Глава 7 Происхождение эукариот: эндосимбиоз, удивительная история интронов и исключительная важность единичных событий в эволюции
Глава 7
Происхождение эукариот: эндосимбиоз, удивительная история интронов и исключительная важность единичных событий в эволюции
Пер. А. Грызлова
Организмы с большими, сложными клетками известны как эукариоты, что означает «обладающие настоящим ядром». Эти организмы включают три царства многоклеточных форм жизни: растения, бурые водоросли и животные, а также обширный набор одноклеточных форм, известных как протисты. Обычно эукариотические клетки крупнее прокариотических и обладают сложной внутриклеточной организацией с разнообразными ограниченными мембраной органеллами, включая ядро, давшее им имя, и митохондрии, произошедшие от эндосимбиотических бактерий. Таким образом, по любому разумному критерию, эукариотические клетки значительно более сложны, чем бактерии и археи. Возникновение такого уровня сложности — загадка эволюционной биологии, представляющаяся еще более интригующей вследствие особого, «личного» интереса. Ведь изучая происхождение эукариот, мы всматриваемся в исток нашего собственного происхождения.
Фундаментальные различия в клеточной организации между эукариотами и прокариотами являют собой, можно сказать, парадоксальное противопоставление особенностям организации генома, которых мы уже касались в главе 3. В то время как эукариотические клетки обладают гораздо более замысловатой, упорядоченной и сложной организацией, чем прокариотические клетки, геномы эукариот намного менее оптимизированы и содержат гораздо больше «мусора», чем геномы прокариот. Объяснение эволюционных основ этого явного парадокса представляет собой принципиальную проблему, и ее решение, вероятно, прольет свет также и на происхождение эукариот — или, точнее говоря, на происхождение клеточной организации эукариот. Эта проблема далека от решения и остается предметом жарких (иногда даже слишком жарких) споров. В данной главе мы рассмотрим эту загадку и, говоря шире, эволюционные связи между археями, бактериями и эукариотами настолько объективно и логично, насколько это возможно, чтобы понять, как могло возникнуть «эукариотическое состояние живого». В заключение этой дискуссии я надеюсь показать, что, хотя некоторые ключевые детали все еще требуют объяснения, контуры приемлемого специфического сценария возникновения эукариот становятся ясными, и этот сценарий согласуется по крайней мере с некоторыми необычными чертами геномов эукариот и удивительной сложностью эукариотической клеточной организации. Более того, в данной главе мы придем к выводу о том, что трехдоменная схема Вёзе не дает адекватной картины истории жизни, и обсудим некоторые общие следствия эволюционной истории эукариот, которые имеют непосредственное отношение к центральной теме этой книги — взаимодействию случайности и необходимости в эволюции.
Эукариотическая клетка, ее внутренняя архитектура и пропасть между прокариотической и эукариотической клеточной организацией
В этой книге нам нет никакой необходимости вдаваться в бесчисленные тонкие детали биологических структур. Однако для того, чтобы обсуждать происхождение эукариот (далее эукариогенез) предметно, мы должны полностью оценивать природу и глубину пропасти, разделяющей эукариотические и прокариотические клетки. Действительно, существует резкое различие по сложности организации клетки между эукариотами и прокариотами: типичная эукариотическая клетка примерно в тысячу раз больше по объему, чем обычная бактерия или архея, и обладает исключительно сложной внутриклеточной компартментализацией, которой нет даже у самых «продвинутых» прокариот. Ниже в этой главе мы кратко обсудим некоторые интересные исключения, такие как гигантские прокариотические клетки и прокариотические клетки, содержащие внутриклеточные компартменты. Тем не менее тщательное рассмотрение этих случаев подтверждает фундаментальную дихотомию эукариот и прокариот по клеточной организации.
Компартментализация эукариотических клеток основана на сложной, разносторонней системе внутренних мембран и актинтубулиновом цитоскелете. Удивительное следствие внутриклеточной компартментализации — это физическое отличие эукариотических клеток от прокариотических. У прокариот содержимое клетки является раствором, пусть и вязким, так что макромолекулы (белки и нуклеиновые кислоты) диффундируют более или менее свободно и достигают своих пунктов назначения в клетке в результате сочетания стохастических перемещений и переноса в составе специфических комплексов. Напротив, у эукариот макромолекулы не могут свободно диффундировать и вместо этого добираются до своего «места работы» при помощи сложных транспортных систем. Это различие ясно демонстрируется простым экспериментом, в котором мембраны прокариотических и эукариотических клеток искусственно делаются проницаемыми (пермеабилизуются): белки и нуклеиновые кислоты «вытекают» из пермеабилизованных бактериальных клеток, но, как правило, не из эукариотических (Hudder et al., 2003). Таким образом, цитозоль эукариотических клеток обладает значительно более низкой энтропией, чем у прокариот, — трудно придумать более фундаментальное различие.
Ядро — органелла, давшая название эукариотам, — содержит в себе геномную ДНК, организованную в виде хроматина и распределенную между несколькими хромосомами; это место транскрипции, сплайсинга и сборки рибосом. Само по себе ядро является частью системы внутренних мембран: оболочка ядра переходит в мембраны эндоплазматического ретикулума. Очевидно, что для функционирования эукариотической клетки ядро должно постоянно взаимодействовать с цитозолем. Действительно, оболочка ядра пронизана порами, чрезвычайно сложными структурами, отвечающими как за пассивный, так и за активный перенос всех видов молекул (и даже макромолекулярных комплексов, таких как субъединицы рибосом) в ядро и из него. Заметим, что ограничение расположения хроматина и транскрипции ядерным компартментом исключает сопряжение транскрипции и трансляции, отличительную черту экспрессии генов у прокариот. Ниже в этой главе мы рассмотрим фундаментальные последствия этого разобщения.
Изнутри ядро заполнено высокоструктурированным матриксом и в этом отношении походит на эукариотический цитозоль. Эукариотический хроматин, содержащийся в ядре, никоим образом нельзя считать просто молекулой ДНК, защищенной белками и регулярным образом упакованной в трехмерные структуры. Хроматин является чрезвычайно сложной динамической системой молекулярных машин, состоящих из множества специализированных белков, которые регулируют и координируют процессы репликации и экспрессии в основном посредством так называемого ремоделирования хроматина — модификации структуры хроматина, которая изменяет фактуру доступных участков (Clapier and Cairns, 2009). Хотя картина регуляции прокариотической экспрессии становится все сложнее и уже очень далека от простой схемы Жакоба—Моно (см. гл. 5), ничто в прокариотической клетке не может сравниться со сложностью эукариотического хроматина.
Качественные различия между эукариотами и прокариотами многочисленны и охватывают многообразные аспекты клеточной биологии, в частности те, которые имеют отношение к переработке информации, передаче сигналов и внутриклеточному переносу веществ (см. табл. 7-1). Сложность физической организации эукариотической клетки дополняется чрезвычайно изощренной сетью взаимодействующих сигнальных путей. Основные сигнальные системы эукариот — это киназно-фосфатазный механизм, регулирующий работу белков посредством фосфорилирования и дефосфорилирования; система убиквитина, которая управляет оборотом и локализацией белков посредством их обратимого убиквитинирования; регуляция трансляции посредством микроРНК; и регуляция транскрипции на уровне индивидуальных генов и ремоделирования хроматина.
Таблица 7-1
Краткое сравнение основных структурных и функциональных признаков эукариотических и прокариотических клеток
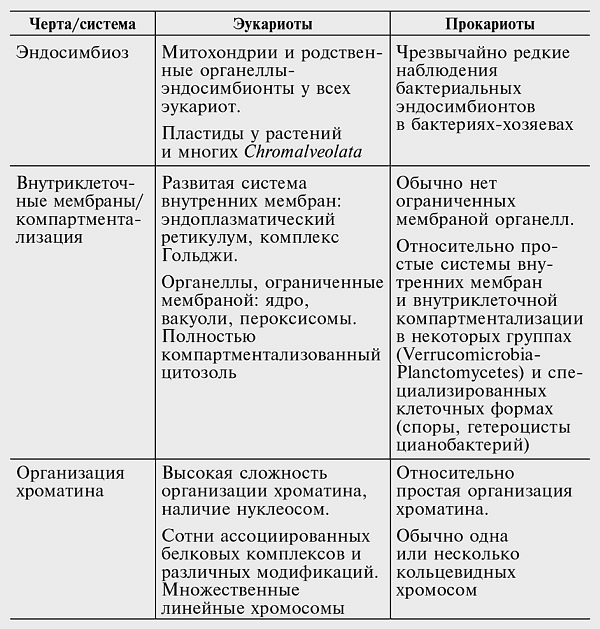
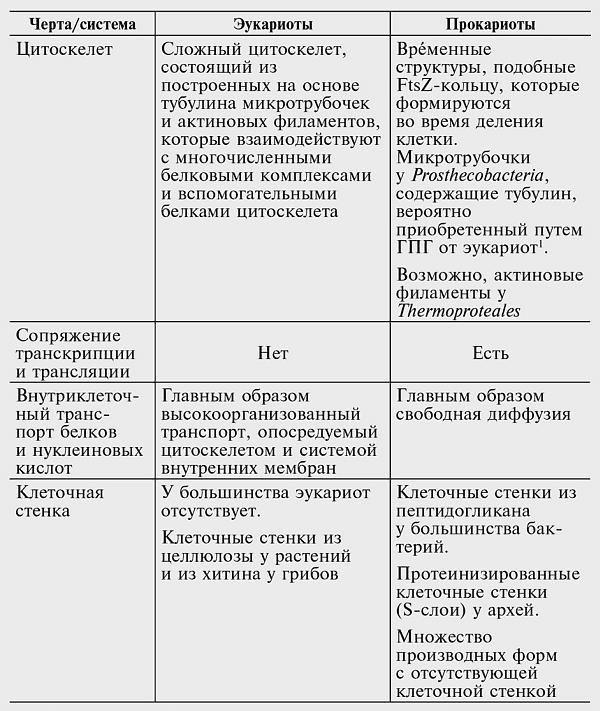
—————
1 Совсем недавно тубулин, вероятно являющийся эволюционным предшественником тубулинов эукариот, обнаружен также у архей, принадлежщих к типу Thaumarchaeota. (Yutin N, Koonin EV. Archaeal origin of tubulin. Biol Direct. 2012 Mar 29;7:10).
В главе 3 мы обсуждали некоторые из основных отличий в устройстве генома прокариот и эукариот. Далее в настоящей главе мы подробнее рассмотрим эволюцию и возможные источники одной из самых удивительных характерных черт эукариот, экзон-интронной структуры генов. Заметим, что различия проявляются на всех уровнях организации генома, от очевидных признаков, например разделения генома на множество линейных хромосом, до таких тонких деталей, как размер и строение нетранслируемых областей в белок-кодирующих генах (см. табл. 7-1).
У архей или бактерий нет ничего похожего на характерные для эукариот органеллы, особенности устройства генома и функциональные системы. Поэтому сама природа эволюционных связей между прокариотами и эукариотами кажется загадочной. В самом деле, сравнение полных геномных последовательностей однозначно показывает, что несколько тысяч эукариотических генов, отвечающих за ключевые функции клетки (трансляцию, транскрипцию и репликацию), происходят от общего предка с гомологичными генами архей и/или бактерий. Это эволюционное единство клеточных форм жизни делает объяснение того, как общие компоненты дают начало клеткам, столь непохожим в таком множестве черт, чрезвычайно сложной и интересной задачей.
В предыдущем разделе мы подчеркнули некоторые фундаментальные различия между прокариотическими и эукариотическими клетками (см. табл. 7-1). Одно из этих отличий представляется наиболее поразительным и может содержать ключ ко всей проблеме происхождения эукариот. Эта главная черта эукариотической клетки — присутствие митохондрий, играющих важнейшую роль в преобразовании энергии, а также выполняющих многие другие функции в клетках эукариот, такие как участие в различных формах передачи сигналов и программируемой клеточной смерти. Митохондрии — это органеллы характерной формы («дамская туфелька»), окруженные двойной мембраной; внутренняя мембрана содержит электронтранспортную цепь, состоящую из выстроенных в строго определенном порядке белковых комплексов. Удивительно, что митохондрии обладают своим собственным геномом, обычно представленным кольцевой молекулой ДНК, которая варьирует в размерах в разных царствах эукариот (очень небольшие, около 10 Кб, у животных, и более крупные, от 100 Кб до 1 Мб у других эукариот), и кодирующим небольшое число белков (только тринадцать у большинства животных; в основном это субъединицы комплексов электронтранспортной цепи), а также 34 рРНК и тРНК. Более крупные митохондриальные геномы растений, грибов и протист могут содержать больше функциональных генов — до ста у Reclinomonas americana из группы Excavata, — но в основном большие митохондриальные геномы состоят из встроенных мобильных элементов (Barbrook et al., 2010). Более того, митохондрии обладают собственными системами транскрипции и трансляции, которые обеспечивают экспрессию митохондриального генома. Эти системы во всех отношениях больше напоминают прокариотические, чем эукариотические аналоги. Многие эукариотические клетки содержат более одной митохондрии, и под электронным микроскопом это выглядит так, будто эукариотическая клетка нашпигована множеством паразитических или симбиотических бактерий. И на самом деле так оно и есть.
Сегодня у биологов нет сомнений, что митохондрии произошли от бактерий, бывших эндосимбионтами предков эукариот, и претерпели редуктивную эволюцию, которая превратила их в органеллы, полностью зависимые от клетки хозяина, однако сохранившие некоторые характерные прокариотические черты. Идентифицировать бактериальных предков митохондрий было относительно нетрудно (Yang et al.,1985): филогенетический анализ митохондриальных рРНК и некоторых белок-кодирующих генов точно поместил их среди альфа-протеобактерий, представителей отдельной ветви Proteobacteria, которая, что интересно, включает, наряду с большим числом свободноживущих бактерий, некоторое число внутриклеточных паразитов (таких как Rickettsia) и эндосимбионтов (таких как Wolbachia). Таким образом, по крайней мере в общих чертах, путь от альфа-протеобактерий до митохондрий кажется ясным. Однако на молекулярном уровне это превращение ни в коей мере не тривиально. В самом деле, большинство митохондриальных геномов претерпели сокращение до крайнего минимума, и это сокращение сопровождалось переносом сотен бывших бактериальных генов в геном хозяина (пока что будем использовать нейтральное определение «хозяин эндосимбионта», но позже в этой главе мы обсудим природу этого хозяина подробно). Белковые продукты большинства этих генов — включая среди прочих все белки, которые составляют митохондриальную систему трансляции, — направляются обратно в митохондрии, где они и выполняют свои функции (см. рис. 7-1). Для того чтобы этот механизм работал, гены, перенесенные в хромосомы хозяина, должны быть транскрибированы, что требует соответствующих регуляторных сигналов; транскрипты должны быть транслированы в цитозоле, что требует полной совместимости с эукариотическими сигналами трансляции; наконец, полученные белки должны быть импортированы в митохондрию, что требует специальных сигналов для импорта и специализированного белкового механизма в наружной мембране митохондрий. Проблема приспособления перенесенных генов эндосимбионта к прохождению через этот сложный путь на первый взгляд кажется совершенно неразрешимой. Однако, по-видимому, имеется очевидное решение; пока что я сохраню интригу и расскажу о нем в разделе о происхождении эукариотической клетки, ниже в этой главе.
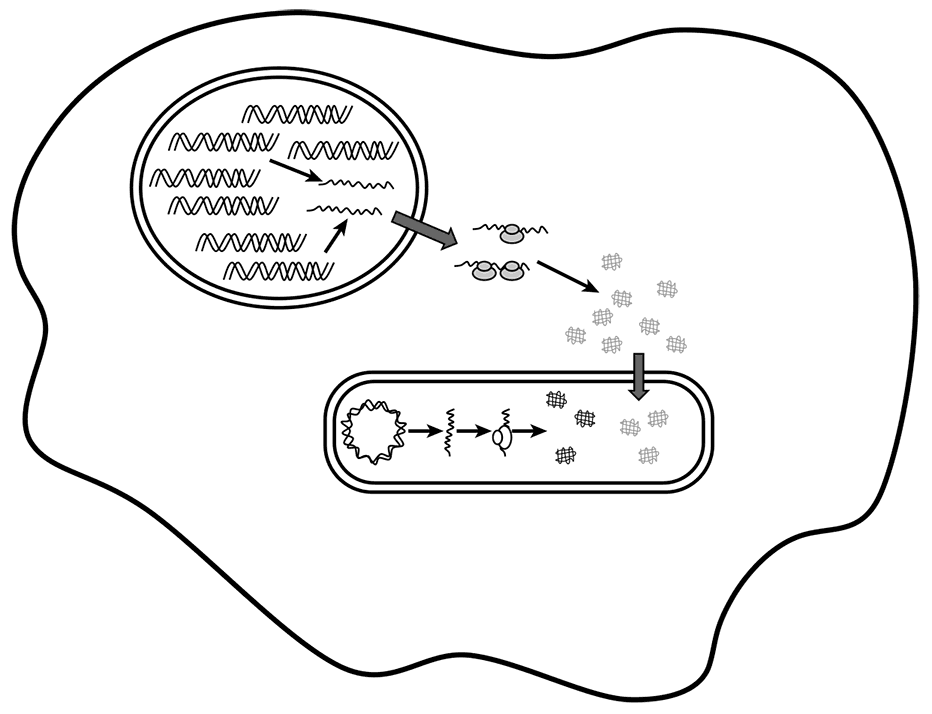
Рис. 7-1. Схематическое изображение митохондрии с ее геномом и системой трансляции, а также транспорта митохондриальных белков в эукариотической клетке.
Эндосимбиоз, митохондрии, гидрогеносомы и пластиды
Важнейшее, хотя и несколько недооцененное открытие первых лет XXI века состоит в том, что все эукариоты, изученные достаточно детально, обладают митохондриями или подобными им органеллами (Shiflett and Johnson, 2010; van der Giezen, 2009). Разнообразные одноклеточные эукариоты (протисты), такие как амебы, микроспоридии, некоторые анаэробные грибы и различные Excavata, лишены типичных митохондрий и долгое время считались примитивными, первично амитохондриальными эукариотическими формами (часто их объединяли в группу Archezoa). Однако недавние ультраструктурные исследования обнаружили ранее неизвестные миниатюрные органеллы, напоминающие митохондрии, во всех этих организмах. Эти протисты — анаэробы, так что эти органеллы, называемые гидрогеносомами, или митосомами, или просто митохондриеподобными органеллами (МПО), не участвуют в аэробном дыхании, подобно митохондриям. Однако все они обладают железо-серными кластерами, которые в митохондриальной электронтранспортной цепи являются главными каталитическими центрами. В МПО эти кластеры вместе с набором остальных ферментов катализируют другие, анаэробные окислительно-восстановительные реакции; в частности, один важный путь производит молекулярный водород, который используется метаболическими системами цитозоля. Несмотря на существенные отличия, МПО содержат некоторое количество белков, общих с типичными митохондриями. Они также используют механизм импорта белков, очень похожий на митохондриальный. Хотя эти миниатюрные МПО лишены генома и системы трансляции, которые всегда присутствуют в настоящих митохондриях, гены для нескольких характерных белков, общих для МПО и митохондрий, обнаружены в ядерных геномах соответствующих организмов. Рассматриваемые в совокупности, все эти факты не оставляют сомнений в том, что МПО представляют собой производные деградировавших митохондрий, которые, вероятно, потеряли свой геном при переходе соответствующих организмов к анаэробному образу жизни. По всей видимости, эта редукция митохондрий произошла в нескольких независимых случаях на протяжении эволюции эукариот. Отсюда следует важнейший вывод: в настоящее время нам неизвестны амитохондриальные эукариоты. Конечно, не исключено, что, пока я пишу эти строки, какие-нибудь архезои тихо размножаются, например, в маленьком пруду поблизости. Но с каждым новым эукариотическим организмом, у которого обнаруживаются митохондрии или МПО, это предположение становится все менее вероятным.
История эндосимбиоза у эукариот конечно же не ограничивается одними митохондриями. Вторым ключевым событием в истории эндосимбиоза было приобретение цианобактерий одноклеточным общим предком зеленых водорослей и наземных растений. Эти цианобактериальные эндосимбионты превратились в пластиды, которые в последующем разделились на хлоропласты и хромопласты. После цианобактериального эндосимбиоза ряд протист устроил настоящее буйство по части захвата зеленых водорослей и других обладающих пластидами эукариотических клеток. В результате возникали сложные эндосимбионты, состоящие из пластиды (возможно, это был фактор отбора для эволюционной фиксации эндосимбиоза) и остатка эукариотической клетки (часто называемого нуклеоморфом), первоначального хозяина пластид. Эндосимбиоз, по-видимому, является главным фактором расхождения Chromalveolata, одной из супергрупп эукариот (см. ниже) (Bhattacharya et al., 2007; Lane and Archibald, 2008).
В нескольких известных статьях, а также в исключительно смелой книге, написанной в соавторстве с Дорионом Саганом, Линн Маргулис, открывательница концепции эндосимбиоза в ее современном воплощении, представила эндосимбиоз как единственный доминирующий эволюционный процесс у эукариот (Margulis, 2009; Margulis et al., 2006; Margulis and Sagan, 2003). Маргулис не только предположила, что, помимо митохондрий и пластид, некоторые органеллы, такие как центриоли и жгутики, возникли путем эндосимбиоза, но даже утверждала, что эндосимбиоз лежал в основе всего видообразования у эукариот. Однако, в противоположность случаям митохондрий и пластид, эти идеи имеют очень слабое эмпирическое подтверждение либо не имеют его вовсе. Известные случаи бактериального эндосимбиоза у эукариот достаточно многочисленны, но по большей части носят переходный характер, хотя есть и замечательные исключения, такие как долговременный мутуалистический эндосимбиоз у ряда насекомых (Gibson and Hunter, 2010).
Супергруппы эукариот и корень эукариотического эволюционного древа
Как отмечалось в главе 6, древообразный эволюционный процесс гораздо лучше отражает эволюцию эукариот, чем прокариот. Главной причиной чего является частичное подавление неспецифического ГПГ, который преобладает в мире прокариот и который эукариоты заменили регулярным половым размножением (см. обсуждение ниже в этой главе). Однако принципиальная уместность метафоры древа не означает, что корректное древо легко реконструировать. Некоторые царства эукариот, такие как животные, грибы, растения и цилиаты, хорошо определены и, без сомнения, являются монофилетическими; более того, эволюционные связи внутри них в основном соответствуют дереву с вполне определенной топологией. Однако расшифровка эволюционных связей между этими царствами и рядом других групп одноклеточных эукариот (протист) — тяжелая задача, a первичное расхождение от стадии последнего общего предка эукариот (Last Eukaryote Common Ancestor, LECA) представляет собой труднейшую проблему среди всех вопросов, связанных с эволюцией эукариот (Koonin, 2010a).
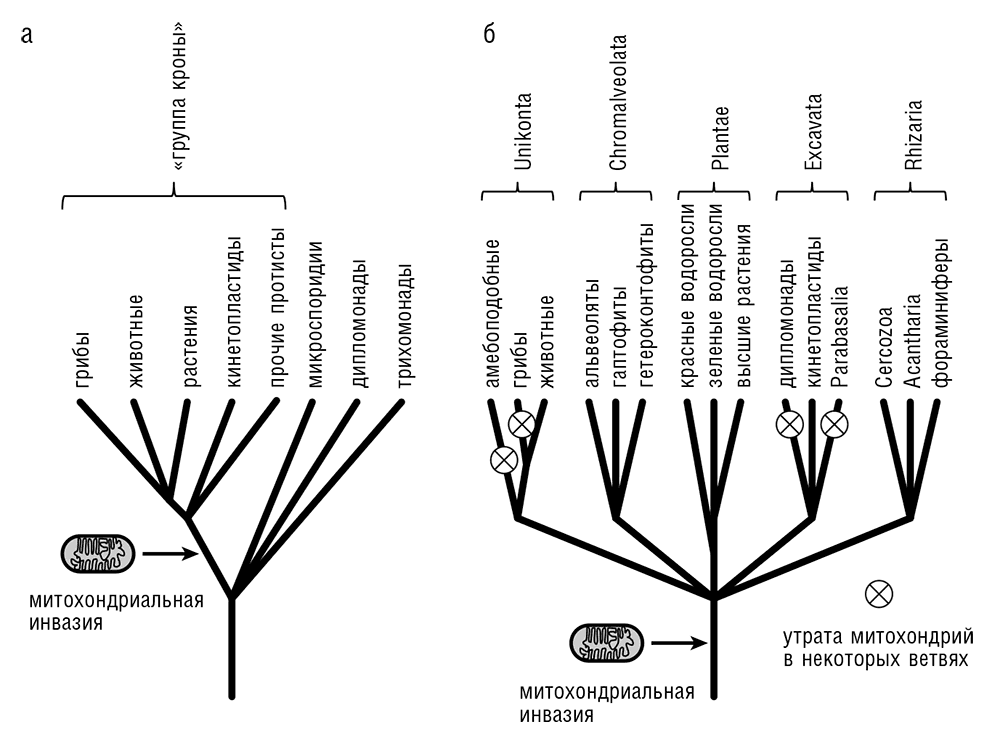
Рис. 7-2. Происхождение эукариот: а — гипотетическое архезойное древо; б — консенсусное «лучевое» древо, включающее пять супергрупп.
Проблема первичного расхождения эукариот связана с повсеместным присутствием митохондрий и МПО у эукариот, как мы уже обсуждали в предыдущем разделе. В течение многих лет большинство эволюционных биологов предпочитало так называемую филогению кроны (или «архезойное» древо), где древо эукариот состояло из «кроны», включающей животных (Metazoa), растения (Viridiplantae), грибы и некоторые группы протист, в зависимости от метода, использованного для построения древа (Cavalier-Smith, 1998; Patterson, 1999; Roger, 1999). Оставшиеся протисты, не имеющие типичных митохондрий, такие как Microsporidia, Diplomonada и Parabasalia, объединялись в группу Archezoa и считались «ранней ветвью эукариот», отделившейся от общего древа до того, как произошел митохондриальный эндосимбиоз (см. рис. 7-2а). Такая топология древа эукариот была совместима с большинством филогенетических деревьев рРНК и различных консервативных белков. В корневых деревьях, полученных путем включения прокариотической «внешней группы» (outgroup), ветви архезой обычно оказываются в стороне от «кроны», как и следует ожидать, если корень эукариот действительно находится между архезоями и «кроной». Однако в течение первого десятилетия XXI века архезойный сценарий расползся по швам (Embley and Martin, 2006). Главной причиной этого было открытие митохондрий или МПО у всех современных эукариот, подорвавшее представления о раннем отделении «примитивных» протист от общего корня. Одновременно серьезно улучшенная статистика таксонов, появившаяся в результате секвенирования разнообразных геномов, вместе с новыми, более мощными методами филогенетического анализа, показала, что глубокое размещение «рано отделившихся» групп протист, видимое на многих деревьях, представляло собой артефакт притяжения длинных ветвей, обусловленный быстрой эволюцией соответствующих групп (Brinkmann and Philippe, 2007).
Таким образом, нет оснований считать какую-либо группу эукариот примитивными, пресимбиотическими архезоями. Вместо этого, принимая во внимание небольшие геномы и высокую скорость эволюции, характерную для тех групп протист, которые раньше считались рано отделившимися (архезоями), и их паразитический образ жизни, становится все яснее, что большинство, а может быть, и все они произошли от более сложных предковых форм путем редуктивной эволюции (Brinkmann and Philippe, 2007; Koonin, 2010a). Паразиты имеют тенденцию терять гены, органеллы и функции, которые обеспечивает им хозяин (редуктивная эволюция), а также быстро эволюционировать, вследствие непрерывной «гонки вооружений» с защитными системами хозяина (более подробно об этом, в применении к вирусам, будет сказано в гл. 10). Таким образом, архезойная филогения была фактически опровергнута, и изучение ранних стадий эволюции и происхождения эукариот пришлось начать с чистого листа (Embley and Martin, 2006).
В настоящее время филогеномный подход — то есть филогенетический анализ на уровне генома — использует большие наборы консервативных генов. Важнейшим достижением на этой новой стадии было обнаружение «супергрупп», каждая из которых сочетает весьма различные группы эукариотических организмов в монофилетическую кладу (Adl et al., 2005; Keeling, 2007; Keeling et al., 2005). Большинство опубликованных до сих пор результатов филогенетического анализа дают пять супергрупп (или шесть, если не объединять Amoebozoa и Opisthokonta в супергруппу Unikonta; см. рис. 7-2б). Хотя демонстрация монофилии — нетривиальная задача для каждой из супергрупп, за возможным исключением растений, общая структура древа с небольшим числом супергрупп, образующих лучевую филогению, воспроизводится устойчиво, и последние исследования, по-видимому, подтверждают монофилетическое происхождение каждой из пяти супергрупп. Рассмотрение состава супергрупп может оказаться весьма поучительным и оказать самое радикальное воздействие на наше восприятие эукариот. Из пяти супергрупп на «лучевом» древе (см. рис. 7-2б) только три — Unikonta, Plantae и Chromalveolata (в последнем случае имеются в виду бурые водоросли) — включают сложные многоклеточные организмы, и даже в этих трех супергруппах многоклеточные организмы образуют только «кроны» (или только одну ветвь у Chromalveolata), тогда как несколько крупных ветвей в этих трех супергруппах и остальные две супергруппы состоят из протист. Хотя животные и растения конечно же наиболее заметные формы жизни, эукариотическая жизнь в основном определяется огромным разнообразием одноклеточных форм, тогда как видимые невооруженным глазом, большие многоклеточные существа суть лишь побочные ответвления в трех ветвях эукариот, образованных протистами.
Выяснение родственных связей между супергруппами представляет собой труднейшую проблему. Внутренние ветви чрезвычайно коротки, что означает быстрое (по меркам эволюции) расхождение супергрупп, возможно напоминавшее эволюционный Большой взрыв (см. гл. 6). В двух тщательных филогенетических исследованиях, в каждом из которых было проанализировано более 130 консервативных белков из нескольких десятков видов эукариот, после изучения эффекта исключения быстро эволюционирующих таксонов было получено филогенетическое древо эукариот, состоящее из трех мегагрупп (Burki et al., 2008; Hampl et al., 2009). Эти мегагруппы представлены Unikonta, Excavata и объединенной группой, составленной из Plantae, Chromalveolata и Rhizaria (см. рис. 7-2б).
Предпринималось несколько попыток вывести корень филогенетического древа эукариот (см. рис. 7-2б). Филогенетические подходы сами по себе не дают информации о корне, а использование прокариотических внешних групп приводит к потере разрешения метода, так что требуется независимая информация. Популярная идея заключается в том, чтобы попытаться идентифицировать так называемые производные состояния признака (синапоморфии), которые могут расщепить древо на два поддрева и таким образом установить положение корня. Проблема состоит в том, чтобы найти такие признаки, для которых вероятность независимого появления в двух и более линиях очень мала. Первое корневое дерево, являющееся альтернативой филогении группы кроны, было предложено Томом Кавалье-Смитом с соавторами (Richards and Cavalier-Smith, 2005; Stechmann and Cavalier-Smith, 2003). Эти исследователи использовали ясно различимые редкие геномные изменения (РГИ), такие как слияние генов, кодирующих два вездесущих фермента (дигидрофолатредуктазу и тимидилатсинтазу), а позже доменную архитектуру миозинов, и поместили корень древа между Unikonta и остальными эукариотами, Heteroconta (см. рис. 7-2б). Это разделение выглядит приемлемым с биологической точки зрения, потому что клетки Unikonta имеют единственный жгутик, тогда как у всех остальных эукариотических клеток их два. Тем не менее в этом выводе можно усомниться, так как, используя небольшое число РГИ, трудно исключить параллельное возникновение тех же самых РГИ, таких как слияние или разрыв генов, в других линиях эволюции (феномен, известный под названием гомоплазии).
Игорь Рогозин и коллеги использовали другой РГИ-подход, основанный на редких замещениях высококонсервативных аминокислотных остатков, которые требуют двух нуклеотидных замен, и пришли к заключению, что наиболее вероятное положение корня древа — между растениями и остальными эукариотами (см. рис. 7-2б; Rogozin et al., 2009). И снова представляется, что такая схема имеет биологический смысл, поскольку цианобактериальный эндосимбиоз, давший начало пластидам, произошел в линии растений и, согласно данному сценарию, мог оказаться тем событием, которое инициировало первичное расхождение эукариот. Несколько крупных ветвей Chromalveolata возникли в результате поглощения одноклеточных водорослей предковыми, беспластидным одноклеточным эукариотами (см. рис. 7-2б)[61].
Другая возможная позиция корня эукариот следует непосредственно из результатов анализа митохондриальных геномов. Как отмечалось ранее в этой главе, представитель Excavata Reclinomonas americana имеет, несомненно, самый сложный из известных митохондриальных геномов, содержащий около ста функциональных генов, тогда как у остальных эукариот их менее двадцати. Можно было бы предположить, что Reclinomonas представляет собой самую раннюю ветвь эукариот, отделившуюся от ствола древа эукариот до окончательной деградации генома эндосимбионтов. Данный сценарий поместил бы корень древа в супергруппу Excavata. Однако имеется жизнеспособная и, возможно, более вероятная альтернатива: последний этап деградации митохондриального генома наступил уже после расхождения главных ветвей эукариот и продолжался конвергентными путями в разных линиях независимо. Последний сценарий подразумевает мощный эволюционный процесс, приводящий к потере (либо к переносу в геном хозяина) всех генов эндосимбионта, за исключением малого числа тех, которые обязательно должны остаться в митохондриальном геноме для сохранения жизнеспособности митохондрий; ниже в данной главе мы обсудим возможную природу этого процесса и требования к митохондриальному геному.
Отсутствие согласия относительно положения корня древа и монофилии по крайней мере в некоторых супергрупах, не говоря о мегагруппах, указывает на то, что, несмотря на некоторые появляющиеся свидетельства, порядок первичных ветвлений в филогении эукариот в настоящее время остается неизвестным. В некотором смысле, если в раннем расхождении эукариот имел место Большой взрыв, то порядок ветвления супергрупп как таковой можно считать не имеющим особого значения. Однако биологические события, повлекшие это раннее расхождение, представляют огромный интерес, поэтому серьезные попытки разобраться в топологии самых глубоких ветвей эукариот, без сомнения, будут продолжаться с использованием более масштабных наборов данных и более совершенных методов.
Сложный, «растущий» LECA и темные века эволюции эукариот
Реконструкция LECA
Закрепление главных черт клеточной организации и, что более важно, существование большого набора генов, остающихся консервативными у всех или у большинства ранообразных эукариот, не оставляет сомнений, что все ныне существующие эукариоты произошли от одного общего предка (Last Eukaryote Common Ancestor, LECA). Как обсуждалось в начале этой главы, все эукариоты, которые были изучены достаточно детально, имеют либо митохондрии, либо МПО. Самое простое (самое экономное) заключение из этого положения вещей состоит в том, что LECA уже обладал митохондриями. И повторюсь, правдоподобность этого вывода возрастает с каждой новой описанной группой эукариот, в которой обнаруживаются органеллы, подобные митохондриям.
Реконструкция эволюции генного репертуара эукариот основана на тех же принципах и методах, что и реконструкция эволюции прокариот, обрисованная в главе 5, — в общем и целом, это принципы наибольшей экономии и наибольшего правдоподобия. Описывая суть этих подходов очень упрощенно, можно сказать, что происхождение генов, представленных у различных ныне живущих представителей главных линий эукариот, и даже предположительно потерянных в некоторых линиях, можно отследить вплоть до LECA. Результаты всех этих реконструкций согласованно указывают на сложность LECA как в отношении числа предковых генов, так и, что может быть даже более важно, в отношении наличия у предка типичных функциональных систем эукариотической клетки. Максимально экономные реконструкции, основанные на филетических паттернах в кластерах ортологичных генов эукариот, относят к LECA приблизительно 4100 генов (Koonin et al., 2004). Такие оценки очень консервативны, поскольку они не принимают во внимание главного аспекта эволюции эукариот — специфической для индивидуальных ветвей утраты предковых генов. Действительно, даже животные и растения, по-видимому наименее склонные к утрате генов эукариотические царства, явно утратили около 20 процентов предполагаемых предковых генов, идентифицированных у свободноживущего представителя Excavata Naegleria gruberi (Fritz-Laylin et al., 2010; Koonin, 2010b). Таким образом, эти реконструкции означают, что геном LECA был не менее сложным, чем геном типичных современных свободноживущих одноклеточных эукариот (Koonin, 2010a).
Этот вывод подтверждается сравнительно-геномными реконструкциями предковой композиции ключевых функциональных систем LECA, таких как поры ядра (Mans et al., 2004), сплайсосомы (Collins and Penny, 2005), аппарата РНК-интерференции (Shabalina and Koonin, 2008), системы передачи сигналов посредством убиквитина и протеасом (Hochstrasser, 2009), аппарата внутренних мембран (Field and Dacks, 2009) и аппарата деления клетки (Makarova et al., 2010). Итоги этих исследований ясны и согласуются между собой, даже когда в качестве шаблона для реконструкции используются различные топологии филогенетического древа эукариот: LECA уже обладал всеми этими структурами со всей их функциональностью, возможно столь же сложной, как и у современных эукариот.
Реконструкция других аспектов геномной композиции и архитектуры LECA также указывает на высокую сложность предкового генома. Сравнительный геномный анализ положения интронов в ортологических генах внутри супергрупп и между ними свидетельствует о высокой плотности интронов у предков супергрупп и у LECA, не меньшей, чем у современных свободноживущих одноклеточных эукариот, a вероятнее всего, близкой к богатым интронами генам животных и растений (ниже в данной главе мы вернемся к примечательной истории эукариотических интронов более подробно).
Систематический анализ широко распространенных паралогичных генов эукариот указывает на то, что LECA предшествовали сотни дупликаций, особенно генов, вовлеченных в кругооборот белков, таких как молекулярные шапероны (Makarova et al., 2005). В итоге эти результаты ясно показывают, что LECA был типичной, полностью развитой эукариотической клеткой. В последующей эволюции эукариот не проявляется постоянной тенденции к повышению сложности клеток, за исключением специфических для ряда линий «украшательств», обнаруженных в группах многоклеточных (животных, растений и бурых водорослей), а также у некоторых протист, таких как зеленые водоросли или жгутиконосцы.
Стволовая фаза: темные века эволюции эукариот
Демонстрация того, что LECA обладал высокой сложностью клеточной организации, подразумевает наличие ключевой, стволовой (то есть предшествующей расхождению ныне существующих ветвей) фазы в эволюции эукариот (см. рис. 7-3), после появления эукариотических клеток, но до появления LECA. Среди остальных эволюционных событий стволовая стадия включала широкомасштабную дупликацию ряда генов, так что набор предковых генов примерно удвоился (Makarova et al., 2005). Как долго продолжалась стволовая фаза в эволюции эукариот? С учетом того, что мы не осведомлены о каком-либо генетическом разнообразии до LECA, интуиция подсказывает, что эта фаза была очень краткой, откуда следует, что события между появлением первой эукариотической клетки и LECA развертывались в быстрой последовательности, возможно взрывообразно (см. рис. 7-3а). Однако существует совершенно законная и логически корректная альтернатива: стволовая фаза была долгой и в ней происходили постоянные расхождения, но LECA (напомню, что это последний общий предок всех современных эукариот) — это тот, кто выжил в одной из линий, тогда как остальные вымерли (см. рис. 7-3б). Некоторые из попыток датировать первичное расхождение эукариот — или, другими словами, оценить возраст LECA — дают результаты, согласующиеся с версией о долгой стволовой фазе.
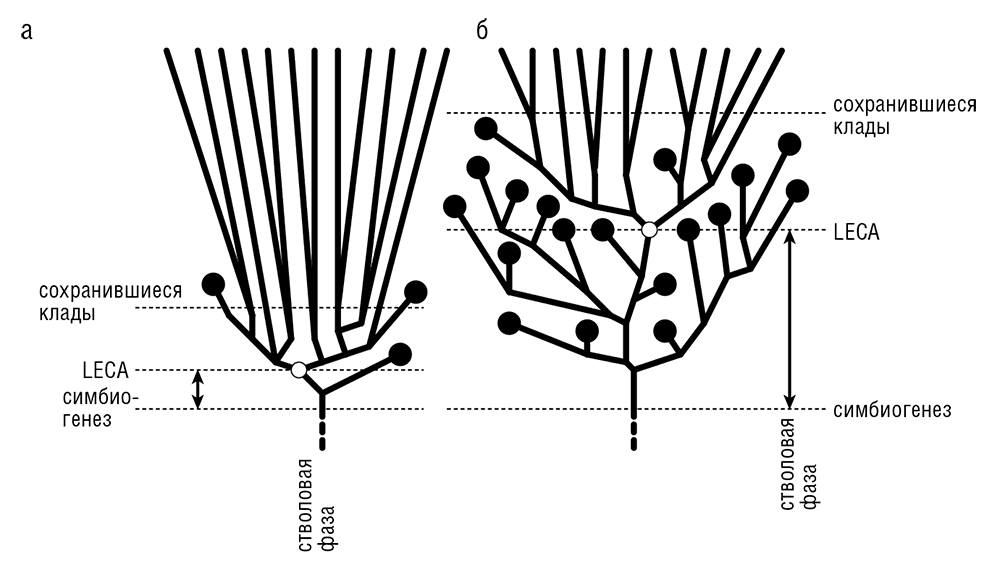
Рис. 7-3. Эволюция эукариот до и после LECA: а — сценарий «взрыва» с короткой стволовой фазой в эволюции; б — сценарий с продолжительной стволовой фазой ствола и значительным вымершим разнообразием, предшествовавшим LECA.
Молекулярная датировка событий эволюционного расхождения — весьма специализированная область исследований с множеством сложных технических проблем (Bromham and Penny, 2003; Graur and Martin, 2004). Нам придется пропустить основную часть технических подробностей и перейти прямо к результатам. Заметим что принцип состоит в том, чтобы сопоставить молекулярные данные о дивергенции (то есть результаты сравнения последовательностей, привязанные к филогенетическому древу) с палеонтологическими свидетельствами, используя нескольких точно датированных ископаемых в качестве калибровочных точек (например, самые ранние несомненные ископаемые млекопитающие датируются примерно 120 миллионами лет назад, так что это — самое позднее время начала распространения млекопитающих). Принимая строгие или нестрогие молекулярные часы, можно получить временную оценку для любого события дивергенции относительно взятых калибровочных точек и для данной топологии древа. Такие оценки могут быть достаточно достоверны, когда они представляют собой интерполяцию (то есть вывод о времени дивергенции внутри временного интервала, ограниченного калибровочными точками), но гораздо менее надежны, если получены путем экстраполяции (даты за пределами калибровочного интервала). К сожалению, для древних датировок, таких как возраст LECA, экстраполяция неизбежна. Временные оценки, полученные разными исследователями, охватывают чрезвычайно широкий диапазон дат между 1000 миллионами лет назад и 2300 миллионами лет назад. Несколько недавних, независимых и технически продвинутых исследований, использовавших нестрогие модели молекулярных часов или РГИ с похожим поведением в ходе эволюции, независимо привели к концепции «молодого LECA», которая помещает первичное расхождение эукариот в промежуток примерно от 1100 до 1300 миллионов лет назад (Chernikova et al., 2011; Douzery et al., 2004). Конечно, проблема не решена, но это, по-видимому, наилучшая из имеющихся на сегодня оценок возраста LECA. Из такой оценки следует долгая стволовая фаза в несколько сотен миллионов лет (см. рис. 7-3б), поскольку не вызывающие сомнений окаменелости эукариот относятся ко времени более чем 1500 миллионов лет назад (Knoll et al., 2006).
Этот вывод заставляет серьезно переоценить наши современные знания о ранней эволюции эукариот. С одной стороны, результаты реконструкции, описывающие LECA как одноклеточного эукариота современного типа с полностью развитыми характерными функциональными системами эукариотической клетки, становятся менее неожиданными: действительно, похоже, что имелось достаточно времени для эволюции этих черт с момента появления (примитивной) эукариотической клетки. То же самое верно и для ряда генных дупликаций, отнесенных к LECA: согласно сценарию длинного ствола, они не произошли взрывообразно; было достаточно времени, чтобы гены дуплицировались постепенно. С другой стороны, стволовая фаза — это настоящие темные века в эволюции эукариот: мы не знаем о них почти ничего и можем надеяться узнать предельно мало. В самом деле, LECA по сути является «горизонтом событий» для сравнительной геномики: используя только сравнение геномов, мы не можем заглянуть в фазу ствола. Можно получить некоторое представление о том, что тогда происходило, тщательно изучая дупликации предковых эукариотических генов, но это практически единственный источник информации о стволовой фазе. Мы не имеем представления о реальном разнообразии эукариот темных веков, и надежда на то, что мы сможем оценить его в будущем, очень мала. Данные ископаемых выявляют некоторое разнообразие, но эта летопись никогда не будет полной, и трудно даже сказать, насколько она неполна. За немногими исключениям, ископаемые останки эукариот раннего и среднего протерозоя вряд ли представляют какой-нибудь из существующих ныне таксонов — наблюдение, которое следует интерпретировать осторожно, но которое в принципе согласуется со сценарием «молодого LECA» и идеей о существовании некоторого вымершего разнообразия, которое в настоящее время недоступно для наблюдения (см. рис. 7-3б). Очевидна возможность, что LECA был тем организмом-«первооткрывателем», который захватил митохондриального эндосимбионта, и что эндосимбиоз инициировал распространение дошедших до нас эукариот. Из этого сценария следует, что предки LECA, разнообразные эукариотические Protozoa, представляют вымершую, первично амитохондриальную биоту Archezoa. Однако есть другая, возможно более убедительная версия — что митохондриальный эндосимбиоз на самом деле дал начало появлению эукариот как таковому, так что эукариоты темных веков уже содержали митохондрии либо МПО. Мы обсудим эту дилемму и доводы в поддержку гипотезы об инициированном эндосимбиозом возникновении эукариот ниже в этой главе, после того как рассмотрим связи между эукариотами и прокариотами, обнаруженные путем сравнения геномов.
Корни эукариот среди архей и бактерий
Поиск архейного и бактериального «родителей» эукариот
Все эукариоты являются гибридными (химерными) организмами как в смысле клеточной организации, так и в отношении набора генов. Действительно, как отмечалось ранее в этой главе, все ныне существующие эукариоты, по-видимому, обладают митохондриями либо МПО, произошедшими от альфа-протеобактерий, а растения и многие группы Chromalveolata к тому же содержат пластиды, произошедшие от цианобактерий. Набор генов у эукариот представляет собой разнородную смесь генов, вероятно происходящих от архей, генов с наиболее вероятным бактериальным происхождением и генов неизвестного происхождения, считающихся в настоящее время специфическими для эукариот. Может показаться парадоксальным, что, хотя филогенетические деревья, основанные на генах рРНК и соединенных последовательностях белков, участвующих в передаче информации, таких как полимеразы, рибосомальные белки и субъединицы сплайсосом, уверенно объединяют архей и эукариот, независимые сравнения полных геномов сходятся на том, что у эукариот в три и более раза больше генов с ближайшими бактериальными гомологами, чем с ближайшими архейными гомологами (см. рис. 7-4; Esser et al., 2004; Koonin et al., 2004; Makarova et al., 2005). Архейный набор значительно обогащен генами, связанными с функциями обработки информации (трансляция, транскрипция, репликация, сплайсинг), тогда как бактериальный набор кодирует в основном ферменты метаболизма, мембранные белки и компоненты биогенеза мембран, различные сигнальные молекулы и другие «операциональные» белки (подробнее см. ниже в этой главе).
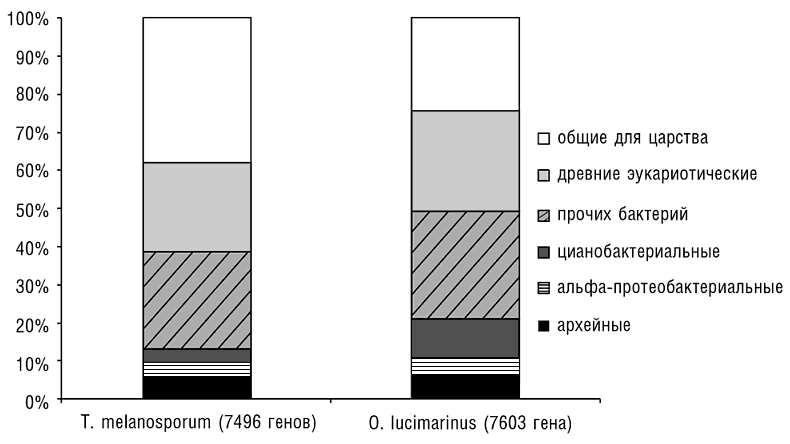
Рис. 7-4. Распределение генов двух дальнородственных эукариот, согласно их предполагаемому происхождению: архейные, бактериальные либо специфичные для эукариот. Последовательности всех кодируемых белков гриба Tuber melanosporum (черный трюфель) и зеленой водоросли Ostreococcus lucimarinus сравнивались с базой данных NCBI RefSeq программой BLASTP (Altschul et al., 1997), а предполагаемое филогенетическое родство для каждого белок-кодирующего гена определялось с использованием специально написанной программы. Стоит отметить похожие, относительно небольшие фракции генов очевидно альфа-протеобактериального происхождения и более крупную фракцию цианобактериальных генов у водоросли.
В первом приближении эти наблюдения прекрасно согласуются со сценарием слияния геномов, согласно которому эукариотический геном появился путем слияния двух предковых геномов, архейного (или родственного археям) и бактериального (вероятнее всего, альфа-протеобактериального), учитывая хорошо установленное происхождение митохондриального эндосимбионта (Embley and Martin, 2006; Rivera and Lake, 2004). Слияние геномов проще всего интерпретировать как отражение симбиогенеза. Однако попытки точно указать конкретных архейных и бактериальных «родителей» не дают однозначных результатов и, по-видимому, свидетельствуют о сложных эволюционных связях. Хотя многие из «бактериальных» генов эукариот имеют альфа-протеобактериальные гомологи, они отнюдь не преобладают среди «бактериальных» генов, показывающих явное эволюционное родство с множеством групп бактерий (см. рис. 7-4). Важная причина этого сложного распределения бактериального компонента в наборе эукариотических генов — большой размер пангенома альфа-протеобактерий (см. гл. 5). Таким образом, не идентифицировав реальную альфа-протеобактерию, давшую начало эукариотическим митохондриям, трудно определить ее генетический вклад (Martin, 1999; Esser et al., 2007). Наряду с этой неопределенностью относительно набора генов эндосимбионта невозможно исключить многочисленные источники бактериальных генов, помимо альфа-протеобактериального эндосимбионта, давшего начало митохондриям. В частности, какова бы ни была истинная природа архейного предка эукариот, он, вероятно, жил при умеренных температурах и вообще не в экстремальных условиях и постоянно пребывал в контакте с разнообразным бактериальным сообществом. Современные археи с подобным образом жизни (например, Methanosarcina) имеют ряд генов, происходящих из различных бактериальных источников, что указывает на широкомасштабный горизонтальный перенос генов от бактерий (см. гл. 5). Таким образом, архейный хозяин эндосимбионта мог уже иметь много бактериальных генов, что отчасти объясняет наблюдаемое преобладание и разнообразие «бактериальных» генов у эукариот.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава VI Этнология, происхождение и история человеческой половой жизни до брака
Глава VI Этнология, происхождение и история человеческой половой жизни до брака Самым важным, как мы видели, при изучении полового вопроса является умение оставаться в пределах объективности, избегая также сентиментального многословия, как эротизма. Но в истории половой
Аллея четвертая: УДИВИТЕЛЬНАЯ РЕШЕТКА
Аллея четвертая: УДИВИТЕЛЬНАЯ РЕШЕТКА Коллекция водных растений Потсдамского ботанического сада не представляла для меня особого интереса — я уже осмотрел экспозиции трех ботанических садов ГДР, везде водные растения были хорошо представлены, и в Потсдаме я не ждал
4. Опыты с Myxococcus xanthus показывают, что способность защищаться от обманщиков может появляться в результате единичных мутаций
4. Опыты с Myxococcus xanthus показывают, что способность защищаться от обманщиков может появляться в результате единичных мутаций Рассмотрим еще один пример, связанный с бактерией Myxococcus xanthus. Для этих микробов характерно сложное коллективное поведение. Иногда они собираются в
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции Пер. А. НадирянВ этой и следующей главах дается краткое описание современного состояния эволюционной биологии, какой она была до 1995 года, когда возникло новое направление науки — сравнительная геномика.
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции Пер. А. НестеровойВ этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись
Глава 9 Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции
Глава 9 Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции Пер. Д. ТулиноваДрама ламаркизмаКак уже отмечалось в предисловии к данной книге, одной из ключевых заслуг
Медицина и сельское хозяйство — две загубленные отрасли: удивительная параллель
Медицина и сельское хозяйство — две загубленные отрасли: удивительная параллель От того сельского хозяйства, каким его знали наши предки, традиционного, исполненного почтения ко всему натуральному, разнообразного и адаптированного к различным климатическим условиям,
Глава 10 Важность запахов
Глава 10 Важность запахов Никто не может с уверенностью сказать, когда именно на протяжении сотни миллионов лет первые живые организмы в древнейших морях начали улавливать доносившиеся до них молекулы особых химических веществ. Это произошло задолго до появления
Эндосимбиоз
Эндосимбиоз Союз двух мировДва царства живой природы — мир растений и мир животных отличаются друг от друга очень многим. Одно из отличий — хлорофилл, особое зеленое вещество. Почти все растения имеют его, животные нет. С помощью хлорофилла растения питаются блеском
8.4.11. Важность выявления людей, обладающих спонтанным даром сверхспособностей
8.4.11. Важность выявления людей, обладающих спонтанным даром сверхспособностей Известно, что, хотя и крайне редко, на Земле обнаруживаются люди, “страдающие” синдромом сверхспособностей. Одним из них является ныне живущий Ким Пик (Kim Peck), “болезнь” которого – абсолютная
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а
Актуальность (важность) вопроса
Актуальность (важность) вопроса Все части этой книги в виде отдельных статей были размещены на моем сайте и обсуждены, в том числе и специалистами в отдельных отраслях знаний. После чего я вносил в текст правки и дополнения, но вот такая отрывочность книги может вызвать
Хроника невероятных событий
Хроника невероятных событий Жил мегатерий не на деревьях, а на земле. Ходил на задних ногах, изредка лишь опираясь передними. Вытянувшись во весь рост, поднимал голову на пять метров. Ел листья. Объев те, до которых мог дотянуться, подкапывал корни дерева
Важность изучения природных популяций
Важность изучения природных популяций В предыдущем разделе были кратко изложены основы современного учения о микроэволюции. Сам факт создания этого учения знаменует собой выдающееся достижение эволюционной мысли первой половины XX в. Но нужно отметить, что это учение