ГЛАВА 6 Поздний докембрий: возникновение многоклеточности. Гипотеза кислородного контроля. Эдиакарский эксперимент
ГЛАВА 6
Поздний докембрий: возникновение многоклеточности. Гипотеза кислородного контроля. Эдиакарский эксперимент
Прежде чем непосредственно приступать к изучению древнейших многоклеточных организмов, давайте задумаемся: а зачем, собственно говоря, эта многоклеточность нужна? Какие преимущества она дает? На эту проблему можно посмотреть с разных сторон. Начнем с экологической точки зрения: что многоклеточность дает не самому организму, а экосистеме и (в конечном счете) биосфере.
Малоразмерные фитопланктонные организмы (прежде всего прокариотные) сталкиваются с одной проблемой, на которую впервые обратил внимание гидробиолог Б. Я. Виленкин. Снабжение этих одноклеточных биогенами и растворенными газами происходит за счет диффузии сквозь клеточную стенку. Казалось бы, это должно благоприятствовать миниатюризации клеток, максимальному увеличению отношения ее поверхности к объему. Однако очень мелкий организм неспособен выделиться из окружающей его водной массы. Пассивно паря в толще воды и оставаясь неподвижным относительно нее, он быстро создает вокруг себя «пустыню» — выедает из непосредственно окружающего его водного слоя все биогены, насыщая воду диффундирующей наружу органикой, которую не в силах удерживать внутри клетки из-за того же (высокого) соотношения «поверхность — объем». При этом безвозвратно теряется более 1/3 клеточной продукции. Но хуже то, что на окисление этой органики расходуется кислород и водный слой, окружающий организм, эвтрофицируется. Милый Пятачок прав: жизнь «очень маленького существа» полна неприятностей.
Организм избежит неприятностей, если будет перемещаться относительно вмещающей его воды. Этого он может добиться двумя способами: либо начнет сам активно двигаться в ее толще, либо прикрепится к неподвижному субстрату, чтобы вода двигалась относительно него. Второй способ прокариоты реализовали немедленно, сконструировав мат. А вот с первым способом (казалось бы, более простым и очевидным) у них возникли большие проблемы, ибо эффективных органов движения (подобных жгутикам и ресничкам эукариотных одноклеточных) у прокариот не возникает, а создание клеточных агрегаций, способных к согласованным движениям (например, волнообразным) затруднено из-за крайней слабости межклеточных взаимодействий (см. главу 5). Поэтому «генеральной линией» этот способ становится лишь при появлении эукариот, способных к образованию высокоинтегрированных клеточных агрегаций, т.е. к настоящей многоклеточности.
Многоклеточность привела, среди прочего, к резкому повышению способности организмов создавать в своем теле запас питательных веществ. Практически лишенные этих запасов одноклеточные (в особенности прокариоты) обречены реагировать на изменения содержания биогенов в окружающей среде единственным способом — изменением плотности популяций. (Примером такого их поведения может служить «цветение» водоемов, когда стремительное размножение одноклеточных и нитчатых водорослей быстро приводит к исчерпанию ресурса экосистемы; в результате безмерно разросшаяся популяция в одночасье вымирает, а на окисление этой мертвой органики расходуется затем почти весь кислород водоема — «замор».) Запасные вещества и резервная биомасса крупных организмов делают их популяции более независимыми от колебаний ресурса и стабилизируют их плотность. В свою очередь, существующие в экосистеме геохимические круговороты обретают в лице этих организмов крупное резервное депо,[16] оказывающее на все эти циклы мощное стабилизирующее воздействие. Такая «взаимная стабилизация» кажется одним из главных экологических следствий возникновения многоклеточных (хотя правильнее сказать — макроскопических, т.е. видимых невооруженным глазом) организмов.
Последняя оговорка неслучайна. Если мы от вопроса «для чего возникла многоклеточность?» перейдем к вопросу «как и когда она возникла?», то нам прежде всего придется определить, какой организм следует считать многоклеточным. Если даже не брать в расчет чисто терминологические проблемы (следует ли называть «колонией» Volvox, у которого есть дифференциация между клетками переднего и заднего полушарий, зародыши, возникающие путем палинтомии — деления без последующего увеличения размеров клеток, и который к тому же смертен), здесь существует и проблема вполне объективная. А именно: соответствует ли строгим критериям многоклеточности строение тела макроскопических водорослей и грибов? Большинство биологов ныне отвечают на этот вопрос отрицательно, оставляя термин «многоклеточные» лишь за зелеными растениями (Metaphyta) и животными (Metazoa); Дж. Корлисс (1983) ввел для этих двух групп удачный термин — многотканевые организмы.
Таким образом, достижение живыми организмами макроскопических размеров может происходить различными способами, а истиная многоклеточность (многотканевость) — лишь один из них. В любом случае, многоклеточность (в широком смысле) почти наверняка возникала в процессе эволюции многократно и независимо: во многих типах водорослей — красных (Rhodophyta), зеленых (Chlorophyta), золотистых (Chrysophyta) — или высших грибов-аскомицетов (Ascomyceta) можно выявить преемственные ряды от одноклеточных форм через колонии (например, нитчатые формы) к макроскопическим организмам с аналогами тканевой дифференцировки.
Эта гипотетическая эволюционная картина вполне соответствует современным палеонтологическим даным (рис. 19). Первые эукариоты появились (как мы знаем из главы 5) около 2 млрд лет назад среди фитопланктонных акритарх; вскоре к ним добавились и нитчатые формы с эукариотными параметрами клеток. Замечательно, что они никогда не встречаются в прокариотных бентосных сообществах (цианобактериальных матах), а с самого начала формируют свой собственный тип растительности, названный У. Шенборном (1987) «водорослевыми лугами». Ныне сообщества такого типа известны лишь в некоторых антарктических внутренних водоемах; в докембрии же они, судя по характеру осадков, были широко распространены в морях за пределами мелководий (которые были заняты матами).
В рифее (1,4–1,2 млрд лет назад) эти нитчатые эукариотные формы достигли значительного разнообразия. Именно тогда в составе водорослевых лугов появились и первые макроскопические водоросли с пластинчатым, корковым и кожистым типами слоевища, а к венду (650 млн лет) основным компонентом этих сообществ становятся вендотении — лентовидные водоросли длиной до 15 см. Есть даже сообщение о находке каких-то пластинчатых водорослей в китайской формации Чанчен с возрастом 1,8 млрд лет; эта датировка нуждается в подтверждении, но не кажется нереальной. Дело в том, что таксономическая принадлежность всех этих форм недостаточно ясна, однако некоторые из них очень сходны с низшими красными водорослями — бангиевыми. А красные водоросли, как полагают многие исследователи, являются самой архаичной ветвью эукариот; возможно, они произошли в результате «независимой эукариотизации» цианобактерий.
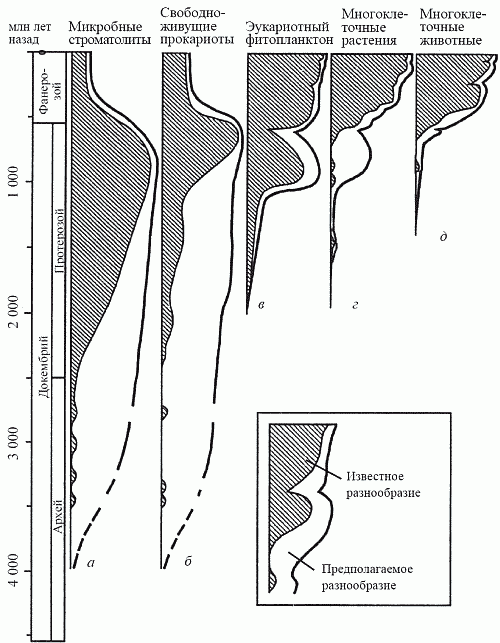
Рис. 19. Изменения во времени относительного обилия основных биотических компонентов
Рис. 19. Изменения во времени относительного обилия основных биотических компонентов: а — прокариоты-строматолитообразователи; б — свободноживущие прокариоты; в — эукариотный фитопланктон; г — многоклеточные растения; д — многоклеточные животные (по Schopf, 1992)
Животные и следы их жизнедеятельности (норки и следовые дорожки на поверхности осадка) достоверно появились в палеонтологической летописи лишь в конце протерозоя — около 800 млн лет назад. (Интересно, что водоросли с минерализованными слоевищами, для которых можно предполагать тот же уровень организации, что и у высших красных и бурых водорослей, появились еще позднее — в венде.) В чем же причина того, что истиная многоклеточность возникла так поздно? В 60-е годы прошлого века (вспомним главу 5) существовала гипотеза «кислородного контроля» Беркнера и Маршалла, согласно которой содержание кислорода в земной атмосфере вплоть до начала фанерозоя (540 млн лет назад) было ниже точки Пастера и не допускало существования более высоко организованных форм жизни, чем водоросли. Поскольку со временем установили, что точка Пастера в действительности была пройдена гораздо раньше — более чем за миллиард лет до времени появления первых многоклеточных, причинная связь между этими явлениями была отвергнута и о гипотезе «кислородного контроля» забыли. Как позже выяснилось, напрасно.
Однопроцентное содержание кислорода (имеется в виду 1% от его современного количества) — это тот критический минимум, ниже которого аэробный метаболизм принципиально невозможен; однако для жизнедеятельности макроскопических животных кислорода необходимо существенно больше. Б. Раннегар недавно провел специальные расчеты, из которых следует, что животным, составлявшим первую фауну многоклеточных — эдиакарскую (о ней речь впереди), кислорода требовалось не менее 6–10% от нынешнего уровня; это в том случае, если они имели развитую систему циркуляции, доставлявшую кислород к тканям. Если же такая система у них еще не развилась и они дышали за счет прямой диффузии (скорее всего, так и было), то требовалось для их жизнедеятельности гораздо больше кислорода (может быть, его содержание сопоставимо с нынешним). Итак, гипотеза «кислородного контроля» кажется вполне логичным объяснением появления макроскопических животных лишь в конце протерозоя — если принять более высокий критический порог, чем однопроцентный, предлагавшийся Беркнером и Маршаллом. Однако возможна ли проверка этой гипотезы? Мы помним (из главы 5), что можно по составу осадков отличить аэробные обстановки от анаэробных, но как определить количественные различия в содержании кислорода в былые эпохи?
Известно, что количество кислорода, создаваемого небиологическими процессами (фотолиз воды и т.д.), совершенно ничтожно; почти весь свободный кислород планеты создан фотосинтезирующими организмами. Однако живые существа не только производят кислород, но и потребляют его при дыхании. В биосфере осуществляется достаточно простая химическая реакция: nСО2+nH2O = (CH2O)n + nО2. «Читая» ее слева направо, мы получаем фотосинтез, а справа налево — дыхание (а также горение и гниение). Уровень содержания кислорода на планете стабилен потому, что прямая и обратная реакции взаимно уравновешиваются; так что если мы попытаемся увеличить содержание свободного кислорода в атмосфере путем простого наращивания объема фотосинтезирующего вещества, то из этой затеи ничего не выйдет. Сместить химическое равновесие, как вам должно быть известно из курса химии, можно, лишь выводя из сферы реакции один из ее продуктов. В нашем случае добиться увеличения выхода О2 можно, лишь необратимо изымая из нее восстановленный углерод в форме (CH2O)n или его производных.
Таким образом, производство кислорода биосферой начинает превалировать над потреблением этого газа (ею же), только если происходит захоронение в осадках неокисленного органического вещества. Этот вывод кажется достаточно парадоксальным и противоречащим расхожим представлениям. Так, например, природоохранная пропаганда вбила в голову широкой публики магическую фразу «Леса — это легкие планеты»; и мало кто дает себе труд задуматься над тем, что в действительности любое сбалансированное сообщество (в том числе тропический лес) потребляет ровно столько кислорода, сколько и производит. Если что и можно назвать «легкими планеты», так это болота, где как раз и идет процесс захоронения неокисленной органики. Поэтому, если мы установим, что в некий период геологической истории происходило интенсивное захоронение органического углерода, то мы вправе будем заключить, что в это время столь же интенсивно накапливался и кислород. А вот оценить темпы захоронения органического углерода в прошлые эпохи вполне возможно. Для этого существует специальный метод, основанный на изучении изотопного отношения 12C/13C в соответствующих осадках (органический углерод, участвовавший когда-либо в реакциях фотосинтеза, обогащен «легким» изотопом 12C).
Несколько лет назад на архипелаге Шпицберген была открыта уникальная по полноте последовательность позднепротерозойских осадков, отлагавшихся в период с 850 до 600 млн лет назад. Исследовав эти осадки на предмет изотопного отношения 12C/13C, Э. Нолль (1996) установил, что на протяжении всего этого времени темпы захоронения органического углерода оставались самыми высокими за всю историю Земли. Изучение позднепротерозойских осадков в других районах мира — в Канаде, Австралии и Южной Африке — подтвердило выводы Нолля. Итак, появлению макроскопической фауны предшествовало резкое увеличение количества свободного кислорода; вряд ли можно счесть это случайным совпадением (рис. 20).
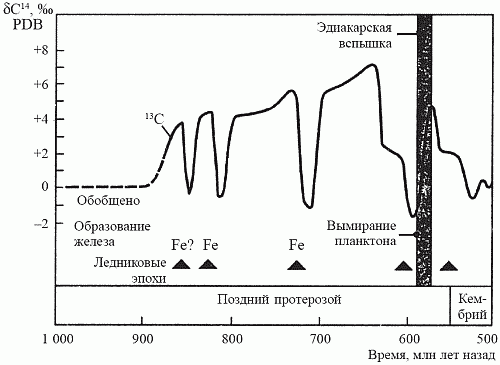
Рис. 20. Ключевые характеристики истории позднего протерозоя
Рис. 20. Ключевые характеристики истории позднего протерозоя. Минимумы на кривой захоронения органического углерода соответствуют ледниковым эпохам и интенсивному образованию железорудных формаций (по Ноллю, 1995)
О том, что на Земле в это время действительно существенно возросло содержание кислорода, могут свидетельствовать и другие факты. Во-первых, в океанах вновь происходит осаждение джеспеллитов, прекратившееся около 1,8 млрд лет назад (см. рис. 16). Это может означать, что кислород впервые насытил глубоководные части океана, располагающиеся ниже фотической зоны, и при этом произошло осаждение еще сохранявшихся в тех глубинах запасов закисного железа. Во-вторых, это было время как минимум трех покровных оледенений, последнее из которых — Лапландское оледенение (600 млн лет назад) — было, по всей видимости, самым крупным за всю историю Земли. Одной из причин начала оледенений считают так называемый обратный парниковый эффект: когда в атмосфере планеты отношение О2/СО2 смещается в сторону кислорода, планета начинает гораздо хуже удерживать тепло, доставляемое центральным светилом.
Все это, вероятно, стало следствием событий, произошедших в биосфере около 1,2 млрд лет назад, когда начался постоянный рост численности и видового разнообразия эукариотного фитопланктона (закончился 600 млн лет назад). Известно, что в планктонных экосистемах детритная пищевая цепь развита очень слабо, а оборот вещества и энергии идет почти исключительно по пастбищной цепи; практически вся мертвая органика «проваливается» из экосистемы вниз, на дно водоема, где и утилизуется в тамошних детритных цепях. Однако донные детритные цепи протерозоя существуют в условиях сильного недостатка кислорода и потому обладают ограниченной «пропускной способностью». Начиная с некоторого момента они перестают справляться с тем нарастающим потоком органики, что поставляется сверху наращивающими свою продуктивность фитопланктонными сообществами. Другим источником органики могут быть лишенные в ту пору сколь-нибудь эффективных консументов («травоядных») водорослевые луга. В результате органический углерод начинает захораниваться в осадках на дне океана; оборотной стороной этого процесса является, как мы теперь знаем, накопление свободного кислорода, приводящее — через обратный парниковый эффект — к оледенению.
Сочетание этих двух факторов (заметный рост содержания кислорода и появление ледников) создает в гидросфере планеты принципиально новую ситуацию. До сих пор кислород мог распространяться из фотической зоны, где он вырабатывается, в нижележащие слои океана только за счет такого несовершенного механизма, как диффузия: ведь более богатая кислородом вода поверхностного слоя — более теплая, а потому она не «тонет». Заметим, что само «богатство» это весьма относительное, т.к. чем теплее вода, тем меньшее количество газа (при равном давлении) может быть в ней растворено. Известно, что вода имеет минимальную плотность при температуре 0°C и ниже (лед), а максимальную — при 4°C, поэтому с появлением на планете льда и «тяжелой» (четырехградусной) воды картина меняется. Плавающий по поверхности лед охлаждает омывающую его воду, и когда ее температура достигает 4°C, она «тонет» — вместе с растворенными в ней газами из поверхностного слоя; в океане образуется холодный придонный слой — психросфера. Таким образом, в гидросфере возникает «ленточный транспортер», доставляющий кислород в ее глубокие слои (одним из следствий этого процесса стало осаждение в океанах джеспеллитов). Оледенения случались на Земле и раньше (например, Гуронское оледенение 2,3–2,5 млрд лет назад), но тогда этому транспортеру еще нечего было переносить; теперь же (650–600 млн лет назад) вся гидросфера оказалась насыщенной кислородом до того уровня, который допускает существование макроскопических животных.
В 1947 году Р. Спригг сделал в местечке Эдиакара в Южной Австралии одно из самых замечательных открытий за всю историю палеонтологии. Им была найдена и описана богатая фауна удивительных бесскелетных организмов, получившая название эдиакарской. Первоначально эту фауну сочли кембрийской, однако в 1959 году М. Глесснер правильно датировал ее концом докембрия — вендом (620–600 млн лет назад); таким образом, период достоверного существования на Земле многоклеточных животных удлинился почти на 100 млн лет. В дальнейшем эдиакарскую фауну нашли еще в нескольких районах мира (Намибия, Ньюфаундленд); более того, выяснилось, что этих существ находили и ранее (например, на Украине в 1916 году), но принимали за неорганические остатки. Самое же богатое и наиболее информативное в плане палеоэкологии местонахождение (многие тысячи особей, относящихся к нескольким десяткам видов) известно в России, на Белом море.
В то время как все многочисленные группы многоклеточных, появившиеся в начале кембрия (540 млн лет назад), были представлены мелкими организмами (миллиметры или первые сантиметры), эдиакарская фауна состояла из крупных или очень крупных беспозвоночных, размером до полутора метров. Среди них были как радиально-симметричные формы, называемые медузоидами, так и билатерально-симметричные; одни из них (петалонамы) внешне напоминают современные кораллы морские перья (Pennatularia), другие (дикинсония и сприггина) — кольчатых червей и членистоногих (рис. 21). Первые исследователи эдиакарской фауны (Спригг и Глесснер) считали эти формы реальными предками современных кишечнополостных и червей и включали их в состав соответствующих типов и классов животных. Эта точка зрения имеет сторонников и поныне («австралийская школа»), однако большинство исследователей полагает, что сходство тут чисто внешнее и что эдиакарские организмы представляют собой нечто совершенно особенное. А. Зейлахер (1984) ввел для этих существ специальное собирательное название — вендобионты.
Вендобионты имеют отличный от фанерозойских животных план строения. Почти у всех билатерально-симметричных вендских организмов эта самая симметрия несколько нарушена; у «членистых» форм правые и левые половинки «сегментов» смещены друг относительно друга — примерно так, как в застегнутой молнии или на автомобильном протекторе-елочке (см. рис. 21, а). Эту асимметрию принято было приписывать деформациям тел в процессе захоронения, пока М. А. Федонкин (1983) не обратил внимание на то, что данные нарушения подозрительно регулярны и единообразны. Он доказал, что для вендобионтов характерен особый план строения, который математики называют симметрией скользящего отражения; среди многоклеточных же животных этот тип симметрии встречается крайне редко (таково, например, взаиморасположение мышечных элементов — миомеров и миосепт — у ланцетника).
Брюс Раннегар (1982) показал, что у изученных им вендобионтов увеличение размеров тела в ходе онтогенеза (инивидуального развития организма) достигается за счет изометрического роста, когда все пропорции тела остаются неизменными (как при простом увеличении изображения предмета). Между тем, все известные метазоа, включая кишечнополостных и червей, обладают не изометрическим, а аллометрическим ростом с закономерной сменой пропорций тела (например, у человека в ряду «зародыш — ребенок — взрослый» абсолютный размер головы увеличивается, тогда как относительный — уменьшается).

Рис. 21. Эдиакарская фауна
Рис. 21. Эдиакарская фауна: а — различные радиально– и билатерально-симметричные формы;б — строение дикинсонии (по Dzik, 1997)
Против отнесения эдиакарских организмов к современным таксонам животных есть возражения и более частного характера. Так, например, попытка напрямую связать вендских медузоидов с настоящими медузами наталкивается на отсутствие у этих вендобионтов стрекательных клеток, которые по идее должны хорошо сохраняться в ископаемом состоянии. Под давлением подобных аргументов сторонники прямого родства между эдиакарскими и фанерозойскими животными начали «сдавать» вендобионтов по одному («Да, похоже, что сприггина все-таки не настоящее членистоногое…»), и это продолжалось до тех пор, пока А. Зейлахер (1984) не предложил принципиально иное решение этой проблемы. Суммировав отличия вендских животных от их современных аналогов, он привел и общую для них черту: они представляют собой различные варианты широкой ленты со вздутиями (рис. 21, б). Этот тип организации (Зейлахер назвал его «стеганое одеяло») совершенно отличен от всех ныне существующих. Судя по всему, такой план строения — это особый путь к достижению больших размеров тела именно бесскелетными формами.
При увеличении размеров тела происходит относительное уменьшение его поверхности. Поскольку многие функции организма (газообмен, питание) прямо связаны с его поверхностью, ее относительный размер нельзя уменьшать до бесконечности: есть определяемый «из физики» физиологический предел размеров тела. Одно из решений этой проблемы нашли современные (фанерозойские) многоклеточные: сохранить в общем округлое сечение тела, но развить выросты или впячивания с большой поверхностью — жабры или легкие, сложноскладчатую пищеварительную систему и т.д. Однако возможно и принципиально иное решение: превратить все тело в плоскую ленту, приблизив таким образом внутреннюю поверхность к внешней; тогда можно достигнуть больших размеров и не создавая сложных внутренних структур. Это решение используется ныне очень редко (по этому пути пошли некоторые крупные кишечные паразиты).
Форма тела вендобионтов («стеганое одеяло») с ее очень высоким отношением «поверхность — объем» позволяла им, как полагает Зейлахер, поглощать из воды кислород и метаболиты всей поверхностью тела. Эта идея находится в полном соответствии с тем обстоятельством, что у самых крупных эдиакарских организмов нет никакого подобия рта. Питаясь за счет диффузии через поверхность тела, эти существа не нуждались во внутренних органах; возможно, они даже были не многоклеточными, а синцитиальными (многоядерными). Кроме того, многие исследователи полагают, что эти плоские (и, видимо, прозрачные) существа были буквально нашпигованы симбиотическими одноклеточными водорослями, что делало их практически независимыми от внешних источников пищи. Их современным экологическим аналогом могут служить так называемые «автотрофные животные» — глубоководные погонофоры и некоторые моллюски, использующие при питании симбиотических бактерий-хемоавтотрофов.
Итак, на мелководьях вендских морей (именно здесь, судя по характеру вмещающих осадков, обитали вендобионты) существовала удивительная экосистема из «автотрофных животных». Сейчас известны тысячи экземпляров различных представителей эдиакарской фауны, однако ни на одном из них нет повреждений и следов укусов; судя по всему, в это время не существовало хищников, да и вообще животных, питающихся крупными кусками пищи. По образному выражению С. Гоулда, в венде трофические цепи были столь коротки, что полностью помещались внутри тела консумента. Поэтому вендскую биоту часто называют, вслед за М. Мак-Менамином, «Сад Эдиакары» — по аналогии с райским садом, где никто никого не ел. Ситуация райского сада, как ей и положено, просуществовала недолго: в конце венда вендобионты полностью вымерли, не оставив после себя прямых потомков. Эдиакарский эксперимент — первая в истории Земли попытка создать многоклеточных животных — окончился неудачей.
Впрочем, по поводу судьбы эдиакарской фауны есть и иные мнения. Помимо двух диаметрально противоположных позиций (австралийской школы и Зейлахера) есть и «компромиссная», на которой стоят, например, Федонкин и Раннегар. Эти исследователи считают, что в составе эдиакарской фауны помимо вендобионтов, уникальных по своей организации и свойственных только для этого времени (а также, возможно, и реликтов каких-то довендских фаун), имеются и отдаленные предки некоторых фанерозойских групп.
В этой связи можно вспомнить элементы характерной для вендобионтов симметрии скользящего отражения, наблюдаемые в строении примитивнейшего из хордовых — ланцетника. С другой стороны, один из эдиакарских организмов — ярнемия, с ее мешковидным телом и двумя «сифонами», очень напоминает близкого родственника хордовых — асцидию; к тому же оказалось, что отпечатки этого организма сильно обогащены ванадием — тем самым металлом, что является основой дыхательного пигмента асцидий. Так что некоторые исследователи не исключают, что мы с вами (как представители хордовых) ведем свой род прямиком от вендобионтов.
Это, впрочем, еще не самая экзотическая гипотеза относительно природы и родственных связей вендобионтов. Кем только их не объявляли — даже гигантскими морскими лишайниками! Например, А. Ю. Журавлев (1993) предложил весьма остроумную гипотезу относительно связей некоторых эдиакарских организмов с гигантскими (до 20 см в диаметре) многоядерными простейшими из подкласса ксенофиофор (класс саркодовых).[17] А недавно Д. В. Гражданкин и М. Б. Бурзин (1998) привели аргументы в пользу того, что тела вендо-бионтов представляли собой вовсе не толстое «стеганое одеяло», а тонкую гофрированную мембрану — ее по аналогии можно назвать «картонкой для яиц»; в захоронение попадает, собственно, не сама мембрана, а те «куличики из песка», которые получаются при засыпании ее «лунок» взмученным осадком. Эти «картонки для яиц», имеющие идеальное соотношение «объем — поверхность», неподвижно лежали на дне, поглощая из морской воды растворенную в ней органику…
Подобный разнобой в гипотезах может произвести на стороннего наблюдателя удручающее впечатление, однако «в оправдание» ученых, изучающих докембрийские организмы, необходимо сказать следующее. Решаемая ими задача является, возможно, самой сложной во всей палеонтологии, ибо актуалистический метод реконструкций явно работает здесь на самом пределе своей разрешающей способности. Палеонтологи фактически находятся в положении космонавтов, столкнувшихся с фауной чужой планеты — с тем лишь уточнением, что они вынуждены иметь дело не с самими инопланетными существами, а с созданным теми «театром теней».
Не очень давно выяснилось, что эдиакарская попытка создания многоклеточных животных была не первой по счету, а в лучшем случае второй. В 1986 году Сун Вэйго обнаружил в Китае (Хайнань) в позднерифейских отложениях с возрастом 840–740 млн лет богатую фауну макроскопических бесскелетных животных; ее, по традиции, назвали хайнаньской биотой. Почти одновременно сходная фауна была найдена М. Б. Гниловской в России, на Тиманском кряже; к сожалению, изучена она пока недостаточно. Известно лишь, что составляющие ее организмы уступают по размеру эдиакарским и являются не лентовидными, а червеобразными, причем иногда кажутся членистыми; многие из них строят из органического материала сегментированные трубки бокаловидной формы. Очень важно то, что среди этих организмов нет ни медузоидов, ни каких-либо форм, близких к губкам; судя по всему, протерозойская хайнаньская биота не может рассматриваться как предковая ни для эдиакарской, ни для современной (фанерозойской).
Из этого, правда, не следует, что все представители хайнаньской биоты должны были вымереть до эдиакарской попытки; возможно, именно им принадлежит часть следов рытья в вендских осадках. Однако в целом хайнаньский эксперимент явно оказался столь же неудачным, как и эдиакарский, хотя, возможно, по совершенно иным причинам. Обращает на себя внимание тот факт, что хайнаньская попытка совпадает по времени (840 млн лет назад) с началом «кислородного бума». Впрочем, существование макроскопических водорослей в отложениях возраста 1,4 млрд лет наводит на мысль о том, что и эта попытка создания многоклеточных животных может оказаться не самой первой.[18]
Итак, мучивший Дарвина вопрос о появлении животных в начале кембрия («всех и сразу») вроде бы начал проясняться. Преподавателям теории эволюции на первый взгляд полегчало: как-никак, этот рубеж сильно отодвинулся в глубь времен (с 540 до 840 млн лет назад), а первые животные оказались действительно сильно непохожими на тех, что существуют, начиная с кембрия. Но это только на первый взгляд. Ведь новооткрытые-то фауны оказались непреемственными между собой; они всякий раз появляются, как чертик из коробочки, и туда же исчезают. Ситуация, если вдуматься, складывается еще более скандальная, чем во времена Дарвина: тот имел дело с одним «актом творения» — кембрийским, а мы — как минимум с тремя. М. Б. Бурзин дал эдиакарской и хайнаньской попыткам остроумное определение: «черновики Господа Бога».
Наиболее логичным объяснением тут может быть признано такое. На Земле с достаточно давних времен существовала развитая фауна микроскопических бесскелетных животных (уровня простейших, коловраток и т.п.). Ее представители периодически совершали попытки выйти в крупный размерный класс, но лишь последняя из этих попыток — кембрийская — оказалась вполне успешной. Шансов попасть в захоронение у этих существ нет никаких (ведь от фитопланктонных организмов остается только целлюлозная оболочка либо слизистый чехол, которых животные лишены), а потому их отсутствие в палеонтологической летописи не должно нас смущать. К сожалению, последнее обстоятельство делает данную гипотезу непроверяемой, а цена таких гипотез, как мы помним из дополнения к главе 1, крайне мала. Впрочем, если в палеонтологической летописи не сохраняются сами эти организмы, нельзя ли попытаться обнаружить в ней какие-нибудь следы их жизнедеятельности или иные косвенные указания на их существование?
Действительно, в соответствующих осадках часто находят мелкие или мельчайшие гранулы обогащенного органикой вещества, трактуемые как фекальные пеллеты (пища, прошедшая через организм неких консументов). Однако более интересное решение этой проблемы было недавно предложено М. Б. Бурзиным (1987). Основываясь на эмпирических данных о распространении в геологической истории основных размерных классов и морфологических групп фитопланктона, он показал, что тенденции, выявленные в эволюции планктонного сообщества, могут объясняться лишь наличием в его составе консументов, и даже «вычислил» некоторые их характеристики (подробнее см. в дополнении к главе 6). В качестве первичного биотопа этих существ обычно указывают уже знакомые нам водорослевые луга; в этих густых подводных зарослях должна была наблюдаться повышенная концентрация кислорода.
Судя по всему, именно деятельность зоопланктона спровоцировала вендский фитопланктонный кризис — первое палеонтологически документированное крупномасштабное вымирание (см. дополнение к главе 6). Затем бесследно исчезла эдиакарская фауна, а позже произошло стремительное уменьшение численности и разнообразия строматолитовых построек. На фоне этих кризисных событий разворачивается так называемая кембрийская «скелетная революция», сформировавшая основные черты того мира, в котором мы живем.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава 8 Моя собственная «водяная гипотеза». «X-pithecus». «Водяные дети» Чарковского. «Грудная гипотеза» Морган и моя. Сексуальные гипотезы Морриса. Прямохождение. Краткий обзор развития икспитека.
Глава 8 Моя собственная «водяная гипотеза». «X-pithecus». «Водяные дети» Чарковского. «Грудная гипотеза» Морган и моя. Сексуальные гипотезы Морриса. Прямохождение. Краткий обзор развития икспитека. Десмонд Моррис – зоолог и директор Лондонского зоопарка. Кстати, после
Глава 13 В чем причина оледенений? Моя гипотеза. Тотальное оледенение. Данные изучения Марса.
Глава 13 В чем причина оледенений? Моя гипотеза. Тотальное оледенение. Данные изучения Марса. «В чем причина оледенений?» – спрашивает Бьёрн Куртен в своей книге «Ледниковая эпоха».Несмотря на все усилия науки, ответить на этот вопрос чрезвычайно трудно. Изложив
Резюме: первые шаги на пути к многоклеточности
Резюме: первые шаги на пути к многоклеточности Вернемся, однако, к нашей основной теме «Бегство от одиночества» и посмотрим, чем же замечательны хищные нитчатые бактерии в рамках эволюционного сценария, который позволил бы реконструировать начальные этапы перехода от
Шаг за шагом по пути к многоклеточности
Шаг за шагом по пути к многоклеточности Замечательную иллюстрацию того, как именно мог идти эволюционный процесс перехода от «бессвязной однородности к сплоченной разнородности» у протистов-эукариот, дают нам так называемые растительные жгутиконосцы (рис. 3.2). Ботаники
Глава II. Возникновение нервных клеток и мозга
Глава II. Возникновение нервных клеток и мозга Причиной возникновения нервной системы стала низкая скорость получения информации о внешнем и внутреннем мире организма с донервной организацией. Его ткани состояли из клеток со сходной химической, электромагнитной и
ГЛАВА 5 Ранний докембрий: древнейшие следы жизни на Земле. Маты и строматолиты. Прокариотный мир и возникновение эукариотности
ГЛАВА 5 Ранний докембрий: древнейшие следы жизни на Земле. Маты и строматолиты. Прокариотный мир и возникновение эукариотности В «Происхождении видов» Ч. Дарвин честно и четко сформулировал вопросы, на которые его теория не давала (при тогдашнем уровне знаний)
ГЛАВА 9 Поздний палеозой — ранний мезозой: криоэры и термоэры. Палеозойские леса и континентальные водоемы: растения и насекомые
ГЛАВА 9 Поздний палеозой — ранний мезозой: криоэры и термоэры. Палеозойские леса и континентальные водоемы: растения и насекомые Выдающийся палеоботаник С. В. Мейен, анализируя механизм возникновения прочно устоявшихся научных предрассудков, приводит в качестве
ГЛАВА 10 Поздний палеозой — ранний мезозой: эволюция наземных позвоночных (1). Анамнии и амниоты. Две линии амниот — тероморфная и завроморфная
ГЛАВА 10 Поздний палеозой — ранний мезозой: эволюция наземных позвоночных (1). Анамнии и амниоты. Две линии амниот — тероморфная и завроморфная Мы расстались с самыми первыми тетраподами (девонской ихтиостегой и родственными ей формами), когда те… Чуть было не сказал:
ГЛАВА 11 Поздний мезозой: эволюция наземных позвоночных (2). Завроморфный мир. Маммализация териодонтов. Динозавры и их вымирание
ГЛАВА 11 Поздний мезозой: эволюция наземных позвоночных (2). Завроморфный мир. Маммализация териодонтов. Динозавры и их вымирание Итак, мы знаем, что с самого момента появления истинно наземных позвоночных — амниот — произошло их разделение на две эволюционные ветви:
Глава 8 Неадаптивная нулевая гипотеза эволюции генома и истоки биологической сложности
Глава 8 Неадаптивная нулевая гипотеза эволюции генома и истоки биологической сложности Пер. А. НеизвестногоЭволюционная энтропия и сложностьНемногие модные слова в последние два десятилетия были настолько популярны и в то же время определялись столь разнообразно,
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы Пер. А. НеизвестногоВ предыдущей главе мы обсудили возможные сценарии возникновения клеток и (будем надеяться) достигли
Глава 1 ВОЗНИКНОВЕНИЕ, СОДЕРЖАНИЕ И МЕТОДЫ ФИЗИОЛОГИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 1 ВОЗНИКНОВЕНИЕ, СОДЕРЖАНИЕ И МЕТОДЫ ФИЗИОЛОГИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ С незапамятных времен внимание человека привлекали проявления психической жизни. Откуда берутся мысли? Как понимать сознание, вмещающее весь мир и управляющее нашими поступками? Что
Глава 2. Возникновение жизни: самозарождение и панспермия
Глава 2. Возникновение жизни: самозарождение и панспермия Трудно создать хорошую теорию, теория должна быть разумной, а факты не всегда таковы. Джордж У. Бидл, генетик, лауреат Нобелевской премии 1958 г. в области физиологии и медицины Физик Филипп Моррисон как-то заметил,
Глава 10. Эволюция психики и возникновение культуры
Глава 10. Эволюция психики и возникновение культуры Остается загадкой, почему Homo sapiens, возникший около 130 тысяч лет назад, в течение многих десятков тысячелетий почти не прогрессировал в области культуры. Но в районе 30–35 тысяч лет назад произошел резкий скачок. Чем это
Глава первая Эволюция Возникновение сложности
Глава первая Эволюция Возникновение сложности Без света эволюции ничто в биологии не имеет смысла. Феодосий Добжанский Великая идея: эволюция идет путем естественного отбораЖизнь столь совершенна, что, как долгое время считали, ее было необходимо сотворить особо. Ибо
Докембрий
Докембрий Докембрий — древнейшая геологическая и палеонтологическая система. Составляют ее различные эры истории Земли. Докембрий включает 6/7 всего времени жизни планеты: от самых старых известных нам геологических пород до начала палеозоя и первого его периода —