Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва)
Изменение активности некоторых Ферментов крови и печени крыс при экспериментальном голодании
А. А. ПОКРОВСКИЙ, Г. К. ПЯТНИЦКАЯ (Москва)
Проблема влияния голодания на разные показатели обменных процессов в организме животных и человека продолжает привлекать внимание большого числа исследователей (1, 3, 7, 11).
Теоретический интерес изучения полного голодания связан с возможностью изучения на его примере адаптации организма к условиям эндогенного питания.
Существует ряд причин повышенного интереса к этой проблеме: а) наличие в наши дни больших контингентов населения некоторых развивающихся стран, страдающих от хронического недоедания; б) факторы вынужденного голодания, сопутствующие ряду заболеваний; в) и, наконец, псе более многочисленные и часто успешные попытки применения голодания с лечебной целью.
Основанием для применения голодания с лечебной целью служит мнение, что в период голодания организм освобождается от избыточных запасов жира и отложившихся в тканях «шлаков», в силу чего оно способствует нормализации обмена веществ.
Не оспаривая эту точку зрения, мы в то же время хотели подчеркнуть неравномерность существующих в организме запасов отдельных пищевых веществ и несомненно, происходящие при этом своеобразном стрессовом состоянии глубокие перестройки многих физиологических и биохимических процессов.
Изучение же поведения различных ферментов крови и печени в голодающем организме способствует более глубокому пониманию своеобразного течения метаболических изменений в организме животных.
Однако, изучению динамики изменения ферментных систем в крови и тканях при полном голодании посвящено немного работ.
Впервые исследования ферментативной активности в печени голодающих крыс были проведены в 1948 году (12).
Было отмечено снижение активности ксантиноксидазы, аргиназы и катепсина, происходящие параллельно уменьшению содержания белка и РНК в органе.
В то же время в некотором противоречии с этими результатами находятся данные некоторых авторов, которые показали повышение лизосомальных ферментов, в частности, катепсина, но отметили резкое снижение активности цитохромоксидазы (5,8).
При изучении поведения гликолитических ферментов было установлено, что активность глюкозо-6-фосфатазы и фруктозо-1,6-дифосфатазы оставалась в пределах нормы или даже несколько повышалась в печени животных при голодании (13, 15—17).
Имеющиеся в литературе данные -о влиянии голодания на активность алиэстеразы довольно противоречивы (6, 9, 14). Нам вовсе не удалось найти сообщений о влиянии голодания на активность метилбутириназы.
Целью настоящего исследования явилось изучение поведения ряда ферментов в крови и печени крыс во время адаптации к острому голоданию.
Изучали активность ФМЭ-1 (фосфогидролазы моноэфиров ортофосфата Кф 3. 1. 3. 1.), ФМЭ-2 (фосфогидролазы моноэфиров ортофосфата Кф 3. 1. 3. 2), ?-аланин аминотрансферазы (Кф 2. 6. 1. 12) и ?-аспарагин аминотрансферазы (Кф 2. 6. 1. 10); алиэстеразы и метилбутириназы (Кф 3. 1. 1. 1.) в крови и печени крыс: и ?-глюкуронидазы (?-D-глюкурюнид-глюкуроногидролазы Кф 3. 2. 1. 31) только в печеночной ткани.
Изучаемые нами ферменты являются представителями различных функциональных классов, характеризуются разной внутриклеточной локализацией, участвуют в различных звеньях сложного метаболического пути превращений веществ в организме.
Определение активности ферментов проводили с помощью ультрамикросистемы, созданной в лаборатории клинической энзимологии Института питания АМН СССР по уже опубликованным прописям (14).
Ферментативную активность выражали количеством микромолей субстрата, разложенного одним граммом сырой ткани или одним миллилитром крови в минуту при 37°С. Полученные результаты обработаны статистически и степень достоверности их была установлена.
Методика опытов
Исследования проводились на половозрелых крысах-самцах породы Вистер весом 200—240 грамм. В начале эксперимента с целью унификации условий опыта в течение 7—10 дней все животные получали 18% синтетическую диету следующего состава:
крахмал — 62 г%
казеин — 18 г%
лярд — 11 г%
дрожжи сухие — 5 г%
солевая смесь — 4 г%
В период эксперимента опытные животные получали только воду без ограничения, контрольные продолжали получать ту же диету. Всего было-исследовано 220 крыс, из них 145 опытных и 75 контрольных.
В зависимости от продолжительности голодания опытные животные были подразделены на следующие группы:
1 группа — 24-часовое голодание
2 группа — 72-часовое голодание
3 группа — 120-часовое голодание
4 группа — 120-часовое голодание с последующим возобновлением кормления 18% синтетической диетой.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Результаты статистической обработки исследований крови и печени животных представлены на рис. 1, 2.
Анализ полученных данных показывает, что уже после 24 часов голодания увеличивается активность липолитических ферментов (алиэстеразы и метилбутириназы) в крови и в печени животных в среднем до 150%.
Активность ФМЭ-2 в крови опытных крыс увеличивается до 125%, в то время как активность ФМЭ-1 снижается до-уровня 65%. Активность ?-глюкуронидазы в печени увеличивается в два раза (до 200%),
При удлинении сроков голодания до 72 часов активность алиэстеразы в крови повышается до 175%, активность метилбутириназы сохраняется на уровне 150%. В печени этих животных активность липолитических ферментов остается повышенной, но наряду с этим наблюдается увеличение активности ферментов переаминирования: аспартат и аланин-аминотрансфераз в крови до 126% и 115%, в печени до 130% и 150% соответственно.
Активность ФМЭ-2 в печени животных данной группы повышена до 167%. Обращает на себя внимание выраженное снижение активности ФМЭ-1 в крови до уровня 45%, в печени до 89%.
Интересно отметить, что после 120 часов голодания крыс активность липолитических ферментов в крови и в печени снижалась до уровня, наблюдаемого у контрольных крыс. Продолжала оставаться низкой активность ФМЭ-1 в крови (44%) и печени (50%) активность ФМЭ-2, ?-глюкуронидазы и ферментов переаминирования в крови и печени сохранялась высокой.
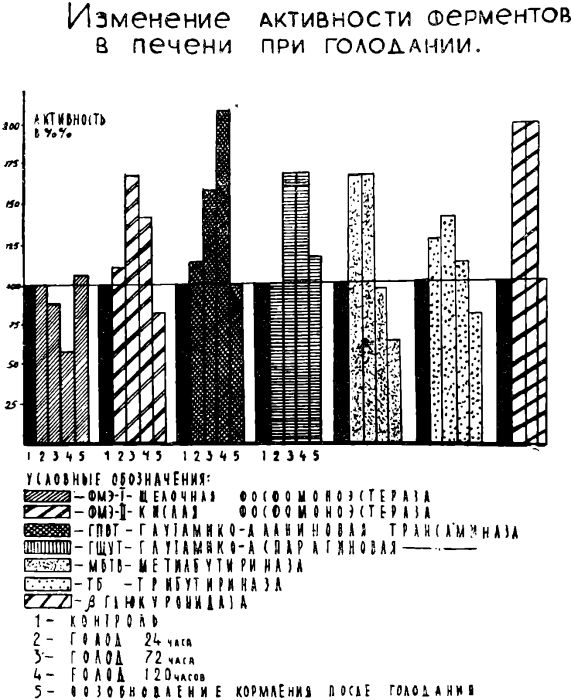
Рис. 1.
Нормальное состояние, голодание и последующее насыщение являются качественно различными состояниями с различными степенями регуляции метаболических процессов, поэтому представляло интерес изучение изменения активности данных ферментных систем при возобновлении кормления. Группа животных после 120 часов голодания получала 18% синтетическую диету в течение 48 часов. При исследовании ферментативной активности в печени и в крови этих животных было отмечено снижение активности аспартат и аланин-аминотрансфераз до уровня контроля. Активность ФМЭ-2 в крови оставалась несколько увеличенной, в то же время значительно повышалась активность ФМЭ-1, достигая уровня контрольного.

Наибольший интерес представляет поведение липолитических ферментов (метилбутириназы и алиэстеразы), активность которых снижается еще в большей степени и достигает уровня 85% и 65% соответственно.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Таким образом, полученные данные отчетливо показывают, что в условиях полного голодания меняется активность изученных нами ферментных систем. В то же время обращает внимание выраженное различие в поведении ферментов, обеспечивающих различные звенья метаболизма клетки. Степень выраженности и характер этих изменений зависят от продолжительности периода голодания.
Вопрос о влиянии голодания на активность алиэстеразы в крови в настоящее время является спорным. Так, у собак в период голодания было обнаружено повышение активности липазы в крови, что, очевидно, связано с усиленной мобилизацией жира из депо и липемией (16). Аналогичное явление было обнаружено и при голодной липемии у людей.
Такая же закономерность была отмечена и у животных при 48—72-часовом голодании (9). С другой стороны, у крыс не наблюдалось сколько-нибудь значительных изменений активности алиэстеразы крови после голодания в течение одной недели (14).
Отмеченные в данной работе изменения активности эсте-раз печени, в частности, ферментов, катализирующих расщепление эфиров глицерина: метилбутириназы и алиэстеразы, можно расценивать как очевидное проявление биохимической адаптации организма в ответ на лишение его пищи и последующее возобновление кормления. Действительно, организм, лишенный пищи, при переходе на эндогенное питание в качестве источников энергии начинает использовать в начальный (очень короткий) период собственные запасы и, в первую очередь, гликоген печени и затем триглицериды жировых депо. Проявлением этой адаптивной перестройки организма является увеличение активности жиромобилизующих ферментов в крови, печени и жировой ткани (2). Однако, вместе с исчерпанием жировых депо происходит быстрое и значительное снижение активности липолитических ферментов.
Пристального внимания заслуживает изучение поведения аминотрансфераз, играющих ведущую роль в клеточном метаболизме. Участвуя в реакциях ферментативного переноса азотосодержащих групп между аминокислотами и соответствующими ?-кетокислотами, стоящими на стыке путей обмена азотистых веществ, углеводов и жиров, они играют определенную роль и в процессах биологического окисления (непрямое окислительное дезаминирование).
Нарастание активности аспартат и аланин-аминотрансфераз в более поздние сроки голодания, вероятно, связано, с одной стороны, с распадом собственного белка, с другой стороны, с ускорением процессов глюконеогенеза, что согласуется с имеющимися в литературе данными некоторых зарубежных авторов (10).
Можно согласиться с мнением некоторых авторов, что повышение активности ферментов переаминирования в печени животных при голодании является результатом синтеза этих ферментов de novo (10), при этом нам бы хотелось подчеркнуть адаптивный характер этого синтеза.
Повышение активности ФМЭ-2 и ?-глюкуронидазы несомненно связано с активацией лизосомальных ферментов и /их выходом в клеточный матрикс.
Полученные данные изменения активности ферментов указывают на существование в организме животных сложного адаптационного механизма, при помощи которого осуществляется его переход на эндогенное питание.
Выводы
Состояние полного голодания в эксперименте на животных характеризуется ранней ферментной перестройкой тканей печени, степень и характер которой зависят от продолжительности голодания.
Наибольшее увеличение активности липолитических ферментов в крови и печени крыс отмечены при 48—72-часовом голодании. Увеличение активности аспартат и аланин-аминотрансфераз наблюдалось в более поздние сроки голодания. Отмечено выраженное снижение активности в крови и печени фосфомоноэстеразы I.
В первые 48 часов возобновления кормления после голодания происходит значительное торможение активности липолитических ферментов, в то время как активность всех остальных изучаемых ферментов возвращается к норме.
ЛИТЕРАТУРА
1. Бакулев А. Н., Колесникова Р. С. «Клиническая медицина», 1962, т. 40, № 2, стр. 14.
2. Лейте с С. М. «Терапевтич. архив», 1962, т. 34, вып. 6, стр. 3.
3. Николаев Ю. С. «Лечение шизофрении дозированным голоданием». В кн.: «Тр. Госуд. НИИ психиатрии МЗ РСФСР», М., 1963, т. 39, стр. 7.
4. Покровский А. А. Руководство по изучению питания и здоровья населения, М., 1964.
5. Аllагd С., De Lomirande, Cantero А. Ехрег. cell research, 1957, N. 1, 69.
6. Abderhalden E. Schutzfermente des tierischen organismus. Ein Beitrag zur Kenntnisder Adwehrmassregeln de tierischen organismus gegen Korperblut and zellfremde stoffe. Berlin, Springer, 1912.
7. Вlооm W. L. Ann. N. J. Aced. Sci. vol., 1965, 131, 623.
8. Beaufay H, d e D u v e. Biochem. y. vol. 1959, 73, 4, 617.
9. Constantinides P., John Stone F. R. Proc. Soc. Exp. Biol. (N. J.), 1959, 100, 262.
10. Sober on G., Sanchez E. Y. Biolog. Chem., 1961, 236, N 5.
11. Duncan G. G., W. K. Yensonet. al. Amer. your, or Medical Science, 1963, 245, N. 5, 35.
12. Miller L. L., Y. Biolog. Chem., 1948, 172, 113.
13. Niemeyer G. H. Acta physiol. latino-amer., 1962, 12, 173.
14. Tuba Т., Ho are R. Canad. Y. Med. Science, 1951, 29, 1, 25.
15. Weber G. and Cant его A. Science, 1954, m. 120, p. 851.
16. Weber G. «Canad. Y. Biochem.», 1965, 43, N. 9, 1549.
17. Weber G. and Cantero A. Endocrinology, 1957, 61, 5—6,
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 4. Регуляция активности ферментов. Медицинская энзимология
Глава 4. Регуляция активности ферментов. Медицинская энзимология Способы регуляции активности ферментов:1. Изменение количества ферментов.2. Изменение каталитической эффективности фермента.3. Изменение условий протекания реакции.Регуляция количества
К характеристике изменения ЭЭГ у больных, находящихся на длительном лечебном голодании В. В. АРШАВСКИЙ, Б. В. КРАЙЦЕРОВ (Москва)
К характеристике изменения ЭЭГ у больных, находящихся на длительном лечебном голодании В. В. АРШАВСКИЙ, Б. В. КРАЙЦЕРОВ (Москва) Из многочисленных данных литературы известно, что центральный отдел висцерального анализатора как у животных, так и у человека проецируется в
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва)
Особенности возбуждения коры головного мозга у животных при длительном голодании Т. Н. ЛОСЕВА, А. А. ПАНФИЛОВ (Москва) Изучению реакции организма на длительное голодание посвящено много работ. Большинство из них касается обменных процессов, биохимии синтеза и распада
Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва)
Эритрон при длительном алиментарном голодании и последующем питании людей Н. А. ФЕДОРОВ, Ю. Л. ШАПИРО (Москва) Голодание, как частный раздел науки о питании, за последние годы вновь стало привлекать к себе пристальное внимание в связи со все более широким его
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва)
Интенсивность эритродиеретических процессов при голодании людей (математический анализ) Ю. Л. ШАПИРО, В. М. ЛУГОВСКОЙ (Москва) Как известно, при полном алиментарном голодании в течение довольно длительного времени (по крайней мере до сроков, характеризующихся потерями
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва)
Показатель степени насыщенности крови кислородом при лечебном голодании В. Б. ГУРВИЧ, Ю. Л. ШАПИРО, М. В. САМОЙЛОВА (Москва) В литературе имеется значительное число работ, посвященных изучению изменения степени насыщения крови кислородом при разнообразных условиях и
Динамика показателей периферической крови при лечебном голодании у больных гипертонической болезнью и ожирением Г. Н. БЖИШКЯН-БОРОДИНА (Москва)
Динамика показателей периферической крови при лечебном голодании у больных гипертонической болезнью и ожирением Г. Н. БЖИШКЯН-БОРОДИНА (Москва) Литературные данные относительно исследования состава периферической крови при лечебном голодании представляют большую
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва)
Фагоцитарная активность лейкоцитов периферической крови при полном голодании и последующем питании людей Ю. Л. ШАПИРО, Ю. С. НИКОЛАЕВ, А Я. ТАБАХ, Л. Ф. ЛЕВИНА (Москва) Изучению фагоцитарной активности лейкоцитов при длительном полном алиментарном голодании посвящены
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва)
pH сыворотки крови больных при лечебном голодании В. А. СКОРИК-СКВОРЦОВА, В. А. КУЛАЧКОВ (Москва) Известно, что так называемые «жесткие константы» сохраняются неизменными в организме В течение длительного срока, несмотря на воздействие каких-либо «отклоняющих» факторов.
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва)
Изменение количества и некоторых параметров полового хроматина у людей в процессе полного длительного алиментарного голодания и последующего питания С. Н. РЕЗИНА, Ю. Л. ШАПИРО (Москва) Половой хроматин — внутриядерное тельце, дающее Фельген-положительную реакцию и
О динамике основного обмена при лечебном голодании больных с ипохондрическим синдромом В. Б. ГУРВИЧ (Москва)
О динамике основного обмена при лечебном голодании больных с ипохондрическим синдромом В. Б. ГУРВИЧ (Москва) Вопрос об обмене веществ, в частности, основном обмене, является одним из ведущих при объяснении патофизиологических механизмов голодания (1, 5, 9, 11, 12, 14).В. В.
СОСТОЯНИЕ ИММУНОБИОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ ОРГАНИЗМА ЧЕЛОВЕКА ПРИ ПОЛНОМ ДЛИТЕЛЬНОМ ГОЛОДАНИИ Ю. С. НИКОЛАЕВ, В. А. СКОРИК-СКВОРЦОВА (Москва)
СОСТОЯНИЕ ИММУНОБИОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ ОРГАНИЗМА ЧЕЛОВЕКА ПРИ ПОЛНОМ ДЛИТЕЛЬНОМ ГОЛОДАНИИ Ю. С. НИКОЛАЕВ, В. А. СКОРИК-СКВОРЦОВА (Москва) Изучение состояния иммунобиологической реактивности организма в период длительного полного голодания и последующего
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва)
Материалы к изучению ферментной адаптации при полном лечебном голодании А. А. ПОКРОВСКИЙ, Ю. С. НИКОЛАЕВ, Г. К. ПЯТНИЦКАЯ, Г. И. БАБЕНКОВ (Москва) В течение последних лет в нашей стране и за рубежом появилось большое число сторонников применения голодания с лечебной целью при
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва)
Влияние полного экспериментального голодания на неосаждаемую активность некоторых лизосомальных гидролаз А. А. ПОКРОВСКИЙ, Г. К ПЯТНИЦКАЯ (Москва) В связи с тем вниманием, которое в настоящее время уделяется роли лизосом в развитии повреждения клеток тканей различных
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва)
Химический состав тканей крыс при полном голодании В. И. ДОБРЫНИНА (Москва) Голодание как метод лечения успешно зарекомендовал себя при некоторых психических и соматических заболеваниях (3, 7, 10—13). Особенно перспективно его применение при обменных, аллергических
Влияние голодания на активность ферментов пентозофосфатного пути в печени и мозге крыс Ю. Л. ЗАХАРЬИН (Москва)
Влияние голодания на активность ферментов пентозофосфатного пути в печени и мозге крыс Ю. Л. ЗАХАРЬИН (Москва) В последние годы в клинике часто применяется с лечебными целями, в частности, для лечения психических заболеваний, полное голодание. Не вызывает сомнения, что