История об истории биоразнообразия
Биологическое разнообразие характеризуется известным числом и номенклатурой различных категорий организмов, особенно животных, поскольку последние оставляют наиболее заметные и многочисленные окаменелости. Впервые история развития биоразнообразия была опубликована геологом Джоном Филлипсом, который также ввел в геохронологическую шкалу понятия «палеозой», «мезозой» и «кайнозой». Филлипс, чей фундаментальный труд вышел в 1860 году, не просто дал определения этим новым понятиям, но смог распознать в известной тогда геологической летописи крупнейшие геологические периоды: он понял, что крупные массовые вымирания древности могут использоваться для разграничения геологических эпох, так как последствия каждого вымирания приводили к появлению новой фауны. Впрочем, это не единственная заслуга Филлипса. Он также утверждал, что разнообразие видов в прошлом было намного меньше, чем сейчас, и что развитие биоразнообразия — непрерывный и всеобъемлющий рост числа видов, за исключением периодов массовых вымираний и непосредственно после них. Согласно его теории, массовые вымирания приостанавливали развитие разнообразия, но лишь временно. Взгляды Филлипса были по тем временам новаторскими. К сожалению, прошло столетие, прежде чем эта тема снова заинтересовала исследователей.
В конце 1960-х годов палеонтологи Норман Ньюелл и Джеймс Валентайн снова обратились к проблеме того, когда именно и с какой скоростью мир был заселен растениями и животными[132]. Обоих интересовало, действительно ли схема развития биоразнообразия представляла собой быстрый рост количества и разнообразия видов вслед за так называемым кембрийским взрывом около 530–520 млн лет назад (имеются в виду новые данные, а не те, что были приняты в 1960-х), а затем — относительно спокойный период. Их аргументы покоились на качестве сохранности материала в древних породах. Возможно, паттерн динамики разнообразия во времени, построенный Филлипсом, в действительности отражал лишь то, что сохранилось, а не реальную картину развития биоразнообразия. Изменения в видовом составе очевидно меньше отражены в любых более древних породах, поэтому так называемое биоразнообразие может быть просто ошибкой. Эту мысль вскоре подхватил палеонтолог Дэвид Рауп: в ряде статей[133] он настаивал, что существуют серьезные возражения против классификации обнаруженных и получивших название древних видов: древние породы подверглись большей перекристаллизации, провалам и прочим изменениям, целые регионы и биогеографические области со временем просто исчезли, и вместе с ними исчезла и более древняя геологическая летопись, то есть сейчас просто исследуются более молодые породы.

Противоположные гипотезы о диспропорции и кембрийском взрыве. «Разнообразие» относится к числу видов, тогда как «диспропорция» — к числу различных морфологических типов, анатомии. Стивен Джей Гулд считал, что во времена кембрийского взрыва наблюдалось гораздо большее (большая диспропорция) количество типов в строении тела, чем сейчас. Он ссылался на многообразие странных форм из сланцев Бёрджесс, называя их «странными чудесами», и полагал, что они относятся к типам, которые сегодня являются полностью вымершими. Противоположная точка зрения принадлежит Саймону Конвею Моррису, который утверждал, что диспропорция со временем усиливается.
Всю вторую половину XX века палеонтологи выясняли, что же происходило: постоянное быстрое развитие разнообразия видов или их развитие, стремительно достигнув какого-то определенного уровня, обрело устойчивый средний показатель скорости. В 1970-х годах Рауп, а также покойный Джек Сепкоски[134] из Чикагского университета вместе с коллегами и студентами начали активный сбор данных, накопившихся в библиотеках. Полученные данные о морских беспозвоночных, а также прочие сведения о наземных растениях и позвоночных животных вроде бы подтверждали точку зрения Филлипса: особенно кривые, построенные Сепкоски, которые показали поразительную вещь — три импульса в развитии биоразнообразия среди различных групп организмов.
Первый импульс прослеживался в кембрии (кембрийская фауна, в которую входили трилобиты, плеченогие и прочие древние беспозвоночные). За ним следовал второй — в ордовике, когда наблюдался относительно спокойный рост биомассы, и так продолжалось до конца палеозойской эры (палеозойская фауна — кораллы, замковые плеченогие, головоногие, древние иглокожие). Кульминацией стало быстрое развитие видов в мезозое, которое лишь ускорилось в кайнозое, разнообразие достигло высоких уровней, что мы и наблюдаем сегодня как результаты эволюции современной фауны (брюхоногие и двустворчатые моллюски, большая часть позвоночных, современные иглокожие и другие группы).
В чистом виде развитие биоразнообразия за последние 500 млн лет соответствовало гипотезе, представленной Джоном Филлипсом в 1860 году: на планете сейчас больше видов, чем было когда-либо в прошлом. Кроме того, что еще более утешительно, траектория развития разнообразия видов показывает, что, по-видимому, «моторчик» эволюции — процессы, отвечающие за возникновение новых видов, — неизменно работает, и в будущем можно ожидать появления на Земле еще большего разнообразия форм. В любом астробиологическом контексте эти данные свидетельствуют о том, что наша планета по-прежнему молода. Как бы то ни было, научный взгляд, имеющий 130-летнюю историю, от Филлипса до Сепкоски, очень успокаивает — сейчас видов больше, чем в прошлом, то есть вековая научная теория предполагает, что мы живем в лучшем из всех известных биологических времен (по крайней мере, с точки зрения биологического разнообразия), и есть все основания полагать, что впереди нас ждут еще лучшие времена — мир станет производительнее и разнообразнее даже без сумасшедших изобретений биоинженеров.
Хотя кажется, что работы Сепкоски убедительно показывают историю развития жизни как непрерывное движение ко все большему разнообразию, начиная с позднего мезозоя и до наших дней, тем не менее сомнения по поводу результатов исследований окаменелостей, описанных в более ранних работах, по-прежнему остаются. К тому же были проведены и другие серии экспериментов в отношении развития биоразнообразия. Больше всего обсуждался феномен под названием «ставка на последнее событие», согласно которому Сепкоски использовал методологию, не учитывающую разнообразие в глубоком прошлом, и показывал все так, будто видовое многообразие проявилось лишь в относительно недавние времена. Поскольку это действительно серьезные сомнения, ученые провели ряд исследований, чтобы проверить биоразнообразие в разные периоды. В самом начале XXI века проблема изучалась большой командой, которую возглавили Чарльз Маршалл из Гарварда и Джон Элрой, который тогда работал в Калифорнийском университете[135]. Эта группа собрала еще большую, чем Сепкоски, базу данных, основой которой послужили коллекции различных музеев, тогда как метод Сепкоски основывался на подсчете числа видов, зафиксированных в научной литературе по определенным геологическим периодам. Большинство были поражены, когда оказалось, что уже первые результаты этого исследования кардинальным образом отличались от общепринятого мнения.
Анализ, проведенный группой Маршалла и Элроя, показал, что разнообразие видов в палеозое было почти таким же, что и в середине кайнозойской эры. Резкий скачок в разнообразии видов, который долгое время принимался за аксиому, в данном исследовании не прослеживался. Выводы просты: развитие разнообразия видов достигло своего пика сотни миллионов лет назад и с тех пор остается в усредненном спокойном состоянии. Возможно, скорость развития биоразнообразия достигла своего максимума на раннем этапе развития животных, и, в противовес всем точкам зрения со времен Филлипса, это развитие больше никогда не ускорялось, а может, даже замедлялось. Хотя, разумеется, многие эволюционные новшества, например, те, что позволили распространиться растениям и животным по суше, способствовали появлению многих новых видов, и это обогатило в целом биоразнообразие, но похоже, что с конца палеозоя количество видов на планете остается почти неизменным.
Итак, после первоначального взрыва в кембрии разнообразие видов животных возрастало по экспоненте и достигло равновесия в палеозойскую эру, затем резко упало в конце пермского периода, после которого последовал неизбежный возврат к росту разнообразия, но с непременным прерыванием этого процесса короткими интервалами снижения разнообразия — массовыми вымираниями, из которых пять были особенно крупными. Хотя все периоды вымираний приводили к существенному обеднению групп организмов, за ними обязательно следовали периоды ускорения в формировании новых видов, и уровни разнообразия не просто восстанавливались, но количество видов в каждый последующий период превосходило уровень, предшествующий периоду вымирания.
Такая история развития жизни предполагает, что на наблюдаемую в фанерозое динамику разнообразия — и вымирания — влияет комплекс общих факторов. Среди многих факторов, которые позволяют объяснить возрастание разнообразия, можно назвать эволюционные нововведения, освоение до этого необитаемых и труднодоступных областей, появление новых ресурсов, а наиболее влиятельными факторами, объясняющими падение разнообразия видов, являются изменения климата, уменьшение обитаемых территорий и количества ресурсов, появление новых конкурентов в борьбе за выживание, в том числе хищников, а также внешние факторы, например, падение метеорита.
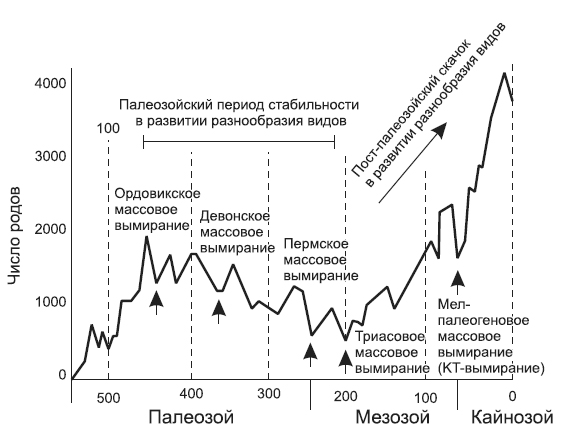
Динамика разнообразия морских беспозвоночных от начала кембрийского периода и далее, продемонстрированная Джеком Сепкоски. Его данные, основанные на долгой и кропотливой работе в библиотеках, показывают, что число родов в палеозое сначала быстро возросло, затем стабилизировалось, а в дальнейшем резко сократилось во время пермского массового вымирания. После Сепкоски отмечает большой скачок в численности родов животных, продолжающийся до наших дней.
Геохимикам давно известно, что уровни содержания в средах углекислого газа и кислорода показывают тенденции, имеющие обратное соотношение: когда увеличивается уровень кислорода, падает уровень углекислого газа. Иногда становится непонятно, как могло повлиять на развитие разнообразия животного мира изменение уровня углекислоты в концентрациях, не оказывающих никакого влияния или очень малое биологическое влияние на отдельный организм. Тогда вполне логично предположить, что дело в не уровнях углекислого газа, а в изменениях уровней кислорода, связанных к тому же с мировыми температурными режимами.
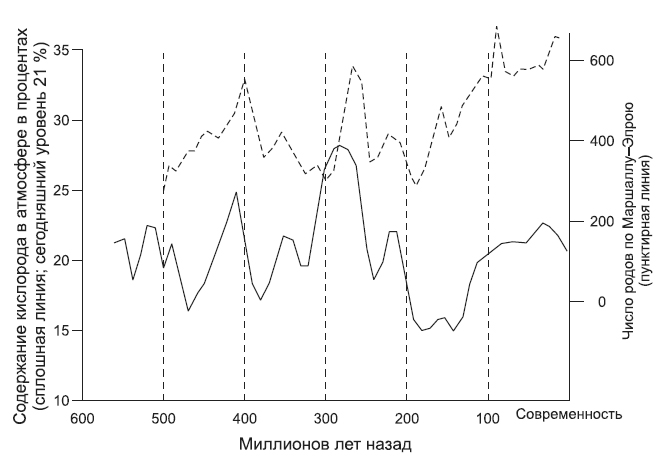
Здесь показаны уровни кислорода по работам Роберта Бернера, с учетом более новых, чем у Сепкоски, подсчетов числа родов животных, опубликованных Джоном Элроем и его соавторами. Примечательно очевидное соответствие между пиковыми показателями уровня кислорода (и высокими, и низкими) и линией, отмечающей динамику численности организмов (Питер Уорд. Из неопубликованных работ).
В холодной воде кислорода больше, чем в теплой. В холодном мире с высокой концентрацией кислорода в атмосфере жизни в океане также вряд ли будет угрожать малый уровень кислорода. С другой стороны, в теплом мире, где кислорода относительно мало, большая часть воды быстро станет застоявшейся. И не только пруды и озера, а целые океаны подвергнутся этому процессу в теплом мире, и таков мир с большим уровнем углекислого газа.
На сегодняшний день научные данные показывают, что (по крайней мере, для морских обитателей) в целом мировое таксономическое разнообразие животных соотносимо с уровнем кислорода, что не является неожиданностью, поскольку все животные не приспособлены к условиям с малым содержанием кислорода. А вот по-настоящему неожиданным оказалось то, что первоначальные уровни разнообразия организмов (измерения проводились в отношении видов и родов, которые объединены в группы по признаку общего предка) находились в обратном соотношении с уровнями кислорода. Высокие показатели возникновения новых видов, характерные для времени кембрийского взрыва (545–500 млн лет назад), приходятся на длительный период, когда уровень кислорода составлял 14–16 % (сегодня он составляет 21 %). Сильные скачки в содержании кислорода в силурийский период, а затем еще раз — в каменноугольный период соответствуют самому низкому показателю возникновения новых форм. Падение уровня кислорода в пермский период соотносится с возникновением новых организмов, но при этом — с падением общего количества видов. Кажется, это вполне ясный сигнал.
Времена, отмеченные высоким уровнем кислорода, напоминают экономический бум. Низкий уровень безработицы, бизнес процветает и разрастается, но при этом случаи открытия нового дела не так часты. Открытие нового бизнеса, как кажется, характерно скорее для периодов упадка. В отчаянные времена возникают отчаянные идеи, и люди легче идут на риск. Но если и появляется много новых предприятий, то лишь немногие становятся успешными, к тому же в плохие периоды предприятия, процветавшие в период подъема экономики, начинают прогорать еще быстрее.
Такая вот двусторонняя система: появляется больше предприятий, но большинство из них быстро банкротится и исчезает, а вместе с ними и многие, до этого процветавшие. Также происходит ослабление циркуляции денежных потоков. Общее число предприятий сильно уменьшается.
По-видимому, нечто подобное наблюдается и в развитии видов. Высокий уровень кислорода означает хороший период для развития: большое количество видов, но совсем немного новых. Но когда уровень кислорода падает, виды исчезают быстрее, чем заменяются новыми, хотя реальное число возникающих видов выше, чем в периоды с большими концентрациями кислорода.
Примеров этому не счесть. Вот один из самых ярких: долговременный период увеличения уровня кислорода, начавшийся в юрском периоде и продолжающийся по сей день, сопровождается и долговременным падением в развитии новых видов, и при этом — большим разнообразием форм. Но что принципиально нового появилось? Птицы, млекопитающие, рептилии, амфибии — все они являются кайнозойскими незначительными модификациями морфологических типов, появившихся в палеозое и мезозое («плохие времена» — низкий уровень кислорода). Прекрасный пример принципиальных эволюционных новшеств, возникших непосредственно во времена с низким уровнем кислорода, — динозавры, а в кайнозое динозавров нет.
Гипотеза, что снижение уровня кислорода вкупе с одновременным увеличением содержания углекислого газа в средах стимулировало формирование новых видов в прошлом, при этом заметно увеличив скорость вымирания, имеет серьезное биологическое обоснование. Совокупным эффектом этих процессов стало уменьшение количества видов в периоды с низким уровнем кислорода. Падение уровня кислорода с сопутствующим повышением температуры становилось худшим для организмов двойным ударом, поскольку адаптация к более жарким условиям с низким содержанием кислорода в среде обитания никогда не бывает быстрым процессом. Больше шерсти, больше перьев, больше жира на теле — все это может по-быстрому решить проблему похолодания. Но сохранять комфортную температуру тела в жару намного труднее и требует более заметных эволюционных изменений. Это утверждение еще более справедливо для ситуаций, когда животные пытаются приспособиться к условиям малого количества кислорода, поскольку такая адаптация предполагает глубокие изменения на уровне многих систем организма, начиная с состава крови, лучшей ее циркуляции, а также усовершенствованных легких или жабр.
Самый поразительный аспект процессов, связанных с кислородом и его отношением к биоразнообразию, продемонстрировал в 2009 году Боб Вернер из Йельского университета. Он обратил внимание на примечательное сходство между кривой, показывающей изменение уровней кислорода в палеозое, и кривой развития разнообразия видов, которую построили Элрой и его группа. Мы приводили эти две кривые ранее. Существует небольшое прямое соотношение между уровнями кислорода и биоразнообразием в отрезках графиков, где обе кривые имеют резкие снижения с интервалами в десять миллионов лет. Но самое удивительное, что существует соотношение между изменениями уровня атмосферного кислорода и изменениями уровня разнообразия в пределах тех самых интервалов в десять миллионов лет. Например, чрезвычайно значима и далеко не случайна корелляция между изменением процента содержания кислорода в атмосфере в период 230–220 млн лет назад и генетическим разнообразием в тот же самый период. С позиций статистики эти результаты весьма показательны.
Итак, наиболее интересным аспектом всех этих показателей является то, что с тех пор, как оба графика были представлены к обсуждению, результаты работы Вернера и его группы (да и других исследователей тоже), в которых оцениваются уровни кислорода и углекислого газа, представляются весьма неоднозначными. Так же, собственно, как и многочисленные кривые Элроя. Каждая группа исследователей выдала результаты (с одной стороны, касательно кислорода и углекислоты, с другой — численности родов живых организмов), которые получались из теоретических моделей с совершенно различными исходными данными. Ни один из множества показателей модели GEOCARB (круговорота углерода) или модели GEOCARBSULF (круговорота углерода-серы) не имеет никакого отношения к тому, сколько видов существовало в определенный период. И модель Элроя также совершенно не зависима от показателей уровней кислорода и углекислого газа. И тем не менее мы видим почти невероятное соотношение, возможно, случайное, но вряд ли — нет там никаких случайностей. По-видимому, уровни углекислого газа и кислорода (особенно кислорода) являются самыми важными факторами развития биологического разнообразия. Две независимые кривые на графиках в совокупности обеспечивают обсуждаемым здесь научным результатам самое главное требование к научному исследованию — наличие достоверности.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК