Некоторые элементарные сведения о происхождении и систематике гидрофитов
Некоторые элементарные сведения о происхождении и систематике гидрофитов
Теперь, когда мы знаем, чем отличаются водные растения от растений суши, пора, вероятно, поговорить об их происхождении.
Эволюция, отмечает академик А. Л. Тахтаджян в книге «Система и филогения цветковых растений» (1966, с. 14), подобна своего рода стратегической игре живых организмов со средой, причем игра эта антиэнтропийна, значит, неупорядочена, живые организмы предпринимают в своем развитии и приспособлении к среде чисто случайные ходы, одни из которых оказываются неуспешными и даже вредными — и это ведет к гибели не сумевших приспособиться организмов, а другие — случайно удачными, способствовавшими лучшей приспособляемости — и это ведет к прогрессу данных организмов, когда эти удачные находки закрепляются в наследственной информации.
Жизнь зародилась на нашей планете в водной среде (в океане), там сформировались и первые растения — водоросли; они освоили не только море, но и пресные воды. Современные водоросли — потомки древних, вымерших — являются настоящими водными, или первичноводными, растениями: вода для них — среда первичная и единственная (лишь очень немногие из современных водорослей живут на суше).
Иная картина эволюции высших водных растений, главным образом цветковых. Огромным прогрессивным шагом в эволюции всего живого на нашей планете явился когда-то в прошлом выход предков теперешних растений из первичной среды на сушу*. В ходе эволюции растения все далее отходили от берегов, осваивали все новые участки суши, многообразились, все более отличались друг от друга, все более совершенствовали свою структуру и приспособляемость к различным условиям среды на суше. В водную среду эти растения пришли с суши, как гости, на разных этапах эволюционного развития. Отсюда разная приспособляемость гигрофитов к жизни в водной среде. И определяют эти растения как вторичноводные, в отличие от первичноводных, которые никогда не выбирались из воды.
* В этом процессе далеко не все ясно до сих пор — почитайте об этом в книге С. В. Мейена «Следы трав индейских» (1981, гл. 1).
«Развитие растительного мира от низших форм к высшим, — пишет С. А. Шостаков в книге «Систематика высших растений» (1971), — шло в основном по пути приспособления к воздушной среде. Высшие растения, как правило, сухопутные формы. Редкий водный образ жизни здесь всегда вторичное явление». И далее С. А. Шостаков подчеркивает: «Жизнь в водной среде, где все части растения находятся в одинаковых условиях существования, не является стимулом к дифференциации, т. е. расчленению растительного организма».
Чтобы достигнуть такого сложного строения, гидрофитам пришлось в филогенезе пройти «суровую школу жизни» на суше.
Адаптивная эволюция растений, по словам Д. Л. Тахджяна, протекает через три стратегических этапа. Первый — это прогрессивная эволюция, совершенствование всего организма. У растений постепенно развиваются механические несущие ткани, система проводящих сосудов, корни, стебли, листья, цветки — разнополые или одновременно имеющие мужские (тычинки) и женские (пестик) органы, — возникает восходящий ток соков от корней к листьям и т. д. Но наряду с дифференциацией и морфологическим усложнением возникает на этой стадии и более высокая интегрированность, гармонизация частей растения, слаженно и четко выполняющих свою функцию и обеспечивающих жизнестойкость всего растения. Возрастает сумма взаимоотношений со средой, ее разнообразными элементами, возникает до известных пределов свобода, вариантность этих взаимоотношений, что позволяет растению захватывать, осваивать все новые участки среды. Такое совершенствование взаимосвязи со средой шире морфологического усложнения.
Возьмем в качестве примера известное растение — гречиху. Она растет у дорог, иногда на довольно сухих участках, еще лучше — во влажных местах, но может расти и в воде, располагая листья на поверхности. Даже полностью погруженная в воду она не погибнет, правда, остановит рост на время, пока не спадет вода. Если не дождется — начнет тянуть хилые листья к поверхности. А дотянется — снова наберет силу, будет нормально расти и цвести. Недаром ее называют гречихой земноводной.
А тропический род гигрофил? Могут расти у дороги, на лугу, в дождевом лесу, у воды, полупогруженные, в воде с верхушкой стебля на поверхности воды и полностью погруженные. Например, криптокорины встречаются на стремнинах, в медленно текущих и стоячих водах — это водные формы; полупогруженные живут в мелкой воде; а наземные формы растут у берега, в низинах, во влажной подстилке леса. Один вид — реснитчатая криптокорина — приспособился жить даже в морской воде среди корней мангровых деревьев.
Конечно, свобода адаптации не бесконечна — ни гигрофилы, ни криптокорины сухих почв и воздуха не перенесут.
Вторым этапом в стратегии эволюции является специализация — возникновение частных, ограниченных приспособлений к существованию в определенных условиях среды. Специализация позволяет организмам расширять территорию своего распространения, приспосабливаться к новым условиям среды. Это плюс. Но есть и минус: всякая специализация заужает возможности растений. Именно такой узостью обладают гидрофиты: при изменении условий среды им трудно «переспециализироваться». Значит, с увеличением специализации снижается способность к адаптации.
Обратимся к примерам. «Наиболее водным» растением можно назвать роголистник. Он и растет под водой, и цветет в воде — без воды он совсем не может существовать. Кувшинка тоже водное растение. Но у нее есть развитая корневая система, а листья и цветки связаны с воздушной средой — специализация есть, конечно, но она не достигла такой степени, как у роголистника. Поэтому и способность приспосабливаться к изменившимся условиям среды у нее значительно выше. В 1912 г. в журнале Киевского общества любителей аквариума был опубликован снимок, вызвавший сенсацию: на потрескавшемся, пересохшем илистом грунте сидят небольшие толстенькие ярко-зеленые кустики кувшинок. Сидят и ждут осенних дождей, чтобы вернуться к обычной «кувшиночной» форме. Именно этот снимок надоумил ленинградского садоводаН. Гаврилова организовать перезимовку кувшинок в горшках, пока ремонтируют бассейн.
Однажды в оранжерее Главного ботанического сада АН СССР в Москве я увидел интересное растение: из бетонного ящика густо тянулись вверх 15-сантиметровые мясистые черешки, на концах которых были устремившиеся вверх, толстые, плотные, дисковидные пластинки листьев. — Да это же лимнобиум, который вы мне привезли два года назад! — сказала мне садовод В. Чеканова. — Просто бассейн подсох немного, давно не подливала в него воды.
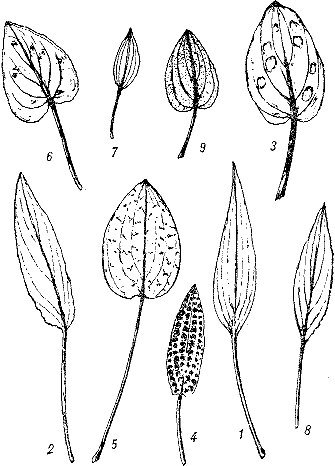
Разнообразие листьев криптокорин: 1 — Cryptocoryne affinis, 2 — С. beckettii, 3 — С. blassii, 4 — С. bullosa, 5 — С. griffithii, 6 — С. longicauda, 7 — С. minima, 8 — С. nuri, 9 — С. thwaitesii (по Г. Брюннеру).
— Лимнобиум — тропический лягушатник, — изумился я, — эта розетка круглых зеленых листьев, плавающая на поверхности аквариума?
Впрочем, когда я спустя год увидел такой же лимнобиум в бассейне под парниковой крышкой у известного садовода И. Л. Заливского, пришлось еще раз удивиться: на поверхности воды плавала тоже розетка, но диаметром не 8 — 12 см, а всего 30! И листья были не светло-зеленые, а вишневые. Вот что сделали с этим растением 30° С в парнике в сочетании с ленинградским весенним солнцем. Ясно, что узкая специализированность, «степень водности», этого растения тоже меньше, чем у роголистника или валлиснерии, которые не могут существовать без воды.
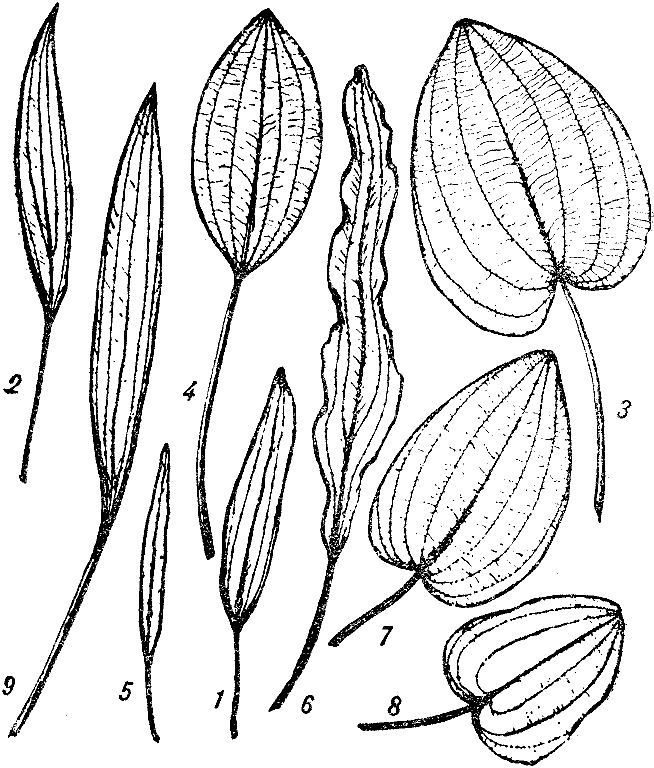
Разнообразие листьев эхииодорусов: 1 — Echinodorus berteroi, 2 — Е. amazonicus, 3 — Е. cordifolius, 4 — Е. andrieuxii var. longistylis, 5 — E. austroamericanus, 6 — E. maior, 7 — E. scaber, 8 — E. nymphaeifolius, 9 — E. bleheri (по Г. Брюннеру).
«Среда, — отмечает А. Л. Тахтаджян, — многообразна и неупорядоченна. Осваивая новые участки ее, новые ареалы, растения изменяются соответственно характеру условий в этих ареалах». Постепенно происходит расхождение признаков у этих растений по сравнению с исходной формой. Сначала эти изменения касаются вегетативных органов, потом — наиболее консервативных — цветков. Сначала эти изменения неустойчивы, характерны только в данных условиях, а в иных — растение возвращается к исходной форме. Потом эти изменения постепенно фиксируются, становятся устойчивыми, передаются по наследству. Происходит дивергенция — расхождение признаков, образование новых разновидностей, а затем и видов.
Третьим этапом стратегии эволюции является регресс, или общая дегенерация. Так, приспособившись к жизни в воде, водные растения стали утрачивать отдельные органы, например, корни. Все живущие в воде растения утратили механические несущие ткани. А ряска пошла по этому пути еще дальше — она утратила не только корневую систему, но и листья, цветет очень редко и размножается преимущественно вегетативно. Такое упрощение обеспечивает успешное развитие и распространение вида. А расширение ареала и есть, по А. Н. Северцову, один из показателей прогрессивного развития вида.
Итак, на первом этапе эволюции водных растений формировалась совершенная структура, дифференциация и многогранность взаимодействия со средой. Этот этап растения проходили на суше, и только на суше. На втором этапе, когда гидрофиты снова «отправились» в воду, возникла специализация. На третьем этапе гидрофиты уже успешно освоили водную среду и избавились от приспособительных приобретений первого этапа, которые в однообразно устойчивой среде оказались лишними. Таков путь, пройденный в эволюции вторичноводными растениями.
Для того чтобы нам легко было ориентироваться в мире современных гидрофитов, познакомимся с некоторыми положениями систематики.
Высшие растения делятся на отделы, классы, порядки. Порядок делится на семейства, каждое из которых подразделяется на роды, а род — на виды. Вид назван Международным кодексом биологической номенклатуры основным рангом в классификации. «Каждое растение, — замечает А. Л. Тахтаджян, — с которым имеет дело исследователь, должно быть определено с точностью до вида, а во многих случаях даже точнее». Чтобы ученые разных стран, публикуя свои исследования на родном языке, могли при чтении работ друг друга или при встрече, понимать, о чем идет речь, решено было в науке пользоваться международным языком, понятным всем ученым мира. Для научного общения был избран латинский язык, на котором когда-то говорили жители древнего Рима. Теперь этот очень красивый, стройный и удобный язык мертвый, ничей, — на нем никто не говорит. Ученые всего мира договорились давать научные, понятные всем ботаникам мира названия растений на этом языке. Но разве нельзя обойтись русскими названиями? К сожалению, не всегда.
Русские названия — полушница, стрелолист, водяная лилия, водокрас, телорез, чилим... — издавна известны нашему народу, но для ботаника из другой страны они непонятны, ведь на его родном языке названия этих же растений звучат совсем иначе. Кроме того, народные названия не всегда точны. Водяная лилия, например, совсем не похожа на садовые лилии, а название «стрелолист», полученное растением благодаря сходству его листьев с пучком стрел в колчане, не может объединить растения, относящиеся к одному роду, но имеющие другую форму листьев. Кроме того, растения, обитающие, например, в тропических районах земного шара, часто не имеют исконно русских названий.
В ряде случаев ученые переводят на русский латинские названия. Однако иногда переводом не только трудно, но и неудобно пользоваться. Например, «растение, образующее колючие семена на соцветии» — это перевод одного слова «эхинодорус». Или апоногетон: А. Вендт посвятил полстраницы выяснению, похоже ли это слово на греческое, выдвинул несколько гипотез о его происхождении, а в итоге признался, что «происхождение этого названия не ясно...» Дословный перевод латинских названий на русский язык часто бессмыслен и не нужен.
В этой книге мы будем пользоваться латинскими названиями для тех растений, у которых нет обоснованных русских, и для мало распространенных.
Семейство обычно называется по названию самого характерного для него растения. Скажем, семейство алисмовых (Alismaceae) получило название от названия растения алисма (частуха). Но в семейство входят и другие роды — бальделия, сагиттария, эхинодорус. Это все водные и болотные растения. А вот в семейство ароидных (Araceae) входит много наземных растений, водные и болотные составляют небольшую часть видов этого семейства. И наоборот, в семейство апоногетоновых (Aponogetaceae) входит только один род апоногетон — все растения водные.
Род объединяет ряд видов. Растения одного рода имеют одно название. Для различения видов К. Линней ввел бинарную систему — к названию рода (существительное) добавляется прилагательное, характеризующее именно данный вид: Cryptocoryne aponogetifolia (криптокорина апоногетонолистная), Aponogeton madagascariensis (апоногетон мадагаскарский).
Но что такое вид? В вид объединяют растения, сходные по морфологическим и физиологическим особенностям, имеющие общее происхождение и занимающие определенную область распространения. Известный советский ботаник В. Л. Комаров говорил: вид — это «морфологическая система, помноженная на географическую определенность». Для определения вида сегодня изучают морфологию, особенности клеточной структуры, генетическую характеристику, физиологию, биохимию растений и т. д.
В то же время вид надо рассматривать не как нечто законченное и уже сформировавшееся, а в динамике, в развитии. «Разновидность, — говаривал Ч. Дарвин, — есть зарождающийся вид». Осваивая новые ареалы с несколько иными условиями, растения могут постепенно приобретать отклонения от исходного вида и образовывать экологические формы, разновидности, или вариации, и подвиды.
Для обозначения таких отклонений от основного вида уже недостаточно бинарной системы, применяется тринарная, например: Cryptocoryne wendtii var. rubella (криптокорина Вендта, вариант (вар.) рубелла) или Cryptocoryne ciliata f. minima (криптокорина цилиата, форма (ф.) минима).
Новые разновидности и виды образуются, путем не только освоения специфических ареалов, но и скрещивания. В природе порой возникают и межвидовые гибриды. Такие гибриды, отмечает К. Ратай, характерны для эхинодорусов*.
* Rataj K. Revision of the Genus Echinodorus Rich. — Praha, Academia, 1975.
Если род распадается на характерные общностные группы, то их называют секцией. Из общего числа апоногетонов с цельнопластинчатыми листьями, обитающих во всех пресных водах Африки, Мадагаскара, Азии и Австралии, решетчатые виды явно выпадают, обособляются. Поэтому их можно выделить в секцию решетчатолистных в этом роде.
Описание нового вида ботаники ведут по эталонному экземпляру, если он представлен в сборе гербария одним растением, или по эталону, выбранному из серии однотипных растений, — по голотипу. Описывающий новое растение ботаник обосновывает выделение его в самостоятельный вид и дает ему название (бинарное) — новым будет только видовое определение. Новые роды выделяются реже, а семейства — в наше время огромная редкость в ботанике высших растений. Так, выделение А. Л. Тахтаджяном в его системе растений самостоятельного семейства барклайевых (при этом сохранено авторство первого ботаника, который такое выделение осуществил) было своего рода ботанической сенсацией.
Когда ученый впервые описывает выделенный им вид, он ставит после бинарного названия «species nov» (вид новый). Такие пометки можно найти около названий криптокорин в книге X. Ц. Д. де Вита, около криптокорин и эхинодорусов — у К. Ратая, апоногетонов — в работах X. В. Е. ван Бруггена. В работах других ботаников упоминание этого же вида будет несколько иным: после бинарного названия идет фамилия ученого, описавшего вид, и дата первой публикации о нем. Для видов, введенных в систему К. Линнеем и сохранивших свои названия, допускается вместо полной фамилии указывать одну заглавную латинскую букву L.
Действительным считается старейшее название, первейший автор. Но порой в последующих изменениях системы меняется название рода, тогда первый автор приводится в скобках. Накопление новых данных, появление новых видов вызывает иногда потребность пересмотреть прежнюю систему. Тогда производится ботаническая ревизия и публикация о ней так и называется: «ревизия рода...», или «ревизия семейства...» Последняя капитальная ревизия семейства апоногетоновых, произведенная X. В. Е. ван Бруггеном, принята всеми ботаниками. Опубликовал ревизии родов эхинодорус и криптокорина К. Ратай. Первая постепенно получила признание многих ботаников, вторая пока вызывает серьезные возражения. Эти три ревизии, естественно, изменили существовавшую до того систему А. Вендта.
Описывая виды водных растений, нам так или иначе придется соприкасаться с определенными системами. Семейства я буду указывать в соответствии с системой А. Л. Тахтаджяна, виды апоногетонов — по X. В. Е. ван Бруггену, криптокорин — по X. Ц. Д. де Виту, эхинодорусов — по К. Ратаю, остальные виды — по А. Вендту.
В бинарном названии на латинском языке родовое всегда пишется с заглавной, а видовое — с маленькой буквы. На русском языке родовое название мы будем писать со строчной буквы, а видовое получит заглавную только тогда, когда название, содержащее имя собственное в определении, пересказывается, так сказать, вольно (криптокорина Вендта). У одного вида может быть несколько названий. Эти названия являются синонимами основного, например: апоногетон мадагаскарский; увирандра; апоногетон фенестралис (если повторяется род основного названия, возможно сокращение — а. фенестралис).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
О питании гидрофитов
О питании гидрофитов «Поскольку гидрофиты никогда не испытывают недостатка в воде, лимитирующим в их развитии становится фактор питания», — замечает Г. Вальтер. Фотосинтез, как мы знаем, происходит на свету. Прежде всего, конечно, за счет использования углекислого газа,
Побываем в гостях у гидрофитов
Побываем в гостях у гидрофитов В природных пресных водоемах, у себя дома, растения развиваются сегодня не изолированно от других живых организмов, а в совокупности с ними. Разные условия среды способствуют образованию различных комплексов живых организмов. Природные
1. ОБЩИЕ СВЕДЕНИЯ
1. ОБЩИЕ СВЕДЕНИЯ Традиционно со времён французского физиолога Биша (начало XIX в.) нервную систему разделяют на соматическую и вегетативную, в каждую из которых входят структуры головного и спинного мозга, называемые центральной нервной системой (ЦНС), а также лежащие вне
6.2.1. Общие сведения
6.2.1. Общие сведения В шерсти различают три типа волос:1. чувствительные (вибриссы),2. остевые (покровные),3. пуховые (подшерсток).Чувствительные волосы самые немногочисленные. Они расположены у собак на губах, подбородке и на бровях. Они выполняют осязательную функцию и,
Когда ученый мир убедился во внеземном происхождении метеоритов?
Когда ученый мир убедился во внеземном происхождении метеоритов? Падения метеоритов люди замечали начиная с самых древних времен, однако не считали их происхождение внеземным. В VIII веке появились две научные (не прибегающих к «божественному промыслу») теории, которые
Гипотеза Пиноккио о происхождении собак
Гипотеза Пиноккио о происхождении собак Широко распространено мнение, что люди создали собак методом искусственного отбора: брали щенков из волчьих логов, приручали и обучали их, использовали для охоты; в итоге через множество поколений такой жизни волки превратились в
Элементарные формы поведения
Элементарные формы поведения Врожденные (генетически обусловленные), имеющие внутреннюю мотивацию формы поведения характеризуются моментом начала проявления в жизни собаки, частотой проявления и моментом его окончания. Например, материнское поведение имеет место у
2. Вопрос о происхождении новых форм
2. Вопрос о происхождении новых форм Концепция Шелдрейка оставляет этот вопрос открытым. Как пишет автор: «Гипотеза формативной причинности есть проверяемая гипотеза об объективно (выделено нами Е. Е.) наблюдаемых закономерностях, существующих в природе. Она не может
Что поведали обезьяны о происхождении государства
Что поведали обезьяны о происхождении государства Теперь, мой Благосклонный читатель, мы с вами уже знаем много такого, что нас так и тянет применить свои познания к собственной истории и к тому, что происходит вокруг нас сейчас. Почему бы нам не оставить на время
Роль языка в происхождении сознания
Роль языка в происхождении сознания Согласно Джулиану Джейнсу (Janes, 1976), единство личности, о котором писал Газзанига, возникло в истории человеческого рода на удивление недавно. Джейнс полагает, что сознание появилось у человека всего лишь около трех тысяч лет назад,
14. Развитие представлений о происхождении жизни на Земле
14. Развитие представлений о происхождении жизни на Земле Вспомните!Что такое жизнь?Назовите основные свойства живого.Вопросы о происхождении жизни на Земле и о возникновении самой Земли всегда волновали человечество. Являясь вечными и глобальными, эти проблемы и
Каждая клетка помнит о своем происхождении
Каждая клетка помнит о своем происхождении Конраду Уоддингтону мы обязаны не только метафорой эпигенетического ландшафта. В 1942 году он стал, как принято считать, крестным отцом понятия «эпигенетика». Слово «эпигенотип» он впервые употребил уже в 1939-м — в своем «Введении
Приложение Б. Нейротрансмиттеры: общие сведения.
Приложение Б. Нейротрансмиттеры: общие сведения. Нейротрансмиттеры — это маленькие молекулы, которые передают информацию через крохотное пространство между нейронами. Они высвобождаются пресинаптическим нейроном в крошечный участок, называемый синаптической щелью,