НЕЙРОФИЗИОЛОГИЯ СОМАТОСЕНСОРНОЙ СИСТЕМЫ
НЕЙРОФИЗИОЛОГИЯ СОМАТОСЕНСОРНОЙ СИСТЕМЫ
Спинной мозг является главным коллектором (собирателем) сенсорных импульсов с поверхности тела. Каждый дорсальный корешок данного сегмента спинного мозга собирает информацию с поверхности тела, которая называется дерматомом. В приложении 8 показана схема таких дерматомов тела человека. Центральная веточка дорсального корешка спинного мозга содержит аксоны разного диаметра, которые проводят импульсацию от различных рецепторов. Толстые миелинизированные аксоны сообщают информацию о прикосновении и от суставных рецепторов. Тонкие нервные волокна, среди которых много немиелинизированных, проводят болевую и температурную чувствительность. После прихода нервных волокон в серое вещество дорсального рога они многократно ветвятся и образуют три типа окончаний: сегментарные (т.е. в пределах одного сегмента спинного мозга), восходящие (часть из них участвует в формировании восходящих путей спинного мозга) и нисходящие (участвуют в осуществлении межсегментарных рефлексов).
Кожа как сенсорная система. Кожа отграничивает организм от внешней среды; она содержит многообразные рецепторы, которые сигнализируют организму о контактах с внешним миром. Кожа состоит из трех слоев: эпидермиса, дермы (собственно кожи) и подкожной клетчатки (рис. 6.20). Под эпидермисом в слое соединительной ткани находится густая сеть нервных волокон с сенсорными окончаниями в эпидермисе и дерме. В дерме и подкожной клетчатке эти волокна собираются в пучки, а затем в крупные нервы, которые направляются в составе задних корешков в спинной мозг.
Рецепторные структуры кожи бывают двух видов: свободные и инкапсулированные окончания нервных волокон. Свободные окончания часто контактируют с клеткой-сателлитом эпидермального происхождения. Однако участвует ли эта клетка в реакции рецептора, – неизвестно. Самым простым типом рецептора кожи является свободный (не покрытый миелином) кончик тонкого афферентного волокна. Свободные нервные окончания находятся между клетками эпидермиса. По-видимому, значительная часть свободных окончаний представляет собой ноцицептивные рецепторы (рецепторы повреждения, болевые). Другие свободные нервные окончания связаны с ощущениями зуда, щекотки; ими же, по-видимому, воспринимается температура (терморецепторы). Некоторые из немиелинизированных волокон являются эфферентными постганглионарными симпатическими волокнами, которые иннервируют гладкие мышцы сосудистой стенки и волосяные фолликулы. Распространенным типом рецепторов являются также свободные нервные окончания, которые окружают волосяные фолликулы. Эти рецепторы возбуждаются при смещении волоса или его подергивании. В области волосяной фолликулы имеются также инкапсулированные окончания толстых миелинизированных волокон, которые, по-видимому, возбуждаются при сгибании волоса.
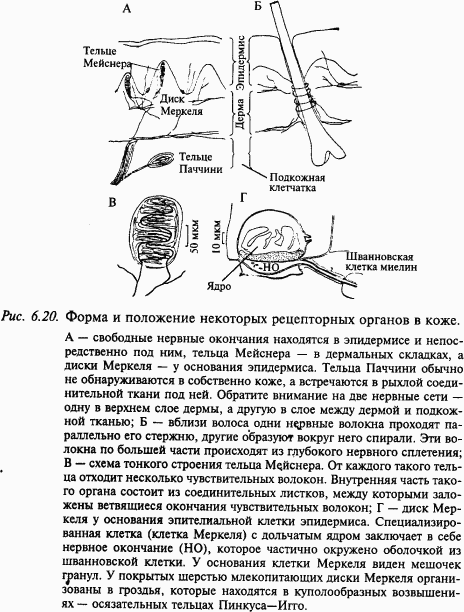
Инкапсулированные окончания составляют важный класс рецепторов кожи. Роль капсулы из соединительной ткани весьма значительна в формировании реакции рецептора. В настоящее время выделяют следующие инкапсулированные окончания кожи: диски Меркеля, тельца Мейснера, тельца Паччини, окончания Руффини. При изучении свойств кожных рецепторов трудно бывает сопоставить их физиологические характеристики с данными гистологии. Это связано с тем, что при регистрации биоэлектрической активности от афферентного волокна достаточно просто и надежно можно определить рецептивное поле данного афферентного волокна, но в этом поле могут находиться сотни рецепторов, которые передают информацию по другим афферентным волокнам. Поэтому очень трудно сопоставить тип реакции афферента с морфологическими типами рецепторов в данном участке кожи. Другими словами, можно дать физиологическую классификацию по реакциям на стимулы и распределить наблюдаемые под микроскопом рецепторы по морфологическим классам. Однако сопоставить функциональную и анатомическую классификации удается далеко не всегда. На рис. 6.20 схематически показаны те виды рецепторов, которые можно обнаружить у млекопитающих в коже, покрытой волосами (А) и не покрытой волосами (Б).
Рецепторы кожи можно разделить на быстро адаптирующиеся, или фазические (например, тельца Паччини); медленно адаптирующиеся, или тонические (например, диски Меркеля, окончания Руффини), и имеющие среднюю скорость адаптации, или фазно-тонические (например, рецепторы волосяных луковиц, тельца Мейснера).
Проприорецепторы. Известно, что млекопитающие могут достаточно точно оценивать положение конечностей, головы и туловища в пространстве. Кроме этого, достаточно точно оцениваются величина сопротивления каждому движению, углы суставов, скорость их изменения и пр. Такая оценка становится возможной благодаря рецепторам, которые находятся в мышцах, сухожилиях, суставах и называются проприорецепторами. Выше были описаны физиологические особенности двух проприорецепторов – мышечных веретен и сухожильных рецепторов. Сигналы от рецепторов кожи, мышц, суставов, сухожилий и внутренних органов поступают через задние корешки в спинной мозг. Здесь эта информация обрабатывается и отсюда передается в верхние (супраспинальные) этажи ЦНС: ядра вентробазального таламуса и далее в кору больших полушарий.
Механорецепция объединяет четыре качества: ощущение давления, прикосновения, вибрации и щекотания. Изучение механорецепции одним из классических методов: прикосновением к коже волосками, или щетинками, позволило установить, что даже легкое надавливание (в диапазоне 0,1–0,5 г, что приводит к вдавливанию кожи всего на 0,1–0,01 мм) вызывает ощущение прикосновения или давления. Но эти ощущения возникают только при надавливании в определенных точках, например кожи ладони. Эти точки называются тактильными точками. Участки кожи с большой плотностью тактильных точек находятся на кончиках пальцев и на губах. Плотность тактильных точек на животе, спине и плечах очень низкая.
Используя ножки специального циркуля (с тупыми концами, чтобы избежать болевого раздражения), можно определять пороги пространственного различения, т.е. минимальные расстояния между двумя одновременными тактильными стимулами, при которых их удается воспринять как раздельные. Исследования показали, что у взрослого человека одновременные пространственные пороги кончиков пальцев и губ особенно низки (1–3 мм), тогда как на спине, плечах и бедрах они существенно выше (50–100 мм), что согласуется с плотностью тактильных точек на этих частях тела человека.
Чувствительность к вибрации часто изучают при обследовании больных. Целью такого обследования чаще всего является нахождение порога субъективного ощущения вибрации. Наибольшая чувствительность к вибрации у человека находится в диапазоне 150–300 Гц. Минимальная (пороговая) амплитуда вибрации в этом частотном диапазоне составляет около 1 мкм.
Боль радикально отличается от других ощущений тем, что передает в нервный центр сигнал тревоги. Модальность «боль» объединяет два типа соматических болей и висцеральные боли.
Если соматическая боль имеет кожное происхождение, ее называют поверхностной болью. Если источник боли находится в мышцах, суставах или соединительной ткани, она называется глубокой болью. Таким образом, поверхностные и глубокие боли это два подкласса соматической боли. При реальном ощущении боли выделяют начальную боль, ощущаемую человеком как вспышку боли, которая исчезает с прекращением стимуляции. За этой резкой и легко локализуемой начальной болью часто следует отставленная боль, имеющая ноющий тупой характер, она начинает ощущаться через 0,5–1 с.
Болевые рецепторы в коже расположены в виде отдельных точек (дискретно). Поэтому в тех участках кожи, где их плотность невелика, всегда можно найти точки, лишенные болевых рецепторов. Специальные исследования показали, что кожа человека содержит значительно больше болевых точек, чем тактильных (соотношение 9:1). Температурные рецепторы (точки холода и тепла) встречаются в коже еще реже, чем тактильные точки, поэтому их соотношение с болевыми точками примерно 10:1.
Висцеральная боль (внутренних органов) также по своему характеру похожа на глубокую боль – имеет тупой диффузный характер. Примечательно, что внутренние органы (например, при вскрытии брюшной полости под местной анестезией) при сжимании и даже разрезах не вызывают ощущения боли, если только не задевать париетальную брюшину и корень брыжейки. Однако растяжение полых органов вызывает резкое ощущение боли. Кроме того, болью сопровождаются спазмы или сильные сокращения гладких мышц, особенно если этому сопутствует нарушение кровообращения (ишемия).
Болевые сигналы от внутренних органов часто приводят к болевым ощущениям не только в этих органах, но и распространяются на соответствующие дерматомы. Это так называемая отраженная боль. Примером такой боли может быть боль в сердце, которая ощущается в груди и узкой полоске на медиальной поверхности руки.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
§ 14. Эффекторные системы
§ 14. Эффекторные системы В основе эффекторных систем лежит всё та же способность клеток воспринимать сигналы химической, механической и электромагнитной природы. Однако источником этих сигналов становится не внешняя среда, а сама нервная система. Внутренние органы и
9.2. Сенсорные системы
9.2. Сенсорные системы Сенсорные системы возникают на очень ранних стадиях эволюции животных для непосредственного восприятия сигналов внешнего мира. В ходе этой эволюции постепенно совершенствуется воспринимающая структура, формируются аналитические центры, из
9.3. Эффекторные системы
9.3. Эффекторные системы Нервная система необходима не только для получения информации из внешнего мира, но и для адекватного ответа. Эту роль выполняет эффекторная система. В ней источником сигналов служит не внешняя среда, а сама нервная система.Эффекторная система
9.6. Функциональная асимметрия мозга и нейрофизиология речи
9.6. Функциональная асимметрия мозга и нейрофизиология речи После фундаментальных работ выдающегося нейрофизиолога Р. Сперри (1913–1994) понятие функциональных различий полушарий человеческого мозга стало общепринятым (Sperry R., 1969). Функциональная асимметрия между двумя
12.2. Системы памяти
12.2. Системы памяти Важной вехой в разработке проблем памяти явились работы канадского психолога Д. Хебба (1904–1985). В книге «Организация поведения» он первый дал обоснованную версию механизма образования энграмм как изменения эффективности синапсов (Hebb D., 1949). Эта гипотеза
12.3. Нейрофизиология памяти
12.3. Нейрофизиология памяти Уже в своих основополагающих работах Д. Хебб не обошел вниманием проблему нейрофизиологических механизмов памяти. Что показали дальнейшие наблюдения? Память и научение – это две стороны одной медали. Процессы перевода памяти в долговременную
5.6. Передача и переработка соматосенсорной информации
5.6. Передача и переработка соматосенсорной информации Чувствительность кожи и ощущение движения связаны с проведением в мозг сигналов от рецепторов по двум основным путям (трактам): лемнисковому и спиноталамическому, значительно различающимся по своим
НЕЙРОФИЗИОЛОГИЯ МОТИВАЦИЙ
НЕЙРОФИЗИОЛОГИЯ МОТИВАЦИЙ В организме под влиянием определенной физиологической потребности развивается эмоционально окрашенное состояние – мотивация. Эффективным методом исследования нейрофизиологических механизмов различных мотиваций является метод
НЕЙРОФИЗИОЛОГИЯ СТРИАРНОЙ СИСТЕМЫ
НЕЙРОФИЗИОЛОГИЯ СТРИАРНОЙ СИСТЕМЫ Полосатые тела (corpus striatum) представлены большими клеточными массами в основании больших полушарий мозга. Полосатое тело состоит из хвостатого ядра и скорлупы (вместе они составляют неостриатум, филогенетически наиболее новое
НЕЙРОФИЗИОЛОГИЯ ЗРИТЕЛЬНОЙ СИСТЕМЫ
НЕЙРОФИЗИОЛОГИЯ ЗРИТЕЛЬНОЙ СИСТЕМЫ Зрение в жизни человека имеет огромное значение. Это основной сенсорный канал, который связывает его с внешним миром. Зрительная система человека устроена очень сложно. Благодаря зрению мы воспринимаем окружающий нас мир в объеме и
НЕЙРОФИЗИОЛОГИЯ СЛУХОВОЙ СИСТЕМЫ
НЕЙРОФИЗИОЛОГИЯ СЛУХОВОЙ СИСТЕМЫ Мир, окружающий человека, полон звуков. Правда, человек слышит не все звуки этого мира, а только те, которые находятся в диапазоне от 20 до 20 000 Гц. Известно, что многие животные обладают значительно более широким диапазоном слышимых звуков.
НЕЙРОФИЗИОЛОГИЯ СЕНСОРНЫХ ПУТЕЙ СПИННОГО МОЗГА
НЕЙРОФИЗИОЛОГИЯ СЕНСОРНЫХ ПУТЕЙ СПИННОГО МОЗГА 1. Дорсальные (задние) столбы спинного мозга представляют собой мощную систему волокон различного происхождения (рис. 6.21). Задние столбы у наземных млекопитающих в эволюции формируются в связи с развитием конечностей, когда
НЕЙРОФИЗИОЛОГИЯ ОБОНЯТЕЛЬНОЙ СИСТЕМЫ
НЕЙРОФИЗИОЛОГИЯ ОБОНЯТЕЛЬНОЙ СИСТЕМЫ Обонятельную систему человека можно разделить на три отдела: периферический (нейроэпителий, выстилающий верхнюю камеру носа), промежуточный (обонятельная луковица и переднее обонятельное ядро) и центральный (палеокорковая,
НЕЙРОФИЗИОЛОГИЯ ВКУСА
НЕЙРОФИЗИОЛОГИЯ ВКУСА Функциональная роль вкусовой системы у позвоночных животных разных видов неодинакова. У наземных животных вкусовая рецепция связана с оценкой качества пищи или отвергаемых веществ, тогда как у рыб вкус играет роль дистантной рецепции и наряду с
9.5. Нервные системы
9.5. Нервные системы Так же как биение отдельной реснички на поверхности одноклеточного животного координировано с таковым соседних одноклеточных с помощью определенных физических связей, сокращение индивидуальных клеток мускулов координировано с помощью