затылочных отделах коры мозга.
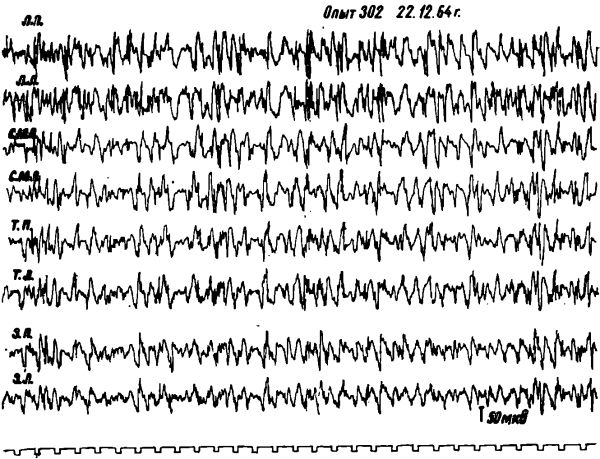
Рис. 3. Электрическая активность коры мозга кошки под уретановым наркозом после кормления.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная правая СМЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.
Pис. 4. Изменения электрической активности коры мозга у голодной кошки после введения внутривенно 40% — 2 мл глюкозы. Момент введения глюкозы обозначен стрелками. Обозначения: ЛП — лобная правая. ЛЛ — лобная левая. СМП — сензомоторная правая. СМЛ — сензомоторная левая. ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени I сек.
Следовательно, голодное состояние животного в какой-то степени влияло на деятельность и этих отделов коры.
Как правило, эффект введения глюкозы продолжался не более 15—20 минут, после чего электрическая активность коры мозга возвращалась к исходному активированному состоянию.
Аналогичные изменения ЭЭГ наблюдались при введении голодным животным крови накормленного животного.
С целью контроля в ряде опытов животным внутривенно вводили физиологический раствор. Однако введение физиологического раствора не изменяло заметно ЭЭГ голодного животного-
Произведенные опыты совершенно определенно указывали на значение гуморальных факторов для активации -коры мозга у голодных животных. Уже на основании этих опытов можно было считать, что активация передних отделов коры мозга у голодных животных определялась возбужденным «голодной» кровью пищевым центром.
Однако для более убедительного доказательства этого положения необходимо было исследовать, как изменится ЭЭГ картина голодного животного после натурального и естественного раздражения пищей.
Проведенные в этом направлении эксперименты доказали, что орошение рецепторов ротовой полости молоком, также как и введение молока в желудок, в ряде опытов приводило к временному устранению активации ЭЭГ у голодных животных.
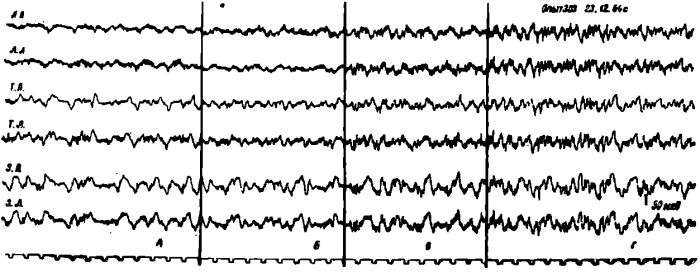
Рис. 5. Изменения электрической активности коры мозга у голодной кошки после введения ей молока в желудок. А — исходная ЭЭГ голодного состояния. Б — через 5 мин. В — через 15 мин. Г — через 40 мин. после введения молока в желудок. Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная правая. СМЛ - сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.
Только через час после введения пищи в желудок наблюдалось устойчивое возрастание амплитуды биотоков и устранение «голодной» активации коры мозга.
Все эти опыты указывали на то, что наблюдаемая у голодных животных под уретановым наркозом избирательная регионарная активация передних отделов коры мозга обусловливалась восходящими активирующими влияниями пищевого центра.
Нейрогуморальные механизмы избирательной активации коры мозга при пищевом возбуждении
Дальнейшие наши эксперименты показали, что активация, коры мозга у голодных животных устранялась не только после введения им в кровь раствора глюкозы, но и после одновременной перерезки обоих блуждающих нервов на шее и спинного мозга на уровне грудных или шейных сегментов.
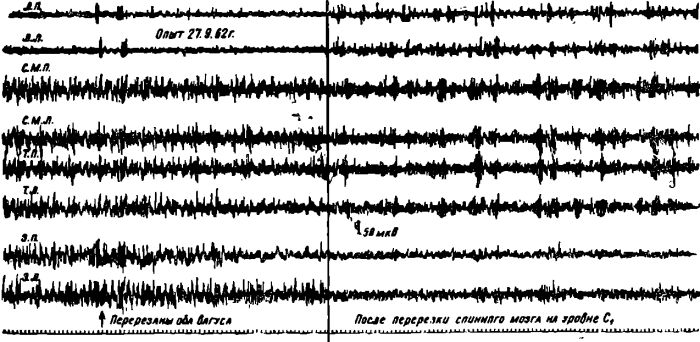
Рис. 6. Изменение электрической активности коры мозга у голодной кошки после перерезки у нее обоих блуждающих нервов и спинного мозга на уровне первого шейного сегмента. В последнем случае животное переведено на искусственное дыхание.
Известно, что основная нервная афферентация, идущая в центральную нервную систему от желудка, поступает именно по этим путям. Изолированная перерезка только блуждающих нервов или спинного мозга не устраняла «голодной» активации коры мозга.
Таким образом, кора головного мозга у голодных животных и особенно ее передние отделы находятся под одновременным двойным влиянием «голодной» крови и афферентной сигнализации, поступающей от пустого желудка как по блуждающим нервам, так и по спинному мозгу.
Важно подчеркнуть, что все эти влияния оказывают в конечном счете одинаковое действие на корковые нейроны, создавая избирательное состояние ЭЭГ активации в передних отделах коры, не блокируемое наркотическим веществом.
Проведенные эксперименты позволили проанализировать некоторые стороны пищевого насыщения и прежде всего разрешить вопрос о том, каким образом при этом устраняется «голодная» активация коры мозга.
Мы установили, что устранение «голодной» активации коры мозга происходит уже при орошении рецепторов ротовой полости. Оно может усиливаться при последующем введении пищи в желудок. Однако наиболее устойчивое устранение ЭЭГ активации коры наблюдается только после поступления в кровь питательных веществ (через час после кормления).
Аналогичным образом эффект действия глюкозы в наших опытах был всегда более отчетливым -и продолжительным, если глюкозу вводили животному после введения пищи в желудок.
Проведенные опыты указывали, таким образом, на то, что изменения пищевого центра, характеризующие состояние насыщения животных, определяются также двойными влияниями нервных и гуморальных факторов- Причем наиболее отчетливое и устойчивое действие гуморальных раздражителей на центральные аппараты пищевого центра осуществляются только после воздействия пищи на рецепторы ротовой полости и желудка.
Тот факт, что после приема пищи исчезает избирательная ЭЭГ активация передних отделов коры мозга, которая отчетливо наблюдается в голодном состоянии, указывает на то, что при насыщении значительно снижаются восходящие активирующие влияния со стороны подкорковых образований пищевого центра на кору головного мозга.
Важно отметить то обстоятельство, что каждый этап прохождения пищи по пищеварительному каналу вызывает путем нервной сигнализации все углубляющиеся однотипные изменения в деятельности пищевого центра — снижение его функциональной активности.
Снижение функциональной активности, которое возникает в пищевом центре при попадании пищи в ротовую полость, подкрепляется в дальнейшее афферентными влияниями, идущими из желудка. Однако эти изменения являются кратковременными. Только после всасывания пищевых веществ в кровь последние вызывают устойчивое снижение возбудимости пищевого центра.
Таким образом, для успешного осуществления процесса насыщения основным условием является естественная последовательность пищеварительных процессов. Проведенные опыты показали, что любое искусственное нарушение последовательности этого процесса, например, введение пищи сразу в желудок или глюкозы в кровь на фоне пустого желудка вызывает только кратковременные изменения ЭЭГ, характерные для сытого состояния.
Об участии подкорковых аппаратов в механизме избирательной активации коры мозга у голодных животных
В связи с наличием у голодных животных избирательной ЭЭГ активации коры мозга, особенно ее передних отделов, перед нами возник вопрос, какие подкорковые образования ее определяют?
Поскольку в настоящее время показано, -что основные центры, определяющие пищевые мотивации животных, расположены в гипоталамической области (17, 19, 21, 23 и др.) мы, естественно, прежде всего решили исследовать степень участия латеральных («центр питания») и медиальных («центр насыщения») отделов гипоталамуса в механизме обнаруженной нами «голодной» активации коры мозга.
Эти исследования были также проведены на кошках, находящихся под уретановым наркозом. Мы исходили из следующих соображений. Если уретан, избирательно блокируя бодрствующее состояние, допускает проявление пищевого возбуждения в деятельности коры мозга, то при этом должны сохранить свою функциональную активность все элементы пищевого центра, на каком бы уровне центральной нервной системы они не находились.
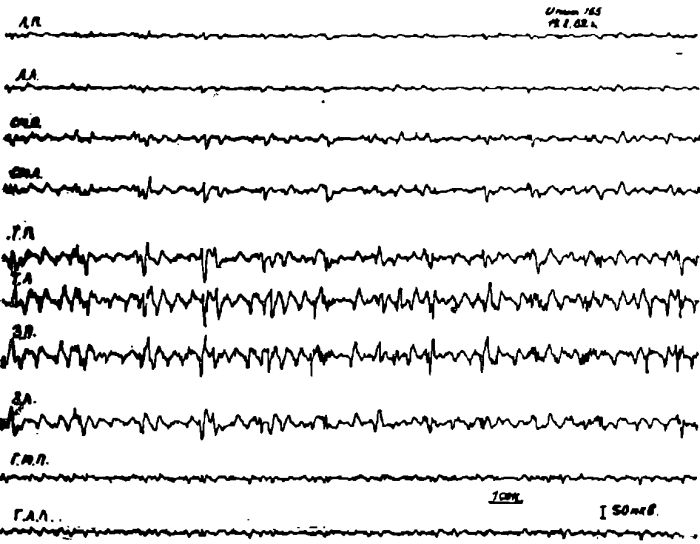
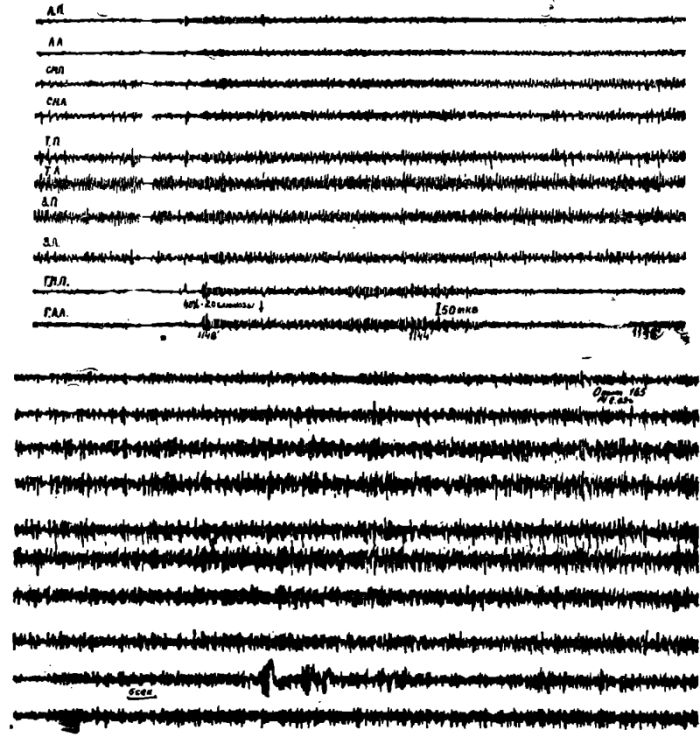
Рис. 7. Электрическая активность коры мозга и пищевых центров латерального и медиального гипоталамуса у кошки после 2-суточного голодания.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная правая, СМЛ — сензомоторная левая, ТГ1 — теменная правая, ТЛ — теменная левая. ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек. ГЛЛ — левая латеральная гипоталамическая область; ГМП — правое вентромеднальное ядро гипоталамуса.
Проведенные опыты показали, что у животных под уретановым наркозом после 1—2 суточного голодания характер электрической активности латеральных и медиальных отделов гипоталамуса почти точно соответствовал электрической активности передних отделов коры мозга.
В подавляющем большинстве экспериментов в гипоталамических отделах пищевого центра регистрировалась такая же как и в передних отделах коры мозга высокочастотная низкоамплитудная электрическая активность (амплитуда 2—5 мкв, частота 30—40 кол/сек).
У животных, которые были накормлены до введения наркотического вещества, в этих отделах гипоталамуса регистрировались медленные высокоамплитудные колебания (амплитуда 50—70 мкв, частота 6—8 кол/сек).
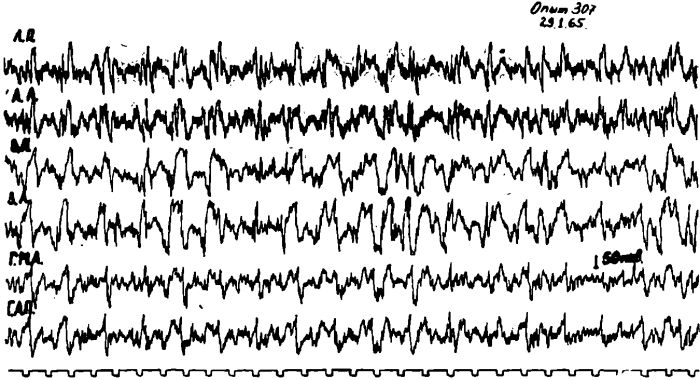
Рис. 8. Электрическая активность коры мозга и пищевых центров гипоталамуса у накормленной кошки, находящейся под уретановым наркозом.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная правая, СМЛ — сензомоторная левая, ГП — теменная правая, ТЛ — теменная левая ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек. ГМЛ — левое всптромеднальнос ядро гипоталамуса; ГЛП — працан латеральная гипоталамнческая область.
Введение глюкозы голодным животным вызывало наряду с появлением медленной высокоамплитудной электрической активности в передних отделах коры возникновение такой же активности в медиальных и латеральных отделах гипоталамуса.
То же наблюдалось при раздражении пищей рецепторов ротовой полости и через 50—70 минут после (введения голодным животным пищи в желудок. Наличие у голодных животных в латеральном и медиальном гипоталамусе, так же как и в передних отделах торы мозга, низкоамплитудной высокочастотной электрической активности уже само по себе указывает на то, что пищевые центры гипоталамуса, так же как и клетки передних отделов коры мозга, находятся у них в состоянии возбуждения. После приема пищи это возбуждение исчезает. В связи с этим перед нами возник вопрос, не являются ли вышеупомянутые центры гипоталамуса тем инициативным пунктом, который в состоянии голода и определяет восходящие влияния на кору мозга. Для выяснения этого вопроса в последующих опытах был применен метод локального воздействия постоянным током (на различные отделы гипоталамуса.
Известно, что локальное действие анодом постоянного тока на любую возбудимую систему вызывает снижение ее возбудимости, т. е. так называемый анэлектротон. С другой стороны, действие катода вызывает повышение возбудимости — катэлектротон. Используя методику воздействия постоянным током, мы рассчитывали и а то, что если восходящие активирующие влияния на передние отделы коры мозга при голоде определяются гипоталамическими отделами пищевого центра, то искусственное снижение их возбудимости при действии анода постоянного тока должно привести к устранению этой активации. Наоборот, активация коры мозга должна, естественно, усилиться при действии на них катода постоянного тока. Результаты опытов полностью подтвердили наши предположения. Двухстороннее локальное воздействие анодом постоянного тока на латеральные и медиальные отделы гипоталамуса вызывало у голодных животных исчезновение ЭЭГ активации передних отделов коры головного мозга и значительное возрастание амплитуды потенциалов во всех корковых отведениях. После этого электрическая активность во всех отделах коры мозга становилась характерной для состояния глубокого сна.
Подобный эффект вызывала и коагуляция вышеуказанных областей гипоталамуса.
Наоборот, действие катода и а эти отделы гипоталамуса приводило к возникновению генерализованной ЭЭГ активации коры мозга.
Все эти опыты указывали на то, что активация передних отделов коры мозга у голодных животных определяется высотой функциональной активностью гипоталамических отделов пищевого центра. При снижении возбудимости этих отделов гипоталамуса при помощи анэйектротона «голодная» активация коры мозга исчезает.
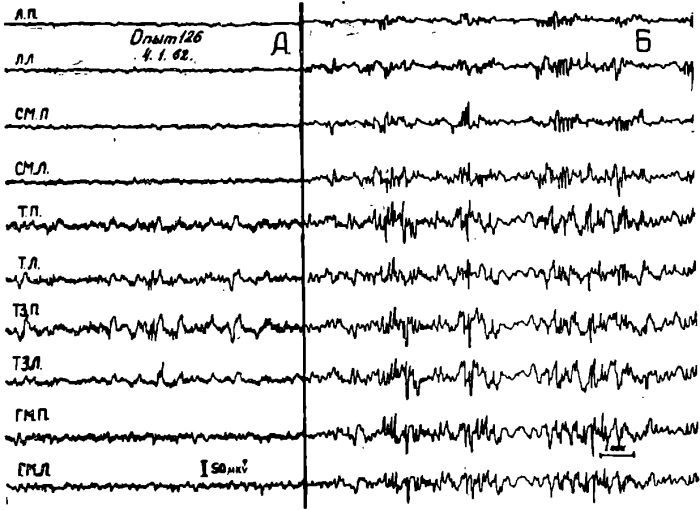
Рис. 9. Изменения характера электрической активности коры мозга голодной кошки (левая часть рисунка) после действия анода постоянного тока (0,5мА — 0,5 мии) на вентромедиальные ядра гипоталамуса.
ГЛЛ — левая латеральная гипоталамическая область; ГМП — правое вентромедиальное ядро гипоталамуса. ГМЛ — левое вентромедиальное ядро гипоталамуса; ГЛП — правая латеральная гипоталамическая область.
Все это позволяет сделать вывод о том, что в состоянии голода избирательная ЭЭГ активация передних отделов коры мозга в значительной степени определяется восходящими активирующими влияниями гипоталамуса. Насыщение, наоборот, связано со снижением функциональной активности последнего, благодаря чему исчезает «голодная» активация коры мозга.
Дальнейшие опыты показали, что у голодных животных под уретановым наркозом наряду с возбуждением передних отделов коры мозга и пищевых центров гипоталамуса наблюдается возбуждение в ретикулярной формации среднего мозга и в медиальных отделах таламуса.
Возбуждение в этих структурах также исчезает после искусственного насыщения животных, т. е. после введения им в кровь глюкозы или жидкой пищи в ротовую полость и желудок.
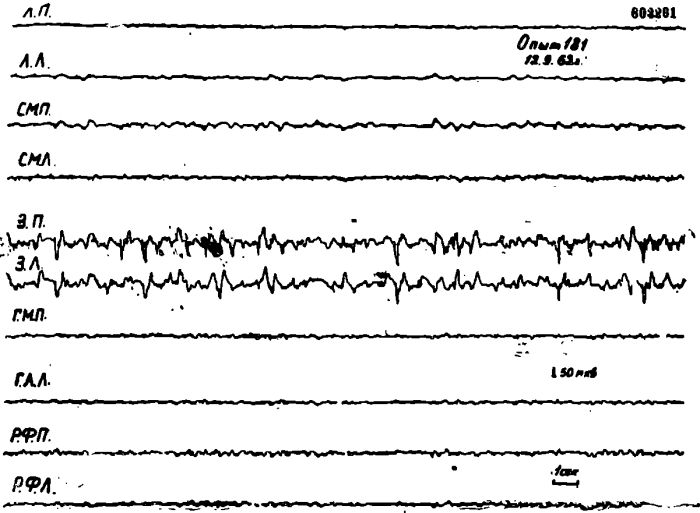
Рис. 10. Электрическая активность коры мозга правого медиального та-ламуса (ТМП), левой латеральной гипоталамической области (ГЛЛ) и области, расположенной справа и слева вокруг Сильвиева водопровода на уровне верхних бугорков четверохолмия (РФП и РФЛ) у кошки под уретановым наркозом после 2-суточного голодания.
Это указывало на то, что у голодных животных имеется целая система возбужденных нервных элементов, объединяющая как различные подкорковые образования, так и нервные клетки коры головного мозга. Примечательно, что вся эта система возбужденных нервных элементов те блокировалась уретаном. В связи с этим перед нами возник вопрос, каковы функциональные взаимоотношения между подкорковыми аппаратами этой системы, какие из них являются ведущими и определяющими возбуждение всей системы в целом. Ответить на этот вопрос позволили следующие эксперименты.
Было установлено, что действие анода постоянного тока на пищевые центры гипоталамуса, как правило, приводило к устранению «голодного» возбуждения в ретикулярной формации среднего мозга и медиальных отделах таламуса, так же как и в коре головного мозга. Такое же воздействие анода постоянного тока на медиальные отделы таламуса приводило к устранению ЭЭГ активации только в коре мозга. В гипоталамических отделах и в ретикулярной формации при этом сохранялось состояние возбуждения. При действии анода постоянного тока на ретикулярную формацию среднего мозга (на уровне верхних бугорков четверохолмия) ЭЭГ активация устранялась только в теменных и затылочных отделах коры мозга (если она там до этого регистрировалась). В передних отделах коры, в медиальных ядрах таламуса и в пищевых центрах гипоталамуса сохранялось состояние возбуждения. Эти опыты указывают на то, что в системе пищевого возбуждения у голодных животных центральная и инициативная роль принадлежит пищевым центрам гипоталамуса. Наряду с восходящими активирующими влияниями на кору мозга эти отделы гипоталамуса оказывают у голодных животных активирующее влияние и на ретикулярную формацию среднего мозга -и медиальные отделы таламуса. Распространение восходящих активирующих влияний гипоталамуса на кору мозга осуществляется преимущественно через медиальную группу ядер таламуса.
Проведенные эксперименты поставили перед нами следующий вопрос. Если гипоталамичеокие отделы пищевого центра являются инициативными в механизме восходящих активирующих влияний на кору мозга, то каким образом эти влияния распространяются на корковые синаптические организации?
Вопрос этот чрезвычайно важен, поскольку только расшифровка механизма избирательного вовлечения синаптических образований коры в пищевое возбуждение, вызванное состоянием голода, позволит прямо установить, на основе каких механизмов формируется целенаправленное пищевое поведение?
Распространение восходящих активирующих влияний гипоталамуса на синаптические организации коры мозга у голодных животных
Приступая к настоящей серии исследований, мы решили в качестве показателя функциональной активности синаптических организаций коры мозга использовать вызванный потенциал, возникающий в коре мозга в ответ на одиночное раздражение седалищного нерва.
Предыдущими исследованиями сотрудников нашей лаборатории (3, 4, 10, 13 и др.) было показано, что вызванный потенциал, регистрируемый с поверхности коры мозга, представляет собой результат множественных восходящих влияний, поступающих к синапсам корковых клеток по различным каналам от различных подкорковых образований.
Согласно этим представлениям, отрицательная фаза и вторичные ответы вызванного потенциала обусловливаются в коре множественными восходящими влияниями неспецифических подкорковых систем, которые адресуются к аксодендритным синапсам поверхностного плексиморфного слоя. В то же время положительная фаза первичного ответа определяется возбуждениями аксосоматических сипапсов на нейронах IV слоя коры за счет прихода возбуждений по классическим лемнисковым путям (2, 3 и др.).
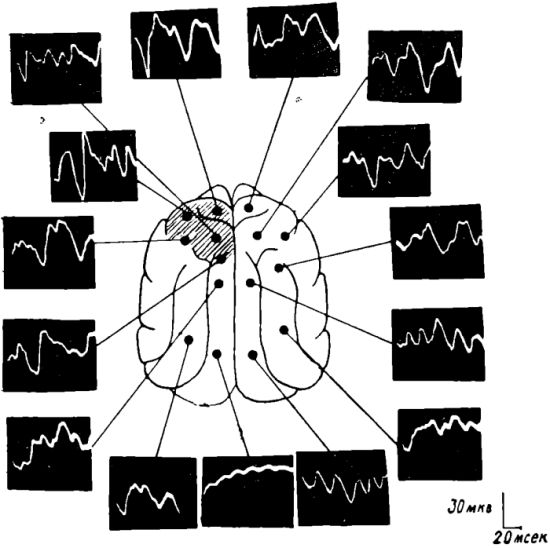
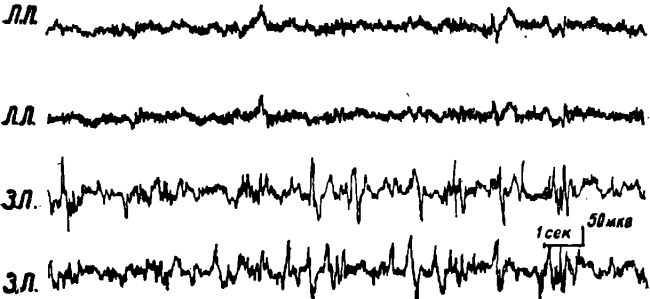
Рис. 11. ЭЭГ (внизу) и карта, показывающая отсутствие распространения соматосензорных вызванных потенциалов на кору мозга у кошки после 2-суточного голодания. Уретановый наркоз.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная правая, СМЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗГТ — затылочная правая, ЗЛ — затылочная левая обл.
Отметка времени 1 сек.
Приступая к настоящей серии экспериментом, мы исходили из следующего предположения. Состояние голода на основе восходящих активирующих влияний подкорковых образований, создает в центральной нервной системе определенное доминантное состояние, сопровождающееся мобилизацией определенных корковых синаптических образований. В этом состоянии любое возбуждение другой биологической модальности, поступающее к коре мозга, должно было неизбежно вступить в какие-то взаимодействия с наличным пищевым возбуждением и, естественно, претерпеть определенные изменения. Особенно отчетливо эти изменения должны быть выражены у соматосензорного вызванного ответа, поскольку формирующее его возбуждение адресуется к тем же передним отделам -коры мозга, куда поступают и возбуждения пищевого центра.
Проведенные опыты показали следующее. У голодных животных, находящихся под уретановым наркозом, соматосензорные вызванные потенциалы в ответ на одиночное раздражение седалищного нерва регистрировались только в ограниченном участке сензомоторной области коры мозга, т. е. в той области, которая является специфической проекционной зоной соматосензорных возбуждений.
Как правило, в этом пункте регистрировались вызванные ответы только с первичноположительной фазой. В ряде опытов у голодных животных соматосензорные вызванные ответы вообще не регистрировались. Характерно, что в то же время и в том же пункте коры вызванные потенциалы, возникающие у голодных животных при раздражении пищевых центров гипоталамуса, выявлялись в коре мозга совершенно отчетливо.
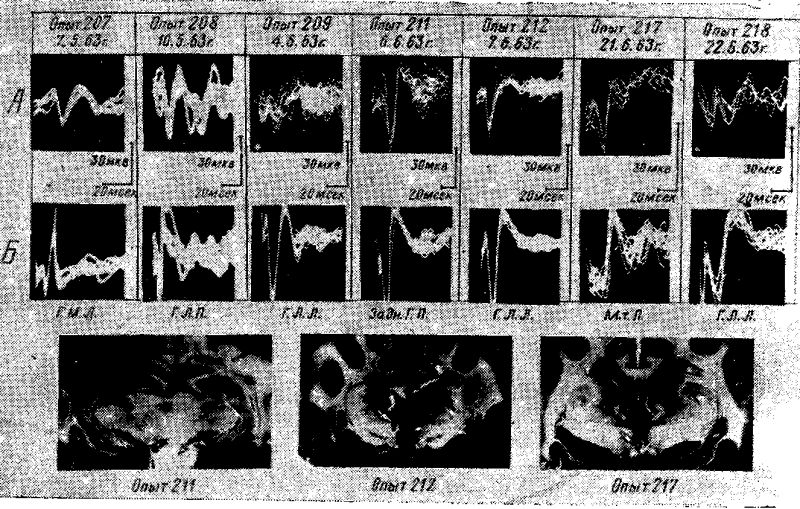
Рис. 12. Вызванные потенциалы в сензомоторной области коры у голодных кошек, находящихся под уретановым наркозом, при раздражении седалищного нерва (А) и различных отделов гипоталамуса 1Б.

Pиc. 13. ЭЭГ (внизу) и карта распространения соматосензорного вызванного потенциала по коре мозга кошки, накормленной перед опытом. Уретановый наркоз.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая. СМП — сензомоторная правая. СМЛ — сензомоторная левая, ТП — теменная правая, TЛ — теменная левая, ЗП — затылочная правая, 3Л — затылочная левая обл. Отметка времени 1 сек.
Наоборот, у накормленных перед опытом животных соматосензорные вызванные ответы с отчетливо выраженными первичной положительной и отрицательной фазами и нередко с вторичным положительным колебанием регистрировались над обширной областью передних отделов коры мозга.
Аналогичный эффект возрастании амплитуды отрицательной фазы в фокусе максимальной активности и расширения зоны регистрации вызванных ответов наблюдался при введении голодным животным раствора глюкозы или при -коагуляции медиальных отделов гипоталамической области.
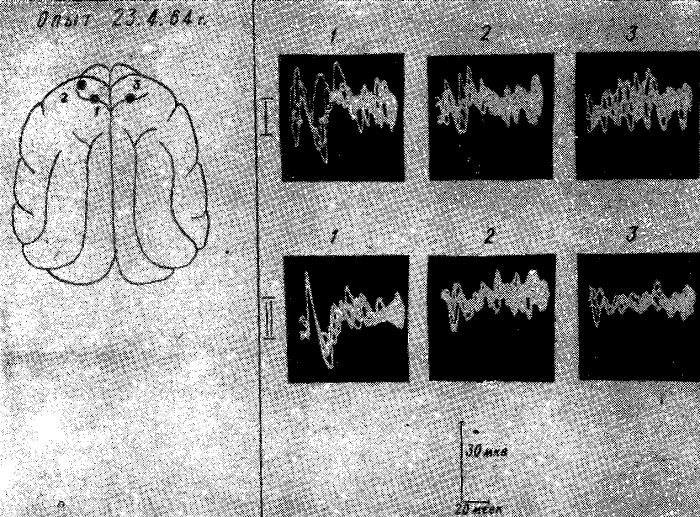
Рис. 14. Изменения характера выявления и распространения соматосензорного вызванного ответа по коре мозга у голодной кошки после внутривенного введения 40% — 2 мл раствора глюкозы. Уретановый наркоз.
I — вызванный ответ в тех же точках 1, 2, 3 до введения глюкозы; II — вызванный ответ в тех же точках через 15 минут после введения глюкозы.
Все это указывало на то, что изменения соматосензорного вызванного ответа у голодного животного определялись восходящими активирующими влияниями гипоталамуса на кору мозга.
В результате этих пищевых возбуждений происходит своеобразное ««вычитание» синапсов из целостной деятельности, участвующей в формировании соматосензорных вызванных потенциалов.
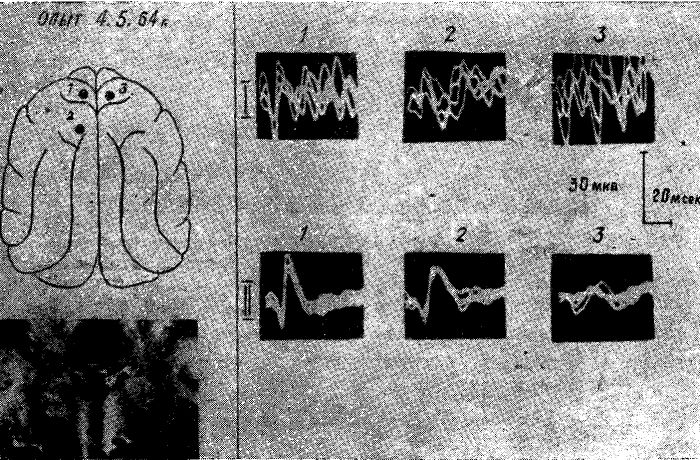
Рис. 15. Изменение характера и распространения соматосензорного вызванного ответа по коре мозга у голодной кошки после коагуляции вентромедиальных отделов гипоталамуса и субталамуса. Уретановый наркоз.
I — исходный вызванный потенциал в точках 1, 2, 3. II — вызванный потенциал в тех же точках после коагуляции медиальных отделов гипоталамуса и субталамуса.
Поскольку в состоянии голода у животных наблюдалось избирательное подавление отрицательной фазы первичного ответа и вторичного положительного колебания соматосензорного вызванного ответа, то можно думать, что возбуждения гипоталамических отделов пищевого центра в состоянии голода адресуются именно к тем синаптическим организациям сензомоторной области коры, которые формируют эти фазы вызванного потенциала. Так как отрицательная фаза первичного ответа, а также вторичные потенциалы обусловливаются восходящими влияниями неспецифических подкорковых систем, адресующихся к аксодендритньим синапсам поверхностных слоев коры (2, 3, 4 и др.), можно думать, что восходящие активирующие влияния гипоталамических отделов пищевого центра специфически адресуются именно к этим синаптическим образованиям коры головного мозга.
Специальные опыты показали, что блокада распространения самотосензорных потенциалов по коре мозга у голодных животных происходит и на уровне медиальных ядер таламуса. Коагуляция медиальных ядер таламуса у накормленных животных прекращает генерализованное распространение вызванных потенциалов на передние отделы контралатерального полушария.
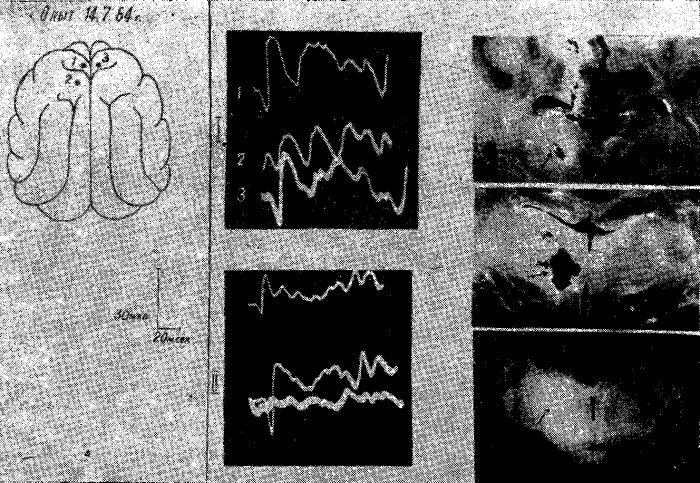
Рис. 16. Изменения распространения соматосензорного вызванного потенциала по коре мозга у накормленной кошки. Уретановый наркоз.
I — до, II — после коагуляции медиальных групп ядер правого таламуса. После этого вызванный потенциал перестает распространяться на ипсилатеральной стороне.
На основании этих опытов мы пришли к выводу, что восходящие активирующие влияния гипоталамуса в состоянии голода специфически распространяются и на аксодендритные синапсы передних отделов коры за счет первичного возбуждения медиальной группы таламических ядер.
Нейрохимические механизмы восходящих активирующих влияний на кору мозга у голодных животных
Наличие у голодных животных системы пищевого возбуждения избирательно объединяющей корковые и подкорковые элементы, поставило перед нами вопрос, какие механизмы производят функциональное объединение различных нервных элементов в единую систему пищевого возбуждения, не блокируемую даже уретаном? Мы предположили, что такое функциональное объединение при пищевом возбуждении может быть основано на едином химическом механизме системы восходящих активирующих влияний гипоталамических пищевых центров на кору головного мозга.
Эксперименты, проведенные в нашей лаборатории показали, что такое адренологическое вещество, как аминазин, избирательно блокирует механизмы восходящей активности коры при болевой реакции, но оставляет при этом относительно интактной активацию коры мозга, обусловленную «голодным» возбуждением животного (15). Это позволяло думать, что пищевое возбуждение у голодных животных определялось не адренергическими механизмами мозга. Учитывая литературные данные о том, что ретикулярная формация ствола мозга использует в своих восходящих активирующих влияниях на кору мозга и холинергические механизмы (7, 8, 9, 18, 28 и др.) на предположили, что механизмы восходящих активирующих влияний на кору мозга у голодных животных строятся на основе холинореактивных элементов. Опыты подтвердили такое предположение.
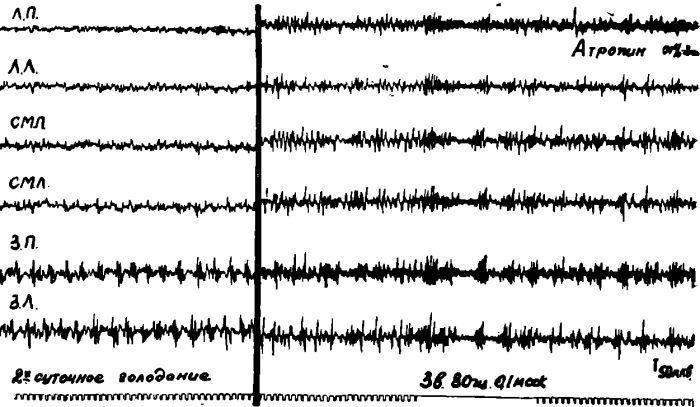
Рис. 17. Изменения ЭЭГ голодной кошки после подкожного введения 0,1%—2 мл атропина (правая часть рисунка). В правой части рисунка кроме того показан эффект раздражения седалищного нерва на фоне действия атропина. Уретановый наркоз.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сензомоторная: правая. СМЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.
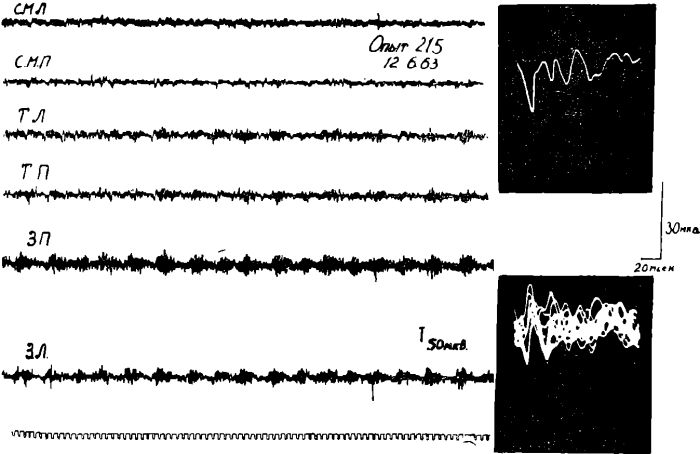
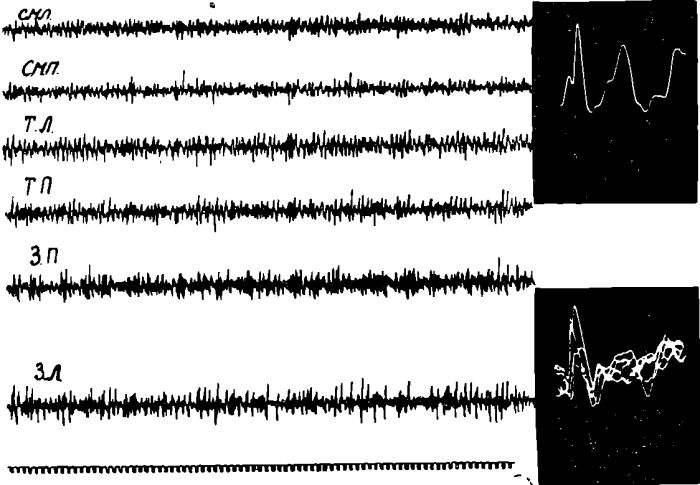
Рис. 18. Изменения характера соматосензорного вызванного потенциала, регистрируемого в сензомоторной коре I — до, II — после введения 0,1% — 2 мл атропина. Уретановый наркоз.
Так, подкожное введение голодным животным растворов атропина или амизила из расчета 0,5—0,6 мг на 1 кг веса на фоне отчетливой избирательной десинхронизации ЭЭГ в передних отделах коры мозга уже через 2—5 минут устраняло «голодное» возбуждение и приводило к появлению во всей коре мозга медленных высокоамплитудных колебаний.
Однако, пороговое раздражение седалищного нерва при этом по-прежнему вызывало, хотя и менее отчетливую, генерализованную ЭЭГ активацию всей коры мозга. После введения голодным животным холинолитических веществ у них, так же как и у накормленных животных, начинала отчетливо регистрироваться отрицательная фаза корковых соматосензорных потенциалов. Это указывало на то, что холинолитические вещества избирательно блокировали только механизмы пищевой восходящей активации коры больших полушарий, оставляя относительно интактными механизмы болевой активации. Дальнейшее подтверждение этого было получено в опытах, проведенных в нашей лаборатории (14).
Аппликация холинолитических веществ непосредственно на кору мозга у голодных животных в его опытах приводила в пункте аппликации к устранению «голодной» активации и резкому возрастанию здесь отрицательной фазы соматосензорных вызванных потенциалов. Аналогичные данные были получены и А. А. Панфиловым (1965), «который показал, что избирательная ЭЭГ активация передних отделов коры мозга у голодных животных исчезает после локальной инъекции холинолитимов непосредственно в область латерального гипоталамуса.
Все это указывало на то, что система пищевого возбуждения у животных после 1—2-суточного голодания строится преимущественно на основе холинергических механизмов.
Однако при этом следует иметь в виду, что в активации коры мозга у «голодных животных принимают участие и адренергические механизмы. Опыты показали, что адренергические механизмы вое более интенсивно возбуждаются при увеличении сроков голодания животных Наблюдаемая у животных после 4-суточного голодания генерализованная ЭЭГ активация, в отличие от региональной активации передних -отделов коры мозга у животных после 1—2-суточного голодания, блокируются аминазином. Это дает основание считать, что адренергические механизмы определяют ту реакцию общего напряжения организма, которая развивается при увеличении сроков голодания.
Заключение
Проведенные нами эксперименты показывают, что пищевое возбуждение у голодных животных охватывает целую систему функционально объединенных нервных элементов, находящихся как в подкорковых образованиях, так и в кюре головного мозга. Эта система «голодного» возбуждения формируется на основе специфических механизмов восходящих активирующих влияний подкорковых образований на кору головного мозга. Возбуждения подкорковых образований строго избирательно мобилизуют элементы коры головоного мозга, что выражается в специфической форме активации ЭЭГ голодных животных.
В механизме этих восходящих активирующих влияний на кору мозга центральная роль принадлежит пищевым центрам гипоталамичеовой области. Искусственное снижение функциональной активности пищевых центров гипоталамуса приводит к устранению пищевого возбуждения у голодных животных, как в подкорковых образованиях, так и в коре головного мозга.
Из всего этого следует, что гипоталамические отделы пищевого центра обладают огромной силой тонического воздействия на другие структуры мозга и прежде всего на кору больших полушарий. Все это дает нам основание рассматривать пищевые центры гипоталамуса как своеобразный «пейцмекер» всей системы пищевого возбуждения у голодных животных.
Проведенные опыты показывают, что распространение восходящих активирующих влияний гипоталамуса на кору мозга у голодных животных осуществляется через медиальные ядра таламуса. Наряду с этим гипоталамические пищевые центры оказывают активирующее действие на ретикулярную фармацию среднего мозга. Эти влияния все более усиливаются при увеличении сроков голодания. Вовлечение в систему пищевого возбуждения ретикулярной формации среднею мозга приводит к появлению генерализованных активирующих восходящих влияний на всю кору головного мозга, что, в свою очередь, определяет еще более высокий энергетический тонус пищедобывательного поведения.
Разобранные нами специальные механизмы восходящих активирующих влияний на кору мозга у голодных животных подтверждают представления нашей лаборатории о такой функциональной организации взаимоотношений коры и подкорки, при которой подкорковые образования берут на себя роль своеобразного источника энергии, который избирательно поддерживает в рабочем тонусе всю обширную функциональную систему данной специфичности. Очевидно, именно, через этот физиологический механизм реализует себя та подкорковая «слепая сила», о которой много раз писал И. П. Павлов.
Наши опыты показали, что восходящие активирующие влияния подкорковых областей на кору мозга при голоде за счет первичного возбуждения медиальных ядер таламуса распространяется преимущественно на аксодендритные синапсы плексиморфного слоя коры мозга. Восходящие активирующие влияния подкорковых отделав пищевого центра у голодного животного производят избирательную мобилизацию корковых синапсов из их общей массы. Только на основе избирательного возбуждения клеток коры мозга может возникнуть специальная пищедобывательная деятельность. Такое избирательное функциональное вовлечение инициативными подкорковыми пищевыми центрами в свою деятельность элементов коры мозга у голодных животных происходит на основе их единых, преимущественно холинергичеcких, механизмов.
Из всего изложенного выше следует, что пищевое возбуждение формируется на основе такой вертикальной построенной функциональной организации, в которой инициативным пунктом являются пищевые центры гипоталамуса, избирательно мобилизующие для пищедобывательной деятельности другие подкорковые аппараты, а также клетки коры головного мозга.
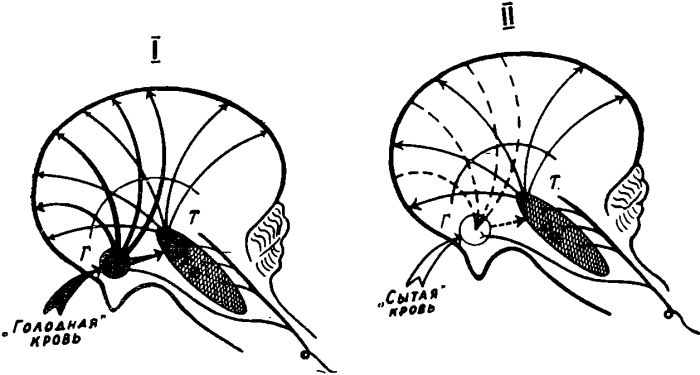
Рис. 19. Схема восходящих активирующих влиянии гипоталамических отделов пищевого центра на кору мозга. I — состояние голода, II — после приема пищи. Обозначения: Г — гипоталамус, Т — таламус, РФ — ретикулярная формация.
Благодаря таким вертикально организованным системам восходящих активирующих влияний подкорковых образований на кору мозга, весьма малое количество начальной энергии, которое образуется при возбуждении центральных рецепторов через мобилизацию многочисленных корковых нейронов, может привести в последующем к появлению огромной энергии. Именно в этом заключается смысл формирования этих реакций по доминантному типу.
Выводы
1. Нейрофизиологическую основу состояния голода и пищевой мотивации, определяющей пищедобывательное поведение, составляют специфические механизмы восходящих активирующих влияний подкорковых отделов пищевого центра на кору головного мозга.
Пищевое возбуждение у животных после 1—2-суточного голодания проявляется в форме избирательной активации ЭЭГ передних отделов коры мозга даже под уретановым наркозом, несмотря на глубокий сон животного.
2. Избирательная активация коры головного мозга у голодных животных определяется двойными влияниями со стороны «голодной» крови и нервных импульсаций, поступающих от желудочно-кишечного тракта по блуждающим нервам и чувствительным волокнам, идущим через спинной мозг.
3. Активация коры головного мозга у голодных животных определяется целой системой возбужденных подкорковых образований, находящихся на различных уровнях ствола мозга. Она объединяет пищевые центры гипоталамуса (латеральные и медиальные его отделы), медиальные отделы таламуса и ретикулярной формации среднего мозга. Во всех этих отделах у голодных животных под уретановым наркозом, так же как и в передних отделах коры мозга, регистрируются высокочастотная низкоамплитудная электрическая активность, характеризующая состояние возбуждения.
4. Центральным и инициативным пунктом системы возбужденных подкорковых образований, оказывающих у голодных животных восходящие активирующие влияния на кору мозга, являются гипоталамические отделы пищевого Центра. Их можно рассматривать как своеобразный «пейцмекер» всей системы пищевого возбуждения.
5. Восходящие активирующие влияния гипоталамуса на кору мозга у голодных животных распространяются преимущественно через медиальные отделы таламуса.
Наряду с восходящими активирующими влияниями на кору мозга гипоталамические отделы пищевого центра у -голодных животных оказывают активирующие влияния и на ретикулярную формацию среднего мозга. Эти влияния гипоталамических центров на ретикулярную формацию среднего мозга значительно усиливаются при увеличении сроков голодания.
6. Восходящие активирующие влияния -гипоталамуса на кору мозга в состоянии голода через первичное возбуждение медиальных ядер таламуса распространяются преимущественно на аксодендритные синапсы поверхностного плексиморфного слоя передних отделов коры мозга.
7. Функциональное объединение различных нервных элементов, находящихся в подкорковых образованиях и в коре мозга, в систему пищевого возбуждения у животных после 1—2-суточного голодания происходит на основе единых химических, преимущественно холинергических механизмов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
§ 28. Возникновение отделов головного мозга
§ 28. Возникновение отделов головного мозга Ранний период истории возникновения предков позвоночных, до формирования хорошо структурированного скелета, довольно туманен. Если допустить, что предковые формы хордовых были мягкотелыми существами размером около 10–15 см,
Увеличение головного мозга
Увеличение головного мозга Итак, чтобы объяснить, откуда возникло название этой главы, следует прежде всего подчеркнуть, что мы понимаем под поведением, и затем показать особую связь поведения с мозгом. Термин «поведение» имеет самые разнообразные значения; мы же будем
Часть I ФИЗИОЛОГИЯ ГОЛОВНОГО МОЗГА ЧЕЛОВЕКА
Часть I ФИЗИОЛОГИЯ ГОЛОВНОГО МОЗГА ЧЕЛОВЕКА Глава 1. Развитие нервной системы человекаГлава 2. Клетка – основная единица нервной тканиГлава 3. Активирующие системы мозгаГлава 4. Физиологические механизмы регуляции вегетативных функций и инстинктивного
ЛИМБИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА
ЛИМБИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА Лимбическая система в мозге человека выполняет очень важную функцию, которая называется мотивационно-эмоциональной. Чтобы было ясно, что это за функция, вспомним: каждый организм, включая организм человека, имеет целый набор
Травмы головного и спинного мозга
Травмы головного и спинного мозга Несомненно, что всех животных с травматическим повреждением любого вида, в том числе и с травмой головного и спинного мозга, прежде всего должен осмотреть ветеринар — это необходимо для уточнения диагноза и определения показаний для
Болезни головного мозга
Болезни головного мозга При заболеваниях головного мозга развиваются следующие расстройства: судороги, параличи, которые возникают в результате поражения двигательной области коры головного мозга или двигательных путей от коры в боковой столб противоположной
ВЫХОДИТ ЛИ РАЗУМ ЗА ПРЕДЕЛЫ ГОЛОВНОГО МОЗГА?
ВЫХОДИТ ЛИ РАЗУМ ЗА ПРЕДЕЛЫ ГОЛОВНОГО МОЗГА? Когда мы смотрим на предметы, где мы их видим в действительности? Находятся ли образы внутри мозга или снаружи — именно там, где мы их видим? Общепринятое научное разъяснение гласит, что эти образы находятся внутри головного
4.1.1. Органическое поражение головного мозга
4.1.1. Органическое поражение головного мозга Животное пытается достать лакомство, но быстро прекращает попытки (истощаемость), либо не пытается достать лакомство (депрессия, апатия), либо постоянно отвлекается на другие внешние раздражители, вплоть до переключения на
Травмы головного мозга
Травмы головного мозга Встречаются относительно редко.¦ ЭТИОЛОГИЯ И ПАТОГЕНЕЗТравмы головного мозга регистрируются после ударов или падения с высоты. Сопровождаются сотрясением мозга и кровоизлияниями различной степени.¦ СИМПТОМЫПосле удара или падения собака
Анализ физиологических механизмов восходящих активирующих влияний на кору мозга у голодных животных методом аппликации различных фармакологических веществ А. И. ТУРЕНКО (Москва)
Анализ физиологических механизмов восходящих активирующих влияний на кору мозга у голодных животных методом аппликации различных фармакологических веществ А. И. ТУРЕНКО (Москва) Многочисленные исследования показали, что основным регулятором потребления пищи является
Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва)
Активность нейронов коры головного мозга у голодных животных Ю. А. ФАДЕЕВ (Москва) Успехи, современной нейрофизиологии, связанные с использованием новых тонких методов электрофизиологического исследования мозга, позволили подойти к изучению центральных механизмов
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва)
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва) Изучению реакции организма на длительное голодание посвящено много работ. Большинство из них касается обменных процессов, биохимии синтеза и распада
Что делает нас людьми: кора головного мозга
Что делает нас людьми: кора головного мозга Человеческий мозг – это прежде всего знакомые нам извилины коры, которая выполняет все высшие когнитивные функции. Большую часть коры головного мозга называют неокортекс, потому что эта часть мозга появилась в процессе