Глава 11. Механика генетического кодирования (XI)
Глава 11.
Механика генетического кодирования (XI)
Об этом можно прочитать в любом учебнике. И все же – чтобы облегчить понимание последующих рассуждений – позволим себе очень коротко остановиться на работе машины кодирования. Барбьери связывает формирование таких машин с возникновением молекул, названных им codemakers – термин, который мы выше перевели как декодеры.
Что это за молекулы? В тех случаях, когда генетическую информацию содержит ДНК (другие варианты мы не рассматриваем, поскольку они принципиально не отличаются от общей схемы), первой такой молекулой становится информационная или матричная РНК (иРНК или мРНК) – комплементарная копия отрезка той нити ДНК (минус—нити или Nonsense, N, «бессмысленной»), которая, в свою очередь, в двойной цепи комплементарна кодирующей, «смысловой» (Sense, S, то есть, содержащей ген) или плюс-нити. Между прочим, двунитевую структуру ДНК обозначают иногда не только как NS, но также – соответственно – как WC, чтобы подчеркнуть совпадающими порядками букв парадоксальный ум Джеймса Уотсона, не всеми с порога принимаемый и понимаемый. мРНК, как правило, содержит копию гена, которая, как правило же, начинается с триплета AUG, кодирующего аминокислоту метионин – М. Этот триплет называется старт-кодоном или кодоном инициации. Транскрибируемый фрагмент ДНК заканчивается сразу перед одним из кодонов терминации (стоп-кодоном). Детали матричного синтеза мРНК или транскрипции (промоторные зоны плюс-цепи ДНК, работу и характер РНК-полимераз и проч.) и детали строения самой мРНК (например, наличие и вариации поли-А сигнала и проч.) большого значения для нас здесь не имеют. У эукариот трансляции, то есть, переводу генетической информации в полипептид, предшествует так называемый процессинг мРНК, в ходе которого из тела этой молекулы вырезаются некодирующие сегменты, интроны, а оставшиеся сегменты, экзоны, формируют кодирующий полинуклеотид. У прокариот интронов нет, их мРНК практически готова к трансляции сразу после синтеза. Дальнейшие события реализации генетической информации, то есть, синтеза полипептида, кодированного в полинуклеотиде, описываются в учебниках примерно так, как коротко изложено ниже.
После транскрибирования мРНК не остается комплементарно связанной с ДНК-шаблоном; она освобождается от ДНК, которая затем восстанавливает свою двойную структуру. В одной эукаритической клетке количество молекул мРНК может составлять свыше 10.000. Наряду с молекулами мРНК на ДНК образуются и другие транскрипты, в том числе молекулы рибосомной и транспортных РНК, которые также имеют важное значение в реализации генетической информации. Все эти РНК называют еще ядерными. Наиболее обильными РНК в клетках всех видов являются молекулы рибосомной РНК (рРНК), которые выполняют роль структурных компонентов рибосом. У эукариот синтез рРНК контролируется огромным количеством генов (сотни и тысячи копий) и происходит в ядрышке. Не похоже, что структура рРНК имеет серьезное значение для обсуждаемой далее формальной организации генетического кодирования, и мы не станем на ней останавливаться. Совершенно другую роль в этих процессах имеют встречающиеся в клетке в несколько меньших количествах молекулы транспортных РНК (тРНК), которые участвуют в декодировании информации, трансляции. Это те самые декодеры или молекулы-посредники (codemakers Барбьери), – которые обусловливают специфическую связь между хранилищем генетической информации, нуклеиновыми кислотами, и строительным материалом, служащим для ее реализации, то есть для конструирования пептидов – аминокислотами.
Роль транспортных РНК в синтезе белка была постулирована еще до их открытия. В 1955г. Крик приписал им функцию адаптера, который может нести аминокислоту и образовывать водородные связи с кодирующей полинуклеотидной матрицей. Гипотеза адаптера оказалась необходимой в связи с упоминавшейся уже невозможностью обнаружить между аминокислотами и нуклеиновыми кислотами стереохимическое соответствие, достаточное для того, чтобы обеспечить считывание генетического кода. В 1957г. в лаборатории Мэлона Хогланда было показано, что в ходе белкового синтеза активированные аминокислоты переносятся на особый тип РНК, получивший тогда наименование растворимой РНК и называемой теперь транспортной.
Стереохимия тРНК хорошо изучена и весьма характерна. Мы остановимся на ней поподробнее. Типичная молекула тРНК – это полинуклеотидная цепь длиной 75—90 (по преимуществу, 76) нуклеотидов. Молекулярные массы тРНК лежат в пределах 17.000—35.000. Часть оснований нуклеотидных пар, уже после синтеза тРНК, в определенных положениях модифицирована, это неканонические, редкие, так называемые (минорные), составляющие до 10% от общего числа. Среди них – дигидроуридин (D), псевдоуридин (?) и инозин (I); последний играет существенную роль в узнавании кодона. В дополнение к этим модификациям несколько нуклеозидов метилированы. Все эти модификации – результат посттранскрипционного процессинга тРНК, которая копируется с «нормальной» матрицы. В 75% случаев молекулы тРНК открываются5?-гуанином (он фосфорилирован) и во всех случаях завершается триплетом ССА-3?.
Вторичная структура этой молекулы сформирована четырьмя короткими двуцепочечными стеблями и напоминает клеверный лист.
Каждый из четырех стеблей состоит из 4—7 уотсон-криковских пар, образующих двойные спирали. Сами стебли носят названия акцепторного, антикодонного, а также D (содержащий дигидроуридин) и T (содержащий риботимидин). Некоторые нуклеотиды консервативны, и их позиции в составе тРНК остаются инвариантными – либо полуинвариантными, если сохраняется их пуриновая или пиримидиновая природа. На акцепторном стебле тРНК имеется участок связывания с аминокислотой; он неспецифичен и для всех аминокислот один и тот же: ССА-3». Противоположный стебель содержит одноцепочечную петлю с антикодоном, распознающим кодон на мРНК. Две другие, боковые, петли предназначены для связывания с рибосомой и с аминоацил-тРНКсинтетазой (АРСазой). Четвертая, не всегда выраженная, петля так и называется – дополнительная или вариабельная (V). У тРНК, узнаваемых АРСазами класса I, она, как правило, короче (4—5 нуклеотидов), у тРНК, узнаваемых АРСазами класса II – длиннее (13—21 нуклеотидов).
Третичная (пространственная) структура любой тРНК складывает все ее четыре ветви (стебли с петлями) в так называемую Г-форму (L-форму, если использовать латиницу):
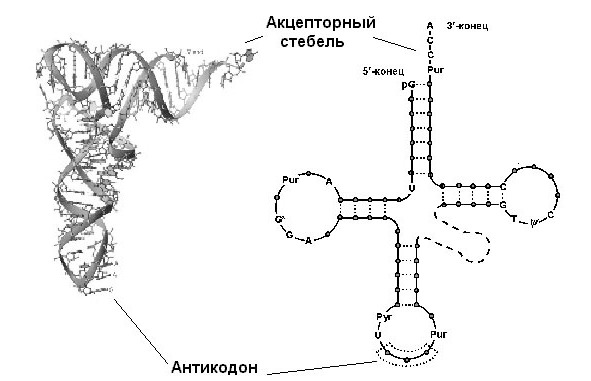
Г-форма состоит из двух почти перпендикулярных друг другу спиралей А-РНК (11 пар оснований на виток). Два отрезка буквы Г образованы ССА-3`-концом и антикодонной петлей, которые находятся на расстоянии 80? друг от друга. Наружный край угла буквы Г образован Т-петлей. Акцепторный и Т-стебли уложены один вслед за другим и образуют единую двойную спираль. В примерно такую же структуру (только с расхождением осей на 26°) уложены антикодонный и D-стебли. Эта структура на предыдущем рисунке обозначена жирной черной кривой.
Уже цитированный Др. Зенгер назвал тРНК «сокровищницей стереохимической информации». Он отмечает, что кроме уотсон-криковских пар, ответственных за большую часть горизонтальных взаимодействий между основаниями (особенно в стеблях), в тРНК имеется ряд нестандартных пар и триплетов. Такие пары располагаются в основном с наружной стороны угла и в шарнирной области буквы Г.
«Качающаяся пара» G4—U69 дрожжевой фенилаланиновой тРНК входит в состав акцепторного стебля. По структуре она сходна с уотсон-криковской и не нарушает двойной спирали, а только создает небольшую выпуклость в сахарофосфатном остове. Обратная непланарная – хугстеновская – пара m1A58-T54 возникает благодаря блокаде уотсон-криковской из-за метилированного аденина-58, и хугстеновское спаривание оказывается единственно возможным способом образования водородных связей с другим основанием (курсив здесь далее в этой главе – до новой ссылки – цитата из книги Зенгера). Пурин-пуриновая пара-m2G26-A44 «длиннее», чем обычная уотсон-криковская и в значительной мере непланарна; именно она ответственна за 26°-ое расхождение антикодонового и D-стеблей.
В тРНК имеется несколько триплетов, в которых уотсон-криковскую пару дополняет третье основание, присоединенное со стороны главного желобка либо одной (m2G10-С25-G45), либо двумя (G22-С13-m7G46 и A23-U12-A9) водородными связями; во всех этих триплетах представлена и параллельная, и антипараллельная ориентация полинуклеотидных цепей. Большинство «третичных» водородных связей образуется между консервативными основаниями.
Многочисленные вертикальные стэкинг-взаимодействия, наличие которых чрезвычайно характерно для архитектуры тРНК, укрепляют эту структуру. В дрожжевой фенилаланиновой тРНК только 42 основания из 76 входят в спиральные участки, но в стэкинг-взаимодействии участвует 71 основание. Схема стэкинга в спиральных доменах тРНК практически такая же, как и у двуцепочечных А-РНК, однако, перекрывание оснований несколько сильнее, и плотность спирали эквивалентна структуре с числом пар, несколько меньшим, чем 11. В отдельных местах молекулы тРНК основание одной цепи встраивается между двумя основаниями другой (интеркаляция), при этом возникает особый вид стэкинга.
Антикодоновый триплет также имеет спиральную структуру и формирует стопку. Кодон информационной РНК образует с антикодоном уотсон-криковские и «качающиеся» пары и, таким образом, на антикодоновом стебле появляется двойная мини-спираль. Поскольку «качающаяся» пара располагается на дальнем конце этой спирали, антикодон может найти оптимальное положение для спаривания. Другими словами, чтобы образовалась «качающаяся» пара, кодону не нужно принимать какие-то необычные конформации, а это согласуется с тем, что на рибосоме все кодоны должны находиться в одной и той же структурной конфигурации для обеспечения их геометрической эквивалентности и быстрого считывания.
Пусть – в порядке гимнастики воображения – Читатель теперь представит себе, что вся эта весьма нетривиальная структура «считывается» в клетке с хромосомной ДНК-матрицы, которая имеет самую обычную монотонную спиральную организацию.
Молекула тРНК отличается и другими структурными особенностями, но и приведенного цитирования из весьма основательного труда Зенгера достаточно, чтобы Читатель понял, что представленная в следующих главах формализация (всегда ведущая к фактическому упрощению) схемы генетического кодирования, в которой молекула тРНК занимает одну из ключевых позиций, сама по себе требует изрядной умственной отваги. Не меньшая отвага нужна, чтобы удержаться от дальнейших упрощений, отличающих, например, креационизм.
Найдено несколько десятков (теоретически 61) индивидуальных тРНК, так как каждая из них способна переносить в процессе белкового синтеза единственную аминокислоту. Конкретные тРНК называют по имени той протеиногенной аминокислоты, которую они акцептируют (например, лизиновая тРНК). Если одна и та же аминокислота акцептируется несколькими индивидуальными тРНК, то последние называют изоакцепторными и нумеруют (например, одна из тРНК для валина – тРНКвал1).
Несмотря на то, что молекулы тРНК строго специфичны, и каждой из них соответствует собственный антикодон и присоединяемая аминокислота, они не являются полноценными посредниками между этими аминокислотами и этими (анти-) кодонами. Последние – это часть структуры тРНК, что до первых, то тРНК не узнают их в принципе, да и садятся они на один и тот же триплет тРНК – концевую последовательность -ССА-3`, не различающую аминокислоты. Требуется еще один посредник, который бы узнавал обе эти молекулы так же безошибочно, как кодон узнает антикодон – и связывал их. Правила узнавания кодона и антикодона, приводящие к выбору той или иной аминокислоты для роста цепочки полипептида, называются генетическим кодом. Эти правила имеют линейный, матричный характер. Ясно, что их недостаточно, чтобы аминокислота нашла свой антикодон (в своей тРНК), поскольку 3`-последовательность, на которую она садится, у всех тРНК одна и та же. Рибосомы же, где происходит полимеризация аминокислот, различают именно тРНК. Таким образом, должна существовать еще, как минимум, одна молекула, способная различать аминокислоты и соответствующие им тРНК. Этот минимум Природа реализовала в виде молекулы аминоацил-тРНК-синтетазы, АРСазы. АРСазы способны различать и фиксировать на своей поверхности каждую из всех двадцати аминокислот. Одновременно они различают и соответствующие изоакцепторные тРНК. Рибосома имеет общий сайт связывания для всех тРНК и не различает их. Других молекулярных посредников между записанной в нуклеиновой кислоте и реализуемой в виде полипептида информации нет, и генетическое кодирование объединяет только три молекулы: тРНК с ее антикодоном, соответствующая ему аминокислота и фермент АРСаза. АРСаза способна присоединять еще и три остатка фосфорной кислоты, служащие источником энергии для реакции аминоацилирования тРНК, которую и катализирует АРСаза. Поскольку мы рассматриваем здесь не весь механизм синтеза белков, а только его кодировку, то есть, машину кодирования, а не машину синтеза, о других молекулярных деталях этой машины – различных полимеразах и других ферментах, информационных и рибосомальных РНК, самих рибосомах и т. п. – мы говорить не будем.
Таким образом, генетический код, как соответствие триплета оснований той или иной аминокислоте, реализуется не как взаимное узнавание (рекогниция, для которой не существует физико-химических оснований), а как узнавание одного и того же посредника – белка АРСазы, структура которого имеет соответствующие сайты. Это узнавание должно иметь вполне убедительную стереохимическую основу. И тем не менее, малая величина аминокислот и однотипная стереохимия тРНК представляют серьезные трудности для рекогниции. Справедливости ради стоит, однако, сказать, что тРНК все же несколько отличаются друг от друга – и не только антикодоном: имеют место небольшие нуклеотидные отличия, так что тРНК с разными антикодонами несколько различны и по своей пространственной конфигурации.
Детальному анализу структуры и функции АРСаз посвящены очень основательные обзоры Карла В?зе52; на русском языке о них довольно подробно можно прочесть в популярных статьях53 замечательного Соровского Образовательного Журнала. Нас интересуют здесь лишь основные характеристики этих ферментов. Википедия – вполне корректно – трактует АРСазу следующим образом:
«Аминоацил-тРНК-синтетаза (АРСаза) – фермент, катализирующий образование аминоацил-тРНК в реакции этерификации определенной аминокислоты с соответствующей ей молекулой тРНК. Для каждой аминокислоты существует своя АРСаза. АРСазы обеспечивают соответствие подготавливаемых ими к встраиванию в белок аминокислот и нуклеотидных триплетов антикодона тРНК, таким образом, обеспечивая правильность происходящего в дальнейшем считывания генетической информации с мРНК при синтезе белков на рибосоме.
На первом этапе происходит активация аминокислоты АТРазой:
аминокислота+ АТР?аминоацил-AMP +РРi (пирофосфат).
На втором этапе активированная аминокислота соединяется с соответствующей тРНК:
аминоацил-AMP + тРНК ? аминоацил-тРНК + АМР
Суммарное уравнение двух реакций:
аминокислота + тРНК + ATP ? аминоацил-тРНК + AMP + PPi
Сначала в активном центре синтетазы связываются соответствующая аминокислота и АТФ. Из тр?х фосфатных групп АТФ две отщепляются, образуя молекулу пирофосфата (PPi), а на их место становится аминокислота. Образованное соединение (аминоацил-аденилат) состоит из ковалентно связанных высокоэнергетической связью аминокислотного остатка и АТФ. Энергии, содержащейся в этой связи, хватает на все дальнейшие этапы, необходимые для того, чтобы аминокислотный остаток занял сво? место в полипептидной цепи (то есть в белке). Аминоацил-аденилаты нестабильны и легко гидролизуются, если диссоциируют из активного центра синтетазы. Когда аминоацил-аденилат сформирован, с активным центром синтетазы связывается 3» -конец тРНК, антикодон которой соответствует активируемой этой синтетазой аминокислоте. Происходит перенос аминокислотного остатка с аминоацил-аденилата на 2» – либо 3» -ОН группу рибозы, входящей в состав последнего на 3» -конце аденина тРНК. Таким образом синтезируется аминоацил-тРНК, то естьтРНК несущая ковалентно присоедин?нный аминокислотный остаток. От аминоацил-аденилата при этом оста?тся только АМФ. И аминоацил-тРНК, и АМФ освобождаются активным центром.
Каждая из двадцати аминоацил-тРНК-синтетаз должна всегда прикреплять к тРНК только свою аминокислоту, узнавая только одну из 20-ти протеиногенных аминокислот, и не связывая другие похожие молекулы, содержащихся в цитоплазме клетки. Аминокислоты значительно меньше тРНК по размерам, неизмеримо проще по структуре, поэтому их узнавание является значительно большей проблемой, чем узнавание нужной тРНК. В действительности, ошибки имеют место, но их уровень не превышает одной на 10,000 – 100,000 синтезированных аминоацил-тРНК. Некоторые аминокислоты отличаются друг от друга очень слабо, например, лишь одной метильной группой (I и V, A и G). Для таких случаев во многих аминоацил-тРНК-синтетазах эволюционировали механизмы, избирательно расщепляющие ошибочно синтезированные продукты. Процесс их распознавания и гидролиза называют редактированием…
Все аминоацил-тРНКсинтетазы произошли от двух предковых форм, и объединены на основе структурного сходства в два класса. Эти классы отличаются по доменной организации, структуре главного (амино-ацилирующего) домена, способу связывания и аминоацилирования тРНК. Аминоацилирующий домен аминоацил-тРНК синтетаз 1-го класса образован так называемой укладкой Россмана, в основе которой лежит параллельный ?-лист. Ферменты 1-го класса являются в большинстве случаев мономерами. 76-й аденозин тРНК они аминоацилируют по 2» -ОН группе. Ферменты 2-го класса имеют в основе структуры аминоацилирующего домена антипараллельный ?-лист. Как правило, они являются димерами, то есть имеют четвертичную структуру. За исключением фенилаланил-тРНКсинтетазы все они аминоацилируют 76-й аденозин тРНК по 3» -ОН группе. Каждый класс дополнительно делится на 3 подкласса – a, b и c по структурному сходству…
Глобула аминоацил-тРНК-синтетазы состоит из двух основных доменов – аминоацилирующего, в котором располагается активный центр и происходят реакции, и антикодон-связывающего, узнающего последовательность антикодона тРНК…»
Этот довольно пространный отрывок дает только самое общее впечатление о сложности структуры и функции АРСаз. Помимо основных описанных функций, они выполняют в клетке и другие, называемые неканоническими; мы касаться их не будем.
И все же функция упомянутого выше антикодон-связывающего домена не является абсолютным условием аминоацилирования тРНК. Нина Энтелис в связи с этим отмечает, что «для аланиновой АРСазы, например, основным элементом узнавания служит неканоническая пара G-U в аминоакцепторном стебле. При замене этой пары на G-C, A-U и даже на U-G аланиновая тРНК теряет способность аминоацилироваться аланином. Если же в любой другой тРНК заменить третью пару аминоакцепторного стебля на G-U, то эта тРНК приобретает сродство к аланиновой АРСазе и способность присоединять аланин. Таким образом, для распознавания своей тРНК аланиновой АРСазе (и она не исключение) достаточно небольшого участка аминоакцепторного стебля». У сериновой и лейциновой АРСаз E. coli антикодон также не участвует во взаимной рекогниции. Это, в частности, значит, что изменение антикодона в таких случаях – а иногда и в других, когда даже весь антикодон участвует в узнавании своей АРСазой, – не сможет повлиять на исходную специфичность аминокислоты – разве что сделает ее несколько менее эффективной.
Стоит еще раз упомянуть две особенности АРСаз. Во-первых, это очень различные в структурном отношении белковые молекулы, преимущественно классифицированные только по узнаваемому субстрату. Во-вторых, они обладают столь высокой специфичностью, что для ее характеристики даже используется особый термин – сверхспецифичность. Это свойство, отмечает Ольга Лаврик, тем более уникально, что «задачу специфичности АРСазы решают дважды: на стадии активации аминокислоты и на стадии взаимодействия с тРНК». И это при скорости роста полипептидной цепи в 20 аминокислот в секунду (для прокариот; у эукариот эта скорость на порядок меньше).
А теперь – имея в виду все, о чем мы только что рассказали, – отметим следующие два обстоятельства:
тРНК транскрибируются на геномной матрице, где естественно – как и всякие гены – подвергаются мутациям, которые приводят к точечным и другим изменениям в транскриптах (в том числе – и в антикодонах);
любая мутация по основаниям антикодона или по другим основаниям тРНК, участвующим в рекогниции АРСазами, которая может привести к изменению соответствия кодон-аминокислота, то есть к изменению кодировки немедленно исключит мутант из процесса декодирования; —
и зададимся такими вопросами: если генетический код столь феноменально консервативен, что оказался способным практически не измениться за три с половиной миллиарда лет (о чем свидетельствует его универсальность), то:
как быстро он сформировался?
почему он стал именно таким?
какие варианты могли ему предшествовать?
Ответ на первый вопрос несложен: быстро. Очень быстро – в масштабах времени, прошедших с той поры, как он сформировался. Второй вопрос вызывает встречный: Каким «таким»? Ответ на него остается загадкой, ей посвящена оставшаяся часть книги, из которой Читатель, возможно, вынесет представление о неслучайности существующей версии. Третий вопрос возвращает нас к гипотезе Георгия Гамова, с которой фактически началась эра ДНК, то есть, молекулярная биология, и с которой мы начнем ответ на упомянутый встречный вопрос в следующей главе.
…………………
Число 11, вынесенное в заголовок этой главы, хотя и менее выразительное, нежели число 111, в определенном контексте также могло бы служить информационным символом. В конце концов, параллель между тремя единицами числа 111 и триплетностью генетического кода упирается в определенное ограничение последней, поскольку генетический код триплетен лишь по размеру кодона. Функциональную же нагрузку несут в кодоне, по преимуществу, только две первые буквы, а третья служит простым разграничителем в восьми случаях из двадцати или одним из двух вариантов такого разграничителя – еще в десяти. И только два кодона являются истинно триплетными – TGG (W), и ATG (М). Еще один триплет универсального кода со всеми тремя значащими буквами – TGA – является пунктуационным кодоном.
Номер этой главы – единственный в этой книжке – совпадает со своей позицией, помеченной (гораздо менее выразительным) римским числом.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Подходы к выявлению чужеродного генетического материала в пищевой продукции
Подходы к выявлению чужеродного генетического материала в пищевой продукции Требование по маркировке продуктов, содержащих ГМ сою или кукурузу, «кроме случаев, когда отсутствуют генетически модифицированные белки или ДНК...» (Положение ЕС 1139/98) привело к развитию
1.2. Организация генетического материала
1.2. Организация генетического материала Структурно-функциональная организация генетического аппарата определяет деление всех живых организмов на прокариот и эукариот. У прокариот (к которым относятся бактерии и археи) ДНК представлена кольцевой молекулой и находится
Глава 11. Механика генетического кодирования (XI)
Глава 11. Механика генетического кодирования (XI) Об этом можно прочитать в любом учебнике. И все же — чтобы облегчить понимание последующих рассуждений — позволим себе очень коротко остановиться на работе машины кодирования. Барбьери связывает формирование таких машин с
Глава А. Аналоговые таблицы генетического кода (XIII)
Глава А. Аналоговые таблицы генетического кода (XIII) Первым, кто попытался упорядочить таблицу генетического кода и построить ее на рациональной основе, был наш выдающийся ученый Юрий Борисович Румер. Он был физиком, учеником Макса Борна, хорошо знал Альберта Эйнштейна,
Глава Б. Барионная оцифровка генетического кода (XIV)
Глава Б. Барионная оцифровка генетического кода (XIV) ФОРМАТЫ 1D и 2D Строго говоря, барионным числом называется сохраняемое квантовое число системы. Нам нет необходимости углубляться в эту тему. Может быть, стоит помнить лишь то, что барион — это элементарная частица,
Глава A. Аналоговые таблицы генетического кода (XIII)
Глава A. Аналоговые таблицы генетического кода (XIII) Первым, кто попытался упорядочить таблицу генетического кода и построить ее на рациональной основе, был наш выдающийся ученый Юрий Борисович Румер. Он был физиком, учеником Макса Борна, хорошо знал Альберта Эйнштейна,
Механика развития и менделевская генетика
Механика развития и менделевская генетика К концу XIX в. ощущалась все большая напряженность во взаимоотношениях между двумя главными философскими подходами к биологии - Аллен (Allen) называет это расхождением между натуралистами и экспериментаторами. Натуралистов
Глава 9 Механика красоты
Глава 9 Механика красоты К чему вздыхать, красотки, вам? Мужчины — род неверный: Он телом — здесь, душою — там, Все ветрены безмерно. Уильям Шекспир, «Много шума из ничего». Акт II, сцена 3 (пер. Т. Щепкиной-Куперник) Сегодня сразу три команды американцев ищут «голубой ген»,
Изучение динамики генетического состава популяции
Изучение динамики генетического состава популяции В начале этой книги подчеркивалось, что одна из важнейших задач современного популяционного исследования — получение материалов по самым разнообразным эволюционным ситуациям в природных популяциях, в частности,