Генетический алфавит
Генетический алфавит
Авторы модели ДНК представляют систему «кодирования» следующим образом: различные свойства ДНК в разных организмах объясняются различием в составе нуклеотидов в ее молекуле. Нуклеотиды, по их мнению, являются своего рода генетическим алфавитом, при помощи которого в ДНК «химическим почерком» закодированы наследственные особенности организма. А поскольку ДНК содержит по меньшей мере несколько тысяч нуклеотидов, возникают неограниченные возможности комбинаций во взаимном чередовании и порядке пар А — Т, Г — Ц в ее молекуле. Каждая новая комбинация придает новые свойства ДНК.
Молекулы ДНК в клетке, поясняют далее Дж. Уотсон и Ф. Крик, способны к «самовоспроизведению» (редупликации) с сохранением постоянного расположения нуклеотидов. Мы знаем, что внутренний «цилиндр» молекулы ДНК состоит из пар оснований А — Т и Г — Ц, соединенных между собой водородными связями. Редупликация молекул ДНК может идти следующим образом. Водородные связи между парными нуклеотидами на одном конце молекулы постепенно начинают прерываться и обе спиральные цепи ДНК «разматываются», освобождаясь друг от друга. Такая развернувшаяся цепь удерживается связями между остатками молекул дезоксирибозы и фосфорной кислоты, «выставив» перпендикулярно оси свои основания. В окружающей среде находятся синтезированные клеткой свободные нуклеотиды, которые могут входить в реакцию со свободными основаниями развернутой цепи ДНК. Но к каждому основанию может приблизиться и соединиться с ним только нуклеотид, имеющий парное, «дополняющее» основание. Это значит, что к развернутой цепи начнет присоединяться другая, недостающая цепь ДНК, причем точно такая, какая была в другой половине макромолекулы и отделилась, чтобы подобным же образом извлечь из среды то, чего ей недостает до полной молекулы ДНК. В результате этих процессов образуются две молекулы ДНК, каждая из которых имеет половину материнской молекулы, дополненную вновь синтезированной. Дочерние молекулы становятся, таким образом, точной копией материнской ДНК. При этом сохраняется и состав генетического материала.
Здесь мы должны, однако, предупредить читателя, что то, о чем он только что прочел, было всего-навсего гипотезой Уотсона и Крика о редупликации ДНК. Вы можете справедливо заметить, что правильность гипотез должна быть доказана экспериментально.
Описанную гипотезу подтвердил американский биохимик А. Корнберг, которому удалось открыть полимеразу ДНК — фермент, участвующий в процессе редупликации. Получив из клеток кишечных бактерий Escherichia coli ДНК и фермент, он соединил их в пробирке, добавив туда же смесь соответствующих нуклеотидов. Через некоторое время количество ДНК в пробирке значительно увеличилось, причем были использованы присутствующие в среде свободные Нуклеотиды. За свое открытие он получил в 1959 году Нобелевскую премию по физиологии и медицине. Эту премию разделил с ним профессор С. Очоа, открывший независимо от Корнберга фермент — полимеразу РНК, которая синтезирует рибонуклеиновую кислоту. Совсем недавно Корнберг обнаружил еще один фермент, участвующий в синтезе ДНК, и назвал его фосфатазой ДНК.
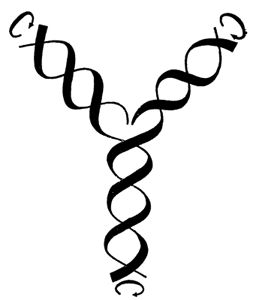
Модель редупликации молекулы ДНК. К развернутым цепочкам (вверху) присоединяются дополнительные цепочки того же состава, что и в материнской молекуле.
Гипотезу о редупликации ДНК подтвердили в 1958 году М. С. Месельсон и Ф. Сталь. Они культивировали бактерии Е. coli в жидкой питательной среде, содержащей вещества с радиоактивным азотом 15N. ДНК этих бактерий оказалась потом «меченной» радиоактивным элементом всюду, где в ее макромолекулах содержится атом азота. Затем ученые культивировали бактерии в жидкой питательной среде, содержащей нерадиоактивный азот 14N. При выращивании бактерии размножались, и можно было наблюдать, как радиоактивный азот ДНК равномерно распределялся по дочерним молекулам ДНК. В каждой последующей генерации бактерий отмечалась половинная радиоактивность ДНК по сравнению с предыдущей. Иначе говоря, при каждой редупликации ДНК радиоактивность материнской макромолекулы равномерно распределялась в двух дочерних макромолекулах, которые создавали недостающие им цепи, привлекая для этого нуклеотиды с нормальным азотом.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Жакоб и Моно обдумывают генетический код
Жакоб и Моно обдумывают генетический код Итак, по мнению Крика и его коллег, ДНК можно считать химическим носителем наследственности, иначе говоря, ДНК — это определяющая составная часть генов. Бидл и Тейтем уже давно сформулировали положение «один ген — один фермент»,
3.2. Генетический материал эукариот
3.2. Генетический материал эукариот Генетический материал эукариот сконцентрирован в ядре и представлен хромосомами, в которых молекула ДНК образует сложный комплекс с различными белками.Каждая клетка любого организма содержит определенный набор хромосом.
4.2. Генетический анализ
4.2. Генетический анализ Совокупность методов изучения наследственности получила название «генетический анализ». Его основа – гибридологический метод, разработанный Г. Менделем. С открытия законов наследования Г. Менделем и начинается история генетики. Не меньшая
6.2. Генетический код
6.2. Генетический код Генетическая информация записывается последовательностями нуклеотидов в нуклеиновых кислотах с помощью 4 символов, как информация текста книги записывается с помощью букв, количество которых зависит от конкретного алфавита. В структуру белка эта
ГЕНЕТИЧЕСКИЙ АНАЛИЗ
ГЕНЕТИЧЕСКИЙ АНАЛИЗ В 1980 годах профессор Алек Джеффрис из Лестерского университета доказал наличие многочисленных участков ДНК, которые не следует считать кодом аминокислот. Эти участки назвали минисателлитные ДНК. Тысячи ДНК разбросаны по всем хромосомам; возможно,
Глава G@C. Генетический код явление "героя" (X)
Глава G@C. Генетический код ? явление "героя" (X) События, связанные с эволюцией Вселенной и коротко описанные выше, привели, в конечном счете (а может быть, и «в том числе») к возникновению жизни, центральным феноменом которой стало объединение мира нуклеиновых кислот и мира
Генетический код
Генетический код Поскольку информация о структуре белков в ДНК и и-РНК записана последовательностью нуклеотидов, для перезаписи в последовательность аминокислот должна существовать система кодировки, которая получила название «генетический код».Генетический код –
Как взломать генетический замок
Как взломать генетический замок Однако эволюционные черепахи перемешивают свои гены сильнее, чем эволюционные зайцы. Открытие Остина Берта о связи между длиной поколения и количеством рекомбинаций свидетельствует о том, что Черная Королева работает. Чем длительнее
Глава 4 Пол и генетический мятеж
Глава 4 Пол и генетический мятеж Черепаха живет между двух пластин, Она или он — вид один. Черепаха, я думаю, очень умна, Раз так, притом, плодовита она. Огден Нэш (пер. О. Волковой). В средневековых английских деревнях одно пастбище было общинным. Каждый имел право пасти
Приложение. Генетический код
Приложение. Генетический код Генетический код — это небольшой словарь, который устанавливает связь между языком нуклеиновых кислот из четырех букв и языком белков из двадцати букв. Каждый триплет оснований соответствует определенной аминокислоте, за исключением трех
Генетический допинг
Генетический допинг Генотерапия Мы знаем, что человек страдает тяжелыми заболеваниями, если имеет дефектные варианты генов. Изменения в гене, из-за которых может возникнуть болезнь, называют патогенной мутацией. Сегодня известны примерно 1500 генов, несущих в себе
Генетический возраст
Генетический возраст Итак, человек умирает. Ну хорошо – все мы рано или поздно умрем. Но как это происходит? Большинство людей, с которыми я обсуждал этот вопрос в винном ресторанчике Heuring, постоянно спрашивали: если человек никогда не болеет, он все равно умрет? Есть у
Генетический рецепт
Генетический рецепт Для меня генетическое будущее человечества, однако, выглядит совершенно иначе. И то, о чем я хочу поговорить в заключение этой книги, не когда-нибудь появится, а уже есть. Я считаю, что будущее принадлежит не изменениям в генетике человека. Знание о его
Генетический пол
Генетический пол Первым в списке Мани значится хромосомный (или генетический) пол. Генетические различия между мужским и женским полом представляют собой фундаментальную основу феномена пола, отражающую важнейшую черту полового размножения. У подавляющего большинства