Перезапуск эволюции
Перезапуск эволюции
Американский теоретический биолог Стюарт Кауфман (Stuart Kauffman) в статье 1985 года хорошо выразил суть вопроса:
Один способ подчеркнуть наше нынешнее невежество состоит в том, чтобы поинтересоваться, если бы эволюция должна была повториться от докембрия, когда ранние эукариотические клетки были уже сформированы, на что были бы похожи организмы через один или два миллиарда лет. И, если бы эксперимент был повторен несметное число раз, какие свойства организмов неоднократно возникали бы, а какие были бы редкими, какие свойства складывались бы в эволюции легко, а какие с трудом? Основной недостаток нашего нынешнего рассуждения об эволюции состоит в том, что она не заставляет нас ставить такие вопросы, хотя ответы могли бы, по сути, привести к глубокому пониманию ожидаемых признаков организмов.
Мне особенно нравится статистическое условие Кауфмана. Он предусматривает не только один мысленный эксперимент, но и статистическую совокупность мысленных экспериментов в поисках общих законов жизни, в отличие от частных проявлений отдельных жизней. Вопрос Кауфмана близок к научно-фантастическому вопросу о том, на что могла бы быть похожа жизнь на других планетах – хотя на других планетах стартовые и действующие условия будут иными. На большой планете сила тяжести наложила бы целую новую систему давлений отбора. Животные размером с пауков не могли бы иметь тонких конечностей (они сломались бы под весом), а нуждались бы в поддержке крепких, похожих на стволы деревьев вертикальных столбов, на которых стоят наши слоны. И наоборот, на меньшей планете животные размером со слонов, но легкого телосложения, могли бы носиться и прыгать по поверхности, как прыгающие пауки. Эти предсказания относительно строения тела будут применимы ко всей статистической совокупности миров с высокой силой тяжести и ко всей статистической совокупности миров с малой силой тяжести.
Сила тяжести – изначально данное условие планеты, на которое не может влиять жизнь. Также ее расстояние до центральной звезды. Также скорость ее вращения, которая определяет длину дня. Также наклон ее оси, который на такой планете как наша, с его почти круглой орбитой, является главным определяющим фактором сезонов. На планете с орбитой, далекой от круговой, такой как Плутон, кардинальное изменение расстояния до центральной звезды было бы намного более существенным фактором, определяющим сезонность. Наличие, расстояние, масса и орбита луны или лун оказывает тонкое, но сильное влияние на жизнь через приливы и отливы. Все эти факторы – исходные данные, не находящиеся под влиянием жизни, и поэтому рассматриваются как постоянные в последовательных повторениях мысленного эксперимента Кауфмана.
Прежние поколения ученых рассматривали бы погоду и химический состав атмосферы также как исходные данные. Теперь мы знаем, что атмосфера, особенно высокое содержание в ней кислорода и низкое содержание углерода, обусловлена жизнью. Таким образом, наш мысленный эксперимент должен учесть возможность, что в последовательных повторениях эволюции атмосфера могла бы измениться под влиянием каких-либо форм развивающейся жизни. Жизнь могла таким образом влиять на погоду и даже на большие климатические происшествия, такие как ледниковые периоды и засухи (Мой покойный коллега У. Д. Гамильтон, который был прав слишком много раз, чтобы его осмеивать, предположил, что облака и дождь – адаптация, произведенная микроорганизмами для своего собственного рассеивания.).
Насколько мы знаем, самые глубинные области Земли остаются незатронутыми жизненной пеной с ее поверхности. Но мысленные эксперименты по перезапуску эволюции должны признать возможные различия в ходе тектонических событий, и, следовательно, в истории расположения континентов. Интересный вопрос, должны ли быть представлены события вулканизма, землетрясений и бомбардировок из космоса, чтобы получить одинаковые последовательные перезапуски Кауфмана. Вероятно, разумно рассматривать тектонику и астрономические столкновения как важные переменные, которые могут быть усреднены, если мы представим себе достаточно большую статистическую совокупность перезапусков.
Как мы подойдем к ответу на вопрос Кауфмана? На что была бы похожа жизнь, если ее повторно «прокрутить» статистическое количество раз? Сразу же мы можем признать целый ряд вопросов Кауфмана постоянно увеличивающейся трудности. Кауфман хотел перевести часы на момент, когда эукариотическая клетка была собрана из своих бактериальных компонентов. Но мы могли бы предложить перезапустить процесс на две или три эры раньше, от самого происхождения жизни. Или, в другой крайности, мы можем перезапустить часы намного позже, скажем, около Копредка 1, нашего раскола с шимпанзе, и поинтересоваться, представлены ли гоминиды в статистически значимых количествах в перезапусках, чтобы жизнь достигла Копредка 1, развилось прямохождение, увеличился мозг, возник язык, цивилизация и бейсбол. Попутно есть вопросы Кауфмана о происхождении млекопитающих, о происхождении позвоночных животных и много других вопросов Кауфмана.
Кроме чистых предположений, представляет ли история жизни, произошедшей на самом деле, нечто похожее на естественный эксперимент Кауфмана, который может послужить нам ориентиром? Да, представляет. Мы ознакомились с несколькими естественными экспериментами в течение нашего странствия. Удачные случаи длительной географической изоляции Австралии, Новой Зеландии, Мадагаскара, Южной Америки, даже Африки дают нам приблизительный перезапуск главных эпизодов эволюции.
Эти массивы суши были изолированы друг от друга и от остальной части мира во время значительного периода после того, как исчезли динозавры, когда группа млекопитающих показала большую часть своего эволюционного творческого потенциала. Изоляция не была полной, но была достаточной, чтобы благоприятствовать развитию лемуров на Мадагаскаре и распространению древних и разнообразных афротериев в Африке. В случае Южной Америки мы выделили три отдельных момента возникновения млекопитающих с длительными периодами изоляции между ними. Австралинея представляет наиболее идеальные условия для подобного рода естественного эксперимента – ее изоляция была почти полной в течение большой части соответствующего периода, и он начался с очень маленького, возможно единственного инокулята сумчатых. Новая Зеландия – исключение, она одна среди этих показательных естественных экспериментов оказалась без млекопитающих в рассматриваемый период.
Глядя на эти естественные эксперименты, главным образом я впечатлен тем, какой схожей оказывается эволюция, если позволить запустить ее дважды. Мы видели, как похож тасманийский сумчатый волк на собаку, сумчатый крот на крота, летающий поссум на белок-летяг, сумчатый саблезубый тигр на саблезубого тигра (и на различных «ложных саблезубых» среди плацентарных хищников). Различия также поучительны. Кенгуру является прыгающей заменой антилопы. Прыжки на двух ногах, отточенные до совершенства эволюционным прогрессом, могут быть столь же внушительно быстрыми, как галопирование на четырех ногах. Но эти две манеры бега радикально отличаются друг от друга особенностями, которые вызвали основные изменения во всей анатомии. По-видимому, при некотором предшествующем разделении путей одна из этих двух «экспериментальных» линий, возможно, пошла путем совершенствования прыжков на двух ногах, а другая, вероятно, оттачивала галопирование на четырех. Оказалось – возможно, первоначально по почти случайным причинам – что кенгуру прыгали одним способом, а антилопы скакали другим. Мы теперь удивляемся нисходящему потоку различий между конечными продуктами.
Млекопитающие претерпели несоизмеримую эволюционную радиацию примерно одновременно друг с другом на различных массивах суши. Вакуум, оставленный динозаврами, дал им такую возможность. Но и у динозавров в свое время была подобная эволюционная радиация, хотя с известными пробелами – например, я не могу получить ответ на свой вопрос, почему, кажется, не было динозавров «кротов». И до динозавров были еще другие многочисленные параллели, особенно среди млекопитающеподобных рептилий, и они также достигали высшей точки в сходном диапазоне типов.
Когда я даю публичные лекции, я всегда пытаюсь ответить на вопросы в конце. Самый распространенный вопрос, безусловно: «Во что в будущем могли бы эволюционировать люди?» Мой собеседник всегда, похоже, трогательно предполагает, что это – новый и оригинальный вопрос, и каждый раз мое сердце сжимается. Потому что это – вопрос, от которого уклонится любой благоразумный эволюционист. Вы не можете подробно предсказать будущую эволюцию любых видов, кроме как сказать, что статистически подавляющее большинство видов вымерло. Но, хотя мы не можем предсказать будущее любого вида, скажем, на 20 миллионов лет вперед, мы можем предсказать общий диапазон экологических типов, которые будут обитать кругом. Будут травоядные животные и хищники, питающиеся травой и ветками, поедающие мясо, рыбу и насекомых. Сами эти диетические прогнозы предполагают, что через 20 миллионов лет все еще будет пища, соответствующая этим понятиям. Животные, питающиеся веточным кормом, предполагают длительное существование деревьев. Насекомоядные предполагают насекомых, или каких-либо маленьких, длинноногих беспозвоночных – какашек (doodoos), если употреблять этот полезный технический термин из Африки. В пределах каждой категории, травоядных, плотоядных и так далее, будет диапазон размеров. Будут бегающие, летающие, плавающие, ползающие и роющие животные. Виды не будут точно такими же, какие мы видим сегодня, или параллелями, которые эволюционировали в Австралии или Южной Америке, или аналогами динозавров, или подобными млекопитающим аналогами рептилий. Но будет похожий диапазон типов, ведущих похожий диапазон образов жизни.
Если в течение следующих 20 миллионов лет будет масштабная катастрофа и массовое исчезновение, сопоставимое с концом динозавров, мы можем ожидать, что диапазон экотипов будет получен из новых наследственных отправных точек, и – несмотря на мое предположение о грызунах на Свидании 10 – было бы весьма трудно предположить, кто из сегодняшних животных обеспечит эти отправные точки. Викторианская карикатура ниже показывает ихтиозавра-профессора, рассуждающего о человеческом черепе из некоторого отдаленного повторяющегося прошлого. Оговорка, во времена динозавров. Ихтиозавр-профессор обсуждал свой катастрофический конец, и ему было бы весьма трудно предсказать, что их место было бы занято потомками млекопитающих, которые были тогда маленькими, ничтожными ночными насекомоядными.

Ихтиозавр-профессор. Викторианская карикатура «Ужасные изменения» Генри Де ла Бича (Henry de la Beche), высмеивающая взгляды, высказанные Чарльзом Лайелем в «Основах геологии». Лайель предположил, что периодические изменения в климате Земли и их связь с дикой природой могли бы в будущем привести к миру, в котором игуанодон будет снова бродить по лесам, а ихтиозавры вновь появятся в морях.
По общему признанию, все это касается весьма недавней эволюции, не столь продолжительной, как перезапуск, представляемый Кауфманом. Но эти недавние перезапуски могут, конечно, преподать нам некоторые уроки о характерной для эволюции сходимости ее результатов. Если ранняя эволюция двигалась в том же направлении, что и более поздняя, то их уроки могли бы быть сведены к общим принципам. Мое предположение, что принципы, которые мы обнаруживаем в недавней эволюции, начиная со смерти динозавров, вероятно, справедливы, по крайней мере, до кембрийского периода, и, вероятно, до происхождения эукариотической клетки. У меня есть догадка, что параллелизм радиации млекопитающих в Австралии, Мадагаскаре, Южной Америке, Африке и Азии может обеспечить своего рода шаблон для того, чтобы ответить на вопросы Кауфмана относительно намного более ранних отправных точек, таких как та, которую он выбрал – происхождение эукариотической клетки. Ранее этого знакового события уверенность испаряется. Мой коллега Марк Ридли (Mark Ridley) в «Демоне Mенделя» подозревает, что происхождение сложности эукариот было чрезвычайно невероятным случаем, возможно, даже более невероятным, чем само происхождение жизни. Под влиянием Ридли я бьюсь об заклад, что большинство мысленных экспериментов перезапусков, которые начинаются с происхождения жизни, не приведут к эукариократии.
Мы не должны полагаться на географическое разделение, как в австралийском естественном эксперименте, чтобы изучить конвергенцию. Мы можем представить себе эксперимент с эволюцией, перезапускаемой не из одной и той же отправной точки в различных географических областях, а из различных отправных точек – весьма вероятно, в одной и той же географической области: конвергенцию животных, столь несвязанных друг с другом, что то, о чем она говорит нам, не имеет никакого отношения к географическому разделению. Считалось, что «глаз» развивался независимо от 40 до 60 раз повсюду в животном мире. Это вдохновило мою главу, названную «В сорок раз более длинный путь к прозрению» в «Восхождении на пик Невероятности», таким образом, я не буду повторяться здесь, лишь скажу, что профессор Майкл Лэнд из университета Суксесса, наш ведущий эксперт по сравнительной зоологии глаз, насчитывает девять независимых принципов оптического механизма, каждый из которых развивался не раз. Он был достаточно любезен, чтобы подготовить для этой книги пейзаж, перепечатанный напротив, в котором отдельные пики представляют независимые развития глаз.
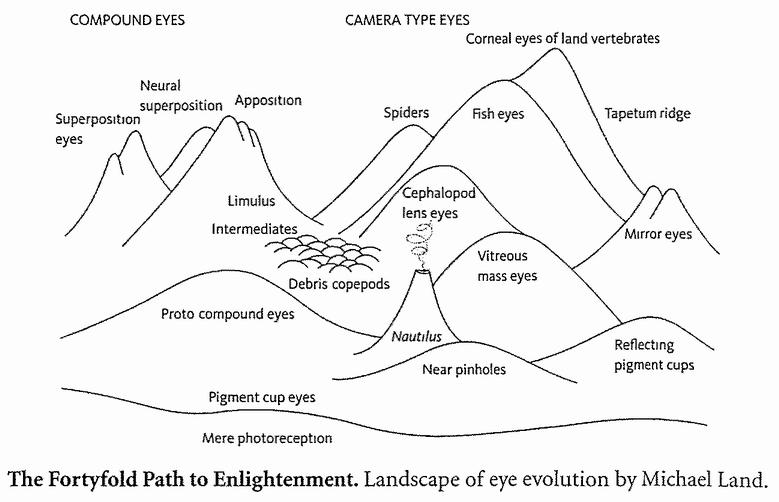
Похоже, что жизнь, по крайней мере, как мы знаем ее на этой планете, почти до неприличия стремится развить глаза. Мы можем уверенно предсказать, что статистическая модель перезапусков Кауфмана достигла бы наивысшей точки в глазах. И не просто в глазах, но и в сложных глазах, подобных глазам насекомых, креветок или трилобитов, и линзовых глазах, как у нас или кальмаров, с цветным видением и механизмами для того, чтобы точно настроить фокус и апертуру. Также, весьма вероятно, в параболических глазах-рефлекторах, как у блюдец, и глазах-обскурах, как у наутилуса, современного, похожего на аммонита моллюска с его плавучей спиральной раковиной, которого мы встретили на Свидании 26. И если есть жизнь на других планетах во Вселенной, бьюсь об заклад, что там также будут глаза, основанные на тех же оптических принципах, которые мы знаем на нашей планете. Есть очень много способов сделать глаз, и жизнь, как мы знаем, весьма вероятно, обнаружила их все.
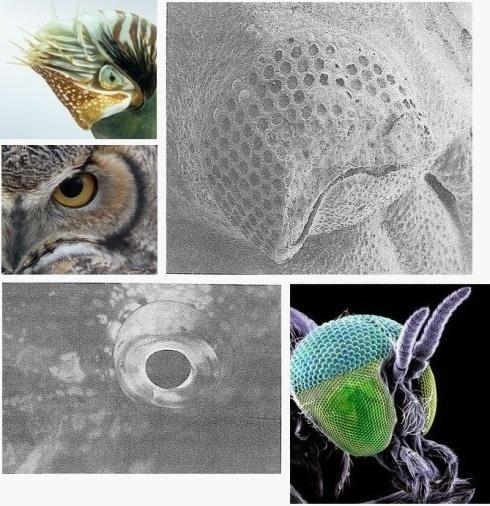
Почти неприлично ревностное стремление развить глаза. Некоторые примеры глаз. По часовой стрелке, сверху слева: Nautilus pompilius (глаз-обскура); ископаемый трилобит (Phacops, сложный глаз, сделанный из кальцитовых линз – некоторые из них могут быть видны на своих местах в верхней части глаза); мошка simulian (Simulium damnosum, сложный глаз); рыба-попугай (Sparisoma viride, рыбий глаз); большой американский пугач (Bubo virgimanus, роговичный глаз).
Мы можем подвести подобного рода итог для другой адаптации. Эхолокация – прием, который заключается в испускании звукового импульса и навигации благодаря точному расчету времени эха – была развита, по крайней мере, четыре раза: у летучих мышей, зубатых китов, гуахаро и пещерных стрижей-саланганов. Не так много раз, как глаза, но все же достаточно часто, чтобы заставить нас считать не слишком маловероятным, что при благоприятных условиях она разовьется. Весьма вероятно, также, что перезапуски эволюции открыли бы вновь одни и те же характерные принципы: одни и те же приемы для того, чтобы противостоять трудностям. Снова же, я не буду повторять свои объяснения из предыдущей книги (В данном случае «Слепой часовщик».), но просто подведу итог того, что мы могли бы предсказать для перезапусков эволюции. Эхолокация должна неоднократно развиваться, используя очень высокочастотные крики (для лучшего обнаружения деталей, чем низкочастотные). Крики, по крайней мере, у некоторых видов, вероятно, будут частотно-модулируемыми, резко понижающимися или повышающимися по тону в течение каждого крика (точность повысилась, потому что начало каждого эха отличается от заключительной части своим тоном). Вычислительный аппарат, используемый, чтобы проанализировать эхо, будет делать очень хорошие (подсознательные) расчеты, основанные на доплеровских изменениях в частоте эха, поскольку эффект Доплера, конечно, повсеместно присутствует на любой планете, где есть звук, и летучие мыши изощренно его применяют.
Откуда мы знаем, что такие вещи как глаз или эхолокация развивались независимо? Глядя на генеалогическое дерево. Родственники гуахаро и пещерных стрижей-саланганов не используют эхолокацию. Гуахаро и пещерные стрижи-саланганы по отдельности освоили жизнь в пещерах. Мы знаем, что они развили эту технику независимо от летучих мышей и китов, так как никто другой в окружающем генеалогическом дереве этого не делает. Различные группы летучих мышей, возможно, независимо многократно развивали эхолокацию. Мы не знаем, сколько раз еще развивалась эхолокация. Некоторые землеройки и тюлени обладают элементарной формой этого искусства (а также некоторые слепые люди). Обладали ли им птеродактили? С тех пор хорошие средства к существованию приносили ночные полеты, и так как летучих мышей в те дни вокруг не было, это весьма возможно. То же самое справедливо для ихтиозавров. Они выглядели в точности как дельфины и, по-видимому, вели схожий образ жизни. Так как дельфины интенсивно используют эхолокацию, разумно задаться вопросом, применяли ли ее также ихтиозавры во времена, предшествующие дельфинам. Нет никаких прямых свидетельств, и мы должны оставаться непредубежденными. Один довод против: у ихтиозавров были необычно большие глаза – одна из их самых заметных особенностей – и это может подсказывать, что они полагались на зрение вместо эхолокации. У дельфинов глаза относительно маленькие, и одна из их самых заметных особенностей, округлая шишка или «дыня» выше клюва, действует как акустическая «линза», сосредотачивая звук в узкий луч, фокусируя его перед животным, как прожектор.
Как любой зоолог, я могу рассматривать свою ментальную базу данных животного мира и придумать предполагаемый ответ на вопросы следующего рода: «Сколько раз X был независимо развит?» Можно было бы сделать хорошую научно-исследовательскую работу, сделав подсчет более систематичным. По-видимому, некоторые Xs дадут ответ, что возникли «много раз», как в случае с глазами, или «несколько раз», как в случае с эхолокацией. Другие «только однажды» или даже «никогда», хотя я должен сказать, что удивительно трудно найти их примеры. И различия могли быть интересными. Я подозреваю, что мы обнаружили бы определенные потенциальные эволюционные тропы, по которым «стремится» следовать жизнь. Другие тропы имеют большее «сопротивление». В «Восхождении на пик Невероятности» я развивал аналогию огромного музея всей жизни, и реальной и мыслимой, с коридорами, уходящими во многих измерениях и представляющими эволюционные изменения, снова же, и реальные и мыслимые. Некоторые из этих коридоров были широко открытыми, почти манящими. Другие преграждались барьерами, которые было трудно, или даже невозможно, преодолеть. Эволюция неоднократно проносится по легким коридорам и только иногда, и неожиданно, перепрыгивает один из трудных барьеров. Я возвращусь к идее стремления и «нежелания» эволюционировать, когда буду обсуждать «эволюцию самой эволюции».
Давайте теперь быстро пройдемся еще по некоторым примерам, где стоило бы делать систематический подсчет того, сколько раз развился X. Ядовитое жало (вводящее яд подкожно через остроконечную трубку) развилось, по крайней мере, десять раз независимо: у медуз и их родственников, у пауков, скорпионов, многоножек, насекомых (У пчел, ос и муравьев жало – измененная яйцекладущая трубка, и жалят только самки.), моллюсков (конусы), змей, группы акул (скаты), костистых рыб (бородавчатка), млекопитающих (самец утконоса) и растений (крапива двудомная). Держу пари, что яд, включая подкожную инъекцию, развился бы в перезапусках.
Издавание звука в социальных целях развилось независимо у птиц, млекопитающих, сверчков и кузнечиков, цикад, рыб и лягушек. Электролокация, использование слабых электрических полей для навигации, развилось несколько раз, как мы видели в «Рассказе Утконоса». А так же – вероятно, в дальнейшем – и использование электрических токов в качестве оружия. Физика электричества одинакова во всех мирах, и мы с некоторой уверенностью могли бы сделать ставку на повторное развитие существ, которые используют электричество и в навигационных и агрессивных целях.
Истинный маховый полет, в противоположность пассивному планированию или парашютированию, кажется, развился четыре раза: у насекомых, птерозавров, летучих мышей и птиц. Парашютирование и планирование различных видов развились множество раз, возможно сотни раз независимо, и они вполне могут быть эволюционными предшественниками настоящего полета. Примерами служат ящерицы, лягушки, змеи, «летающие» рыбы, кальмары, шестокрылы, сумчатые и грызуны (дважды). Я бы поставил много денег на планеристов, появляющихся в перезапусках Кауфмана, и разумную сумму на истинных маховых летунов.
Реактивное движение, возможно, развилось дважды. Головоногие моллюски делают это, в случае с кальмарами – с высокой скоростью. Другим примером, который я могу вспомнить, является также моллюск, но не быстрый. Гребешки в основном живут на морском дне, но иногда они плавают. Они ритмично открывают и закрывают свои две раковины, как пару щелкающих кастаньет. Вы могли бы подумать (я тоже), что это толкало бы их «назад», в направлении, противоположном щелканию. Фактически они двигаются «вперед», как если бы прогрызали себе путь в воде. Как это может быть? Ответ заключается в том, что щелкающие движения прокачивают воду через пару отверстий позади замка раковины. Эти два реактивных двигателя толкают животное «вперед». Эффект настолько алогичный, что является почти смешным.
Как насчет вещей, которые развились только однажды или ни разу? Как мы узнали из «Рассказа Ризобия», колесо, в истинным, свободно вращающемся значении, кажется, развилось только однажды, у бактерий, прежде чем было, наконец, изобретено в технологии людей. Язык также, очевидно, развился только у нас: то есть, по крайней мере, в 40 раз реже, чем глаза. Удивительно трудно размышлять о «хороших идеях», которые развились только однажды.
Я задал сложную задачу своему Оксфордскому коллеге, энтомологу и натуралисту Джорджу МакГэвину (George McGavin), и он придумал хороший список, но все еще короткий по сравнению со списком вещей, которые развились много раз. Жуки бомбардиры рода Brachinus по опыту д-ра МакГэвина уникальны в смешивании химикатов для взрыва. Компоненты создаются и содержатся в отдельных (разумеется!) железах. Когда угрожает опасность, они впрыскиваются в камеру в задней части жука, где взрываются, выпуская отравляющую (едкую и кипящую) жидкость через направленную форсунку во врага. Случай известен креационистам, которые его любят. Они думают, что явно невозможно развиваться постепенными этапами, потому что промежуточные стадии все взорвались бы (Я любил демонстрировать ошибочность этого аргумента во время моих Королевских рождественских лекций для детей, показанных по телевидению Би-би-си в 1991 году. Примеряя шлем времен Второй Мировой войны, и предлагая возбужденным членам аудитории отойти, я смешивал гидрохинон и перекись водорода, два взрывчатых компонента бомбардира. Ничего не происходило. Они даже не нагревались. Взрыв требует катализатора. Я поднимал концентрацию катализатора постепенно, чем неуклонно увеличивал опасный свист до убедительной кульминации. В природе жук обеспечивает катализатор, и не имел бы никаких препятствий для постепенного и благополучного увеличения дозы за время эволюции.).
Следующая в списке в МакГэвина – рыба-стрелок семейства Toxotidae, которая может быть уникальной в стрельбе, сбивая добычу на расстоянии. Она подплывает к поверхности и плюет набранной в рот водой в сидящего насекомого, сбивает его в воду, где и поедает. Другим возможным кандидатом на роль «сбивающего» хищника мог бы быть муравьиный лев. Муравьиные львы – личинки насекомого отряда Neuroptera. Как и многие личинки, они не похожи на взрослых. Со своими огромными челюстями они были бы хорошим кандидатами на роль в фильме ужасов. Каждый муравьиный лев скрывается в песке чуть ниже поверхности в основании конической ямы-ловушки, которую он роет самостоятельно. Он роет, энергично откидывая песок наружу от центра – это вызывает маленькие оползни краев ямы, и законы физики делают остальное, аккуратно формируя конус. Добыча, обычно муравьи, падает в яму и скатывается с крутых краев в челюсти муравьиного льва. Возможное подобие рыбе-стрелку заключается в том, что добыча падает не только пассивно. Она иногда сбивается в яму крупинками песка. Песчинки, однако, не столь нацелены, как плевки рыбы-стрелка, наводимые с сокрушительной точностью сфокусированными бинокулярными глазами.
Шипящие пауки семейства Scytodidae тоже немного необычны. Не имея быстроты пауков-волков и не ставя сети, шипящий паук выбрасывает с некоторого расстояния на свою добычу ядовитый клей, прижимая ее к земле, пока не придет и не искусает ее до смерти. Это отличается от техники сбивания добычи рыбы-стрелка. Различные животные, например плюющиеся ядом кобры, плюются для защиты, а не для того, чтобы поймать добычу. Пауки боладоры, Mastophora, также необычны, и являются, вероятно, другим уникальным случаем. Они, можно сказать, бросают в добычу снаряд (мотыльков привлекает фальшивый половой аромат самок, который синтезирует паук). Но снаряд, маленький шарик шелка, присоединен к шелковой нити, которую паук вращает подобно лассо (или бола) и наматывает (Хамелеоны, можно сказать, плюют в добычу снарядами. Снаряд – значительное утолщение на конце языка, и (намного более тонкий) остальной язык немного похож на веревку, которой возвращается гарпун. В техническом отношении, кончик языка является баллистическим, что означает, что его свободно мечут, в отличие от кончика Вашего языка. Впрочем, хамелеоны не уникальны в этом отношении. Некоторые саламандры также баллистически метают кончик языка в добычу, и их снаряд (но не хамелеона) содержит в себе часть скелета. Он запускается как косточка арбуза, сжатая между Вашими пальцами.).
Прекрасен следующий кандидат МакГэвина на эволюционную уникальность. Это – водяной паук, Argyroneta aquatica. Эти пауки живут и охотятся полностью под водой, но, как дельфины, дюгони, черепахи, пресноводные улитки и другие наземные животные, которые возвратились в воду, им нужно вдыхать воздух. В отличие от всех других изгнанников, Argyroneta, однако, строит свой собственный водолазный колокол. Он прядет его из шелка (шелк – универсальное решение любой проблемы паука), прикрепляя к подводному растению. Паук идет на поверхность, чтобы набрать воздуха, который он носит таким же образом, как некоторые водяные клопы, в оболочке, окруженной волосами тела. Но в отличие от клопов, которые только носят везде за собой воздух, подобно баллону акваланга, паук носит воздух в своем водолазном колоколе и избавляется от него, чтобы пополнить запас. Паук сидит в этом водолазном колоколе, высматривая добычу, и там он ее хранит и поедает.
Но победителем среди примеров уникальности Джорджа МакГэвина является личинка африканского слепня по имени Tabanus. Как и ожидалось, в Африке лужи воды, в которой личинки живут и питаются, в конечном счете, высыхают. Каждая личинка зарывается в грязь и окукливается. Взрослая муха появляется из затвердевшей грязи и летит, питаясь кровью, чтобы, в конечном счете, закончить цикл, откладывая яйца в лужах воды, когда возвращаются дожди. Зарытая личинка уязвима для предсказуемой угрозы. Когда грязь подсыхает, она раскалывается, и есть риск, что трещина пройдет прямо через убежище личинки. Теоретически она имела бы возможность спастись, если бы могла так или иначе создать путь для любой трещины, которая приближается к ней, пуская ее вместо этого в обход. И личинка действительно добивается этого поистине замечательным и, вероятно, уникальным способом. Прежде чем зарыться в своей собственной камере окукливания, она сначала протискивается в грязь вниз по спирали. Затем она протискивается назад на поверхность по противоположной спирали. Наконец она ныряет в грязь прямо вниз по центру между этими двумя спиралями, и это – ее место отдыха в тяжелые времена до возвращения воды. Теперь Вы видите, что это означает? Личинка заключена в грязевый цилиндр, круговая граница которого была ослаблена заранее предварительным рытьем по спирали. Это означает, что, когда трещина вьется через сохнущую грязь, если она поражает край цилиндрического столбика, вместо того, чтобы прорезаться прямо через середину, она идет криволинейным обходным путем вокруг края цилиндра, и личинка спасена. Это похоже на перфорацию вокруг почтовой марки, которая останавливает разрывы, идущие через марку. Доктор МакГэвин полагает, что этот изобретательный прием буквально уникален для данного рода слепней.
Но существуют ли какие-нибудь хорошие идеи, которые никогда не развились под влиянием естественного отбора? Насколько я знаю, ни одно животное на этой планете никогда не развивало орган для передачи или приема радиоволн для дальней коммуникации. Другим примером является использование огня. Человеческий опыт показывает, насколько полезным он может быть. Есть некоторые растения, чьим семенам нужен огонь, чтобы прорасти, но я не думаю, что они используют его так же, как, например, электрические угри используют электричество. Использование металла в целях формирования скелета – другой пример хорошей идеи, которая никогда не развивалась, за исключением человеческих артефактов. По-видимому, ее трудно осуществить без огня.
Подобного рода сравнительные упражнения, подсчитывающие, что развивается часто, а что редко, когда мы делаем их наряду с географическими сравнениями, обсужденными ранее, могли бы позволить нам предсказать обстоятельства жизни вне этой планеты, так же как предположить вероятный результат мысленного эксперимента по перезапускам эволюции в стиле Кауфмана. Мы, безусловно, ожидаем глаза, уши, крылья и электрические органы, но, возможно, не взрывы жука бомбардира или водяные пули рыбы-стрелка.
Те биологи, которые, можно сказать, взяли на себя инициативу покойного Стивена Джея Гулда, расценивают всю эволюцию, включая послекембрийскую, как полностью случайную – удачу, которая вряд ли может быть повторена в перезапусках Кауфмана. Назвав это «перемоткой ленты эволюции», Гулд независимо развил мысленный эксперимент Кауфмана. Вероятность возникновения чего-либо отдаленно похожего на человека при втором запуске широко признана как исчезающе маленькая, и Гулд это убедительно выразил в «Замечательной жизни». Именно эта ортодоксальность привела меня к обдуманному самоотречению в моей вступительной главе; фактически заставила меня предпринять путешествие в прошлое и теперь вынуждает оставить своих странствующих компаньонов в Кентербери и вернуться в одиночестве. И все же... Я долго задавался вопросом, не могла ли грозная ортодоксия случайности зайти слишком далеко. Мой обзор «Полного дома» Гулда (перепечатанный в «Служителе дьявола») защищал непопулярную идею прогресса в эволюции: не прогресса в сторону человека – Дарвин упаси! – но прогресса в направлениях, которые, по крайней мере, достаточно предсказуемы, чтобы подтвердить эти слова. Как я сейчас докажу, кумулятивное наращивание сложной адаптации, такой как глаза, убедительно указывает на версию прогресса – особенно, когда связано в воображении с некоторыми из замечательных продуктов конвергентной эволюции.
Конвергентная эволюция также вдохновила кембриджского геолога Саймона Конвея Морриса (Simon Conway Morris), чья дерзкая книга «Объяснение жизни: неотвратимые люди в одинокой Вселенной» представляет аргументы, совершенно противоположные «случайности» Гулда. Конвей Моррис имеет в виду, что его заглавие имеет значение, которое недалеко от буквального. Он действительно думает, что перезапуск эволюции привел бы ко второму пришествию человека: или кого-то, чрезвычайно близкого к человеку. И для такого непопулярного тезиса он приводит вызывающе храбрые доказательства. Два свидетеля, которых он неоднократно вызывает, это конвергенция и ограничение.
Конвергенцию мы встречали снова и снова в этой книге, включая эту главу. Схожие проблемы вызывают схожие решения, не только дважды или трижды, но и во многих случаях, множество раз. Я думал, что был довольно экстремальным в своем энтузиазме по поводу конвергентной эволюции, но я встретил достойного соперника в лице Конвея Морриса, который приводит ошеломляющее множество примеров, многие из которых я не встречал прежде. Но если я обычно объясняю конвергенцию, прибегая к сходному давлению отбора, Конвей Моррис добавляет доказательство своего второго свидетеля, ограничения. Материалы жизни и процессы эмбрионального развития допускают только ограниченный диапазон решений отдельной проблемы. Учитывая любую специфическую эволюционную стартовую ситуацию, существует лишь ограниченное число готовых решений. Так, если два эксперимента перезапуска Кауфмана сталкиваются с чем-то вроде схожих давлений отбора, ограничения, связанные с развитием, усиливают тенденцию к поиску аналогичных решений.
Вы можете видеть, как квалифицированный адвокат мог использовать эти двух свидетелей в защиту смелой веры, что перезапуск эволюции, вероятно, будет, положительно сходиться на прямоходящем существе с большим мозгом, двумя умелыми руками, направленными вперед камерными глазами и другими человеческими особенностями. К сожалению, это случилось лишь однажды на этой планете, но я полагаю, что должен быть первый раз. Я, признаюсь, был впечатлен аналогичными доводами Конвея Морриса в пользу предсказуемости эволюции насекомых.
Среди определяющих признаков насекомых есть следующие: ясно сочлененный экзоскелет; сложные глаза; характерная шестиногая походка, при которой три из шести шагающих ног всегда находятся на земле и тем самым очерчивают треугольник (две ноги с одной стороны, одна с другой), который придает животному устойчивость; дыхательные трубки, известные как трахеи, которые обеспечивают приток кислорода к внутренностям животного через специальные отверстия (дыхальца) по бокам тела; и, чтобы закончить список эволюционных особенностей, повторное (11 раз независимое!) развитие высокоорганизованных общественных колоний, как у медоносных пчел. Все довольно необычно? Все уникально в большой лотерее жизни? Напротив, все конвергентно.
Конвей Моррис перебирает свой список, показывая, что каждая позиция не раз развивалась в различных частях животного мира, во многих случаях несколько раз, включая несколько раз независимо у самих насекомых. Если природа находит настолько легким развить составные части насекомости отдельно, не является невероятным, что вся совокупность должна развиться дважды. Я склоняюсь перед верой Конвея Морриса, что мы должны прекратить думать о конвергентной эволюции как о колоритной редкости, которая будет замечательной и восхитительной, когда мы ее обнаружим. Возможно, мы должны придти к тому, чтобы считать это нормой, исключения из которой являются поводом для удивления. Например, истинный синтаксический язык, кажется, уникален для одного вида, нашего собственного. Возможно – и я возвращусь к этому – это единственное, чего недоставало бы повторно развившемуся мозговитому прямоходящему?
В моей вступительной главе «Тщеславный взгляд в прошлое» я согласился с предостережением против поиска примеров, рифм или причин в эволюции, но сказал, что буду осторожно флиртовать с ними. Глава «Возвращение Хозяина» дала возможность промчаться по всему течению эволюции в прямом направлении и понять, какие примеры мы можем обнаружить. Идея, что вся эволюция была нацелена на создание Homo sapiens, была, конечно, благополучно отвергнута, и ничто из того, что мы увидели в нашем путешествии в прошлое, ее не восстанавливает. Даже Конвей Моррис утверждает только, что нечто приблизительно похожее на нашу разновидность животного является одним из нескольких исходов – другим, например, являются насекомые – который мы ожидали бы увидеть при возвращении, если бы эволюция запускалась повторно снова и снова.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
5.2. Особенности эволюции
5.2. Особенности эволюции Понятие хаоса в противоположность понятию космоса было известно древним грекам. Синергетики называют хаотическими все системы, которые приводят к несводимому представлению в терминах вероятностей. Другими словами, такие системы нельзя описать
Начало эволюции
Начало эволюции Чрезвычайно приятно быть частью этого грандиозного эволюционного спектакля, даже если нам придется считать себя прямыми потомками тошнотворных газов и грозовых разрядов. X. Шепли От финиша к стартуМы рассмотрели проблему прогресса в живой природе.
Причины эволюции
Причины эволюции Прежде чем перейти к интересующему нас вопросу, попытаемся уяснить, как происходят эволюционные изменения. Труды Чарлза Дарвина окончательно привели ученых к признанию теории эволюции органического мира. Дарвин не только представил множество
Перезапуск эволюции
Перезапуск эволюции Американский теоретический биолог Стюарт Кауфман (Stuart Kauffman) в статье 1985 года хорошо выразил суть вопроса:Один способ подчеркнуть наше нынешнее невежество состоит в том, чтобы поинтересоваться, если бы эволюция должна была повториться от докембрия,
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции Пер. А. НадирянВ этой и следующей главах дается краткое описание современного состояния эволюционной биологии, какой она была до 1995 года, когда возникло новое направление науки — сравнительная геномика.
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции Пер. А. НестеровойВ этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись
Глава 9 Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции
Глава 9 Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции Пер. Д. ТулиноваДрама ламаркизмаКак уже отмечалось в предисловии к данной книге, одной из ключевых заслуг
НОМОГЕНЕТИКА ЭВОЛЮЦИИ
НОМОГЕНЕТИКА ЭВОЛЮЦИИ Номогенетическое направление в теории эволюции аналогично номотетическому в систематике. Для лучшего понимания полезно сказать несколько слов о классификации эволюционных теорий (Любищев, 1925). Поразительно, что многие серьезные биологи
5.1. «Эволюция эволюции»
5.1. «Эволюция эволюции» История эволюционизма, или, как иногда говорят, «эволюция эволюции», является темой огромного числа публикаций (Завадский К. М., Колчинский Э. И., 1977; Waddington С., 1975). Рассмотрим основные вехи этой истории.Эволюционизм возник как альтернатива учению о
Факторы эволюции
Факторы эволюции Процессы, изменяющие частоту аллелей в популяциях, получили название элементарных эволюционных факторов. В популяционной генетике выделяют четыре основных эволюционных фактора.Мутационный процесс. Мутации – единственный процесс образования новых
Скорости эволюции
Скорости эволюции До сих пор мы рассматривали скорости эволюции вообще, не пытаясь определить, какие изменения морфологии за 1 млн. лет соответствуют горотелическим или тахителическим скоростям. Однако из палеонтологической летописи можно извлечь конкретные
8. Факторы эволюции
8. Факторы эволюции Вспомните!Какие существуют причины изменения численности особей в популяции?В чём заключается роль мутаций в процессе эволюции?Наследственная изменчивость. Фактором, который обеспечивает возникновение нового генетического материала в популяции и