Глава 13 Постсовременное состояние эволюционной биологии
Глава 13
Постсовременное состояние эволюционной биологии
Пер. А. Самородова
В предыдущих двенадцати главах мы рассмотрели многообразие аспектов эволюции жизни. Подобное рассмотрение, конечно, никоим образом не может быть всеобъемлющим, но это и не было моей целью. Так или иначе, информация, представленная в предыдущих главах, достаточна (и необходима) для выражения сути этой книги: за 50 лет, прошедших со времени утверждения СТЭ в науке, эволюционная биология кардинальным образом изменилась и вошла в новую эпоху — эпоху «постмодерна».
Согласно синтетической теории эволюции, эволюция жизни — это процесс активной адаптации популяций к изменяющимся условиям среды. Теперь стало очевидным, что, хотя подобная адаптация и является, несомненно, существеннейшим компонентом эволюционного процесса, в количественном отношении она не доминирует. И хотя я полностью сознаю, что любая попытка предложить широкое обобщающее определение приводит к чрезмерному упрощению, вместе с тем предлагаю следующее.
Эволюция жизни — это преимущественно стохастический процесс, основанный на исторической случайности, ограниченный прежде всего разнообразными условиями поддержания основ биологической организации и модулируемый механизмом адаптации.
Ограничения, формирующие ход эволюции, имеет смысл рассматривать максимально широко. Они включают все виды предупреждения и устранения повреждений и локальной оптимизации, такие как снижение уровня ошибок во всех информационных процессах и расход энергии, а также непрерывную «гонку вооружений» между паразитами и хозяевами, которая стимулирует эволюцию различных адаптаций, благодаря эффекту Красной Королевы. В этой заключительной главе кратко излагаются разнообразные аспекты постсовременного состояния эволюционной биологии и рассматриваются осуществимость и возможные очертания постсовременной теории эволюции. В табл. 13-1 представлена постсовременная переоценка нескольких фундаментальных положений концепции Дарвина и СТЭ.
Таблица 13-1
Постсовременная переоценка некоторых центральных положений дарвиновской и синтетической теорий эволюции
[Положение] / Постсовременное состояние
[Материал для эволюции поставляется в первую очередь случайными наследуемым и вариациями] Верно лишь отчасти. Репертуар случайных изменений значительно расширен и включает дупликацию генов, областей генома и целых геномов; потерю генов и вообще генетического материала; ГПГ, в том числе массовый перенос генов при эндосимбиозе; вторжение мобильных эгоистичных элементов и пополнение генома их последовательностями и т. п. Более того, (квази)направленное (ламарковское) изменение генома признается существенным фактором эволюции.
[Фиксация (редких) полезных изменений естественным отбором — главная движущая сила эволюции] Верно лишь отчасти. Естественный (положительный) отбор является важным, но лишь одним из нескольких фундаментальных факторов эволюции и не доминирует количественно. Доминирующими в эволюции являются нейтральные процессы в сочетании с очищающим отбором. Также важно прямое воздействие сигналов окружающей среды на геном, то есть (квази)ламарковские явления.
[Изменения, фиксируемые естественным отбором, крайне незначительны. Эволюцию описывает градуализм] Неверно. Дупликация или горизонтальный перенос даже одного гена — отнюдь не «бесконечно малые» изменения, равно как и делеция или приобретение крупных сегментов ДНК, перестановка частей генома, полногеномная дупликация и, что наиболее существенно, эндосимбиоз. Градуализм не может быть основным режимом эволюции.
[Униформизм: эволюционные процессы в целом оставались одними и теми же в течение эволюции жизни] Верно лишь отчасти. Современные эволюционные процессы важны с момента происхождения репликации. Тем не менее крупные скачки эволюции, такие как происхождение эукариот, могут быть вызваны фактически уникальными событиями, такими как эндосимбиоз; самые ранние этапы эволюции (до LUCA) частично опирались на различные процессы, не участвующие в последующей «нормальной» эволюции.
[Эволюция путем естественного отбора стремится производить все более сложные адаптивные свойства организма, а значит, прогресс есть общая тенденция в эволюции] Неверно. Сложность генома, вероятно, эволюционировала как «геномный синдром», вызванный слабостью очищающего отбора в малой популяции, а не как адаптация. Никакой тенденции к увеличению сложности в эволюции нет, и понятие эволюционного прогресса является необоснованным.
[Вся эволюция жизни может быть изображена единым большим древом] Неверно. Открытие фундаментального вклада ГПГ и мобильных генетических элементов в эволюцию генома отвергает понятие ДЖ в его первоначальном смысле. Тем не менее деревья остаются важной формой представления эволюции отдельных генов и многих фаз эволюции в группах относительно близкородственных организмов. Возможность восстановления ДЖ в правах как центральной тенденции эволюции не исключена.
[Все сегодняшние формы клеточной жизни восходят к малому числу древних форм (возможно, одной-единственной, LUCA)] Верно. Сравнительная геномика не оставляет никаких сомнений в общем происхождении клеточной жизни. Однако она также указывает на возможность, что LUCA(S) сильно отличался от современных клеток.
Плюрализм паттернов и процессов эволюции: смена концепций отбора, вариации и древа жизни
Роль и статус отбора
Двоякий смысл слова «постмодерн» в предисловии к этой книге, возможно, не ускользнул от внимательного читателя. Что бы мы ни думали о постмодернистской философии (см. прил. I), ее мировоззрение, безусловно, подчеркивает богатство и чрезвычайное разнообразие процессов и закономерностей, составляющих реальность. Сложность этих разнообразных тенденций такова, что, по мнению некоторых философов постмодернистского толка, ни одно серьезное обобщение не имеет права на существование. В сегодняшней эволюционной биологии разнообразие процессов и закономерностей является, пожалуй, главной темой, и, коль скоро мы принялись говорить парадоксами, можно сказать, что главной темой является отсутствие всеобъемлющей главной темы.
Центр внимания, лежавший в СТЭ на естественном отборе, действующем на случайные генетические изменения, сместился ко множеству взаимодополняющих фундаментальных эволюционных процессов и моделей (см. рис. 13-1). В новой эволюционной биологии естественный отбор является лишь одним из процессов, формирующих эволюционирующий геном, и, видимо, не доминирует количественно. Эволюцию в значительной степени определяют нейтральные процессы, такие как генетические дрейф и тяга.
Развивая тему плюрализма, заметим, что относительный вклад адаптивных и нейтральных процессов отнюдь не постоянен по всей широте спектра форм жизни. Как очень точно сформулировал Майкл Линч (естественно, перефразируя Добржанского), «ничто в эволюции не имеет смысла, кроме как в свете популяционной генетики» (Lynch, 2007b). Действительно, динамика популяций — или, проще говоря, эффективный размер популяции, в краткосрочной и долгосрочной перспективе — является ключевым фактором, определяющим давление отбора. Эффективный размер популяции может отличаться на порядки даже у близкородственных организмов, поэтому различия в интенсивности отбора весьма велики (например, у насекомых и у млекопитающих). Эти различия определяют различные эволюционные режимы: при высоком Ne эволюция определяется в первую очередь (или даже исключительно) отбором; при низких же Ne на первое место выходит дрейф. В реальном процессе эволюции (почти) все линии спуска проходят через несколько «бутылочных горлышек», где доминирующей силой эволюции становится случайный дрейф; отсюда следует неизбежный и большой вклад случайности в эволюцию всех живых организмов.
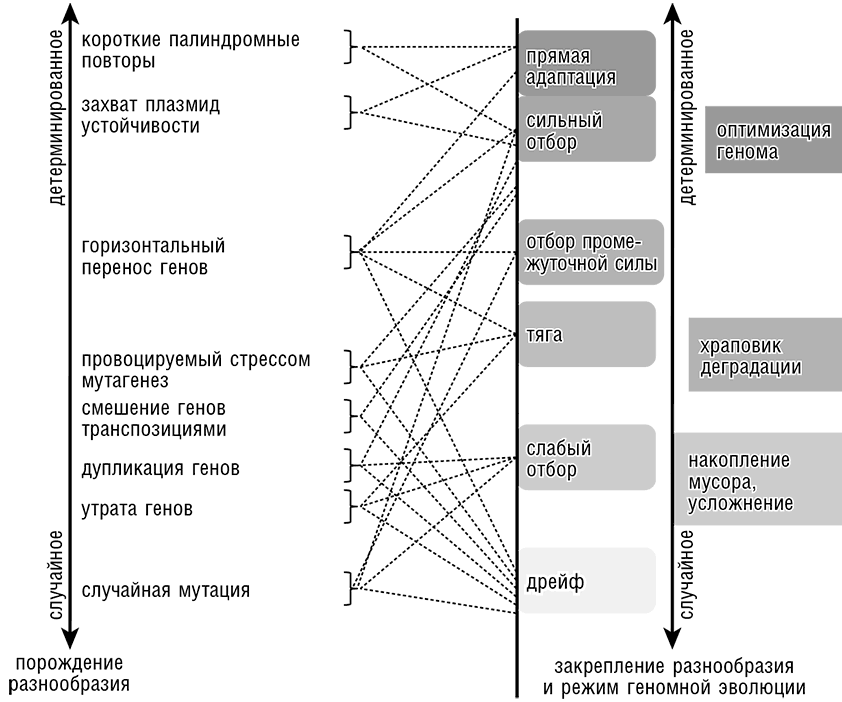
Рис. 13-1. Множественные процессы, порождающие геномное разнообразие, влияющие на его закрепление в эволюции и формирующие эволюцию генома в целом.
В эволюции фенотипов роль отбора, возможно, еще больше, чем в эволюции геномов. Однако достижения системной биологии существенно расширили концепцию фенотипа. Наряду с традиционными признаками организмов, мы теперь изучаем и эволюцию молекулярных фенотипических признаков, таких как экспрессия гена и изобилие белка; молекулярная эволюция фенотипических признаков, как оказывается, включает в себя заметную нейтральную составляющую. Более того, храповик конструктивной нейтральной эволюции, по-видимому, приводит в действие неадаптивную эволюцию сложных фенотипических признаков, выглядящих, с традиционной (нео) — дарвинистской точки зрения, типичными адаптациями.
Даже когда отбор и адаптация несомненно участвуют в процессе, проявление этих факторов эволюции часто (возможно даже, в большинстве случаев) довольно сильно отличается от (нео)дарвиновской идеи «улучшения». Весьма часто адаптации имеют отношение к сохранению целостности клеточной организации, предотвращению сбоев и контролю за повреждениями. В некотором смысле сказанное выше есть тривиальная констатация факта, если мы учтем разнообразие и сложность молекулярных машин, предназначенных для контроля качества каждого из основных процессов передачи информации, примерами которых являются системы репарации ДНК и деградации белков и молекулярные шапероны. Кроме того, важным, чтобы не сказать главным движущим началом эволюции белок-кодирующих генов представляется отбор на устойчивость к ошибкам укладки. В многоклеточных организмах значение отбора для предотвращения сбоев очевидно проявляется на уровне клеток и тканей, примером чего служит сложнейшая система апоптоза, запрограммированной клеточной гибели.
В ретроспективе все эти выводы могут показаться совершенно очевидными, если учесть, как развиты, сложны и в том или ином смысле оптимизированы клетки или даже отдельные белки и РНК. Как только эти сложные системы возникли — а эволюционные реконструкции ясно показывают, что они присутствовали в течение подавляющей части истории жизни, более 3,5 миллиарда лет, — контроль качества и предотвращение повреждений действительно стали основными «задачами» эволюции, невзирая на важность появления время от времени новых адаптаций. Признание этого факта возлагает огромную нагрузку на ранние, доклеточные стадии эволюции, когда изменения были быстрыми, а роль положительного отбора, наряду с конструктивной нейтральной эволюцией, должна была быть куда значительнее, чем в течение последующих 3,5 миллиарда лет. В некотором смысле почти все «действительно интересное» в эволюции жизни произошло на сравнительно короткой ранней стадии, предшествующей установлению основ клеточной организации (см. гл. 11 и 12, а также более подробно ниже в данной главе). Конечно, существуют и заметные исключения, такие как появление эукариотических клеток или многоклеточных эукариотических организмов, но нет никаких сомнений, что большинство фундаментальных эволюционных инноваций «втиснуто» в короткий отрезок первых 5 процентов истории жизни.
Меняющиеся концепции вариации и конец градуализма
В дополнение к меняющимся концепциям отбора и его роли в эволюции были радикально переработаны и идеи о том, что является эволюционно важными геномными и фенотипическими вариациями (см. рис. 13-1). Случайные изменения, ведущие к бесконечно малым полезным фенотипическим изменениям, которые Дарвин рассматривал как ключ ко всей эволюции, остаются важными, но становятся лишь одним из классов изменений, существенных для эволюции, притом не доминирующим — во всяком случае, количественно.
Начнем с того, что почти нейтральные мутации, «невидимые» отбору и не закрепленные дрейфом или же сохраняющиеся в популяции, не будучи закрепленными, встречаются, по-видимому, чаще, чем слегка полезные «дарвиновские» мутации. Важный момент, который не был четко осознан до недавнего времени, состоит в том, что практически нейтральные мутации далеко не безразличны для эволюции. В действительности эти мутации образуют нейтральные сети, составляющие важнейший резервуар эволюционной пластичности.
Кроме того, формы генетических вариаций, которые ни в коем случае нельзя рассматривать как ведущие к бесконечно малым полезным эффектам, имеют решающее значение для эволюции. Это понимание ставит крест на градуализме, который Дарвин и архитекторы СТЭ полагали центральным для всей эволюции. Неградуалистские типы эволюционно важных генетических изменений включают дупликации генов и целых геномов, потерю генов и ГПГ, в частности храповики обширного, направленного переноса генов, провоцируемого эндосимбиозом. Горизонтальный перенос генов является наиболее распространенным эволюционным процессом у прокариот, и, по всей видимости, имеют место специфические адаптации, поддерживающие ГПГ на оптимальном уровне. Скорость ГПГ значительно уменьшается у эукариот, где обширная дупликация с последующей субфункционализацией компенсирует сокращение ГПГ. Кроме того, эндосимбиоз и последующий перенос генов от симбионтов к хозяину сыграли решающую роль в эволюции эукариот.
Не только бесконечно малый эффект изменений, но и его полная случайность также уходит в прошлое. Такие механизмы, как стресс-индуцированный мутагенез, высокоразвитые, сложные системы которого существуют во всех клеточных формах жизни, являются адаптивными и неслучайными, и адаптивная система иммунитета у прокариот, по-видимому, действует через самое настоящее ламарковское наследование. Как правило, эволюционные процессы охватывают континуум ламарковского, дарвиновского и райтовского режимов эволюции, и относительный вклад каждого из них в любом конкретном эпизоде из истории жизни зависит от динамики популяции и давления окружающей среды. Наконец, становится все более ясным, что фенотипические мутации существенны для эволюции и могут способствовать адаптации в сочетании с генетическими мутациями, в частности посредством эффекта предвидения. Все это вместе — важность неслучайных мутаций и (квази)ламарковские механизмы эволюции, наряду с вкладом фенотипических мутаций, — показывает, что способность к эволюции тоже эволюционирует, что, в свою очередь, по-видимому, опровергает, хотя бы частично, одно из заветных убеждений эволюционных биологов — веру в то, что эволюция не имеет предвидения.
От древа жизни к паутине жизни
Приверженность эволюционной биологии древу жизни как единому, окончательному представлению об истории форм жизни на Земле уступает место плюралистической картине, в которой разнообразные сетевые процессы дополняют древовидные процессы генной эволюции. Эти процессы включают ГПГ, особенно распространенный у прокариот, но также вносящий огромный вклад в эволюцию эукариот, особенно через эндосимбиоз, а также через различные формы слияния генома и обмена генетическим материалом между хозяевами и паразитами (см. гл. 5 и 7). В первом приближении можно сказать, что в наших представлениях об истории жизни древо жизни уступило место паутине жизни (или ризоме жизни; Raoult, 2010)[137].
Когда мы глубже всматриваемся в эволюционные процессы, становится ясно, что эволюция жизни может быть осмысленно отражена лишь сложной, динамической картиной взаимодействующих процессов, среди которых древовидная эволюция будет, вероятно, самым фундаментальным, поскольку эта форма эволюции является прямым следствием двоичной репликации генетического материала как базового универсального процесса всей жизни (гл. 6). Более того, имеется неоспоримая динамическая согласованность между эволюционной историей больших ансамблей генов (самые большие из таких коэволюционирующих ансамблей известны как геномы), возрождающая концепцию ДЖ, но уже в виде центральной статистической тенденции в «лесу» филогенетических деревьев отдельных генов. Несмотря на статистическую согласованность генетических историй, «ядро» универсально консервативных, повсеместно распространенных генов клеточных форм жизни чрезвычайно мало из-за потери генов в отдельных линиях эволюции и неортологичного замещения генов — фундаментальных эволюционных явлений, важность которых не могла быть оценена в догеномную эпоху. Суммируя, можно сказать, что паутина эволюции представляет собой высокодинамичное геномное пространство-время, в котором геном каждого вида есть лишь преходящее, метастабильное скопление генов.
Неожиданная значимость простых физических и математических моделей для понимания эволюции: биологическая эволюция как предмет статистической физики
В предыдущем разделе мы рассмотрели множественность структур и процессов, которые являются определяющим аспектом новой концепции эволюции. Упрощенно говоря, эта множественность значительно увеличивает энтропию эволюционной биологии. Тем не менее анализ данных, получаемых в областях геномики и системной биологии, выявляет в равной степени заметную противоположную, «антиэнтропийную» тенденцию к структурированию эволюционной теории. Было обнаружено несколько универсальных распределений и зависимостей, таких как распределение скорости генной эволюции, связь между эволюцией и экспрессией генов и распределение степени связности узлов разнообразных сетей. Более того, некоторые из этих универсалий могут быть легко получены из простых математических моделей эволюции, очень похожих на модели, используемые в статистической физике. Эти модели становятся все более общими по мере того, как они объединяются и совместно объясняют универсальные зависимости, первоначально казавшиеся несвязанными, например распределение скорости генной эволюции совместно с антикорреляцией между эволюционной скоростью и экспрессией, или законы масштабирования для семейств генов совместно с масштабированием по функциональным классам.
Имеется удивительно простое общее объяснение этой неожиданной применимости простых моделей для объяснения эволюции геномов. Эволюционная геномика имеет дело с большими ансамблями объектов (генов, белков), которые можно для многих целей рассматривать как слабо взаимодействующие и перемещающиеся (то есть эволюционирующие) по независимым траекториям. Соответственно, принципы статистической физики столь же применимы к генетическим ансамблям, сколь и к ансамблям молекул. Естественно, статистический подход к эволюционным явлениям подвержен тем же ограничениям, что и аналогичные подходы в физике, а именно эти модели недостаточны для объяснения конкретных биологических явлений, часто связанных с небольшим набором генов, а не с большим ансамблем. Кроме того, взаимодействие между генами, эпистаз, часто вносит существенный вклад в эволюцию[138]. Тем не менее примечательно, что современная геномика и системная биология, раскрывая чрезвычайно сложную, многогранную картину эволюции, в то же время позволяют выработать и простые обобщенные модели. Очень заманчиво предложить еще один новый вариант знаменитой фразы: ничто в эволюции — и в популяционной генетике — не имеет смысла, кроме как в свете статистической физики.
Воспроизводимость эволюции: детерминизм и стохастика эволюционного процесса
Пространство генотипов, даже если рассматривать только относительно простые, небольшие геномы, невообразимо огромно (скажем, для прокариот с геномом в 1 миллион пар оснований имеется 41 000 000 возможных последовательностей, число, значительно превосходящее по величине все, что существует на самом деле в наблюдаемой части Вселенной, например общее число протонов или электронов). Какая часть этих генотипов на самом деле жизнеспособна и, следовательно, могла бы сыграть роль в эволюции? Или, чтобы задать вопрос осмысленным в контексте эволюции образом, какова доля всех возможных траекторий в пространстве генотипов, которые открыты эволюционному процессу? Вышесказанное — формализованная постановка любимого вопроса Стивена Джей Гулда (Gould, 1997b): что бы мы увидели, если бы имели возможность заново проиграть пластинку эволюции? Ответ, данный не только Гулдом, но и Франсуа Жакобом в знаменитой статье об эволюции-«ремесленнике» (Jacob, 1977), Дэном Деннетом в «Опасной идее Дарвина» (Dennett, 1996) и многими другими, был таков: мы бы не увидели ничего подобного реально существующей биосфере, потому что вся эволюция — сплошная цепь непредвиденных стечений обстоятельств. При описании общей картины эволюции Деннет вполне обоснованно обращается к физическому явлению детерминированного хаоса: каждое событие, которое происходит в процессе эволюции, безусловно, имеет конкретные физические причины, но малые возмущения способны вызвать большие изменения в ходе эволюции, так что далекие результаты становятся совершенно непредсказуемыми.
По-прежнему трудно дать уверенный общий ответ на этот ключевой вопрос эволюции, однако имеющиеся ограниченные прямые исследования эволюционных траекторий как индивидуальных белков, так и бактериальных популяций принесли неожиданные результаты (O’Maille et al., 2008; Ostrowski et al., 2008; Weinreich et al., 2006). Похоже, что в большинстве случаев лишь небольшая часть из теоретически возможных путей на самом деле доступна для эволюции, так что эволюция представляется менее стохастической, более детерминированной и более предсказуемой, чем предполагалось ранее (см. рис. 13-2). Эти наблюдения позволяют предположить, что адаптивные ландшафты по меньшей мере некоторых из развивающиеся генов и геномов являются существенно неровными, так что большинство путей прерывается глубокими оврагами низкой приспособленности и, таким образом, запрещены (O’Maille et al., 2008). Основной подоплекой этого, вероятно, является эпистаз, то есть взаимодействие между различными частями одного и того же гена или между различными генами: на пересеченном ландшафте одна мутация часто приводит к фатальному падению приспособленности, но вторая, путем эпистаза, способна привести в область высокой приспособленности. Эпистаз представляется одним из важнейших факторов, поддерживающих целостность эволюционирующих биологических систем, которая проявляется в многих аспектах эволюции (Kogenaru et al., 2009). Как отмечалось в предыдущем разделе, эпистаз неизбежно ограничивает применимость представления эволюционирующих геномов ансамблями слабо взаимодействующих «частиц». Эпистатическое взаимодействие сильно ограничивает диапазон доступных эволюционных траекторий — но насколько сильно, еще предстоит выяснить с помощью дальнейшего моделирования и экспериментальных исследований эволюции. Вполне может оказаться, что модель детерминированного хаоса верна и что обнаруженные ограничения на практике мало влияют на предсказуемость эволюции, то есть на результат метафорического повторного проигрывания пластинки. Доступные траектории, даже если они и составляют лишь малую долю теоретически возможных, все же могут оказаться столь многочисленными и разнообразными, что эволюция окажется на практике непредсказуемой. Важнейшей и пока нерешенной проблемой оказывается взаимосвязь между доступными траекториями. Если эти траектории кластеризуются на небольшом участке геномного пространства-времени, эволюция может быть квазидетерминированной[139]; если же доступные траектории беспорядочно разбросаны, (не)предсказуемость эволюции не будет сильно зависеть от подобных ограничений (см. рис. 13-2).
Скорее всего, результаты детального анализа эволюционных ландшафтов и траекторий на них будут различаться для эволюции на различных уровнях и в различных ситуациях, в согласии с концепцией плюрализма эволюционных процессов, обсуждавшейся выше. Кроме того, следует еще раз подчеркнуть, что соотношение между детерминизмом и стохастичностью определяется давлением отбора, то есть эффективным размером популяции. В эффективно бесконечной популяции эволюция фактически детерминирована, в то время как в небольших популяциях эволюция происходит стохастически в рамках фундаментальных ограничений. Чтобы исключить всякую возможность недоразумений, отметим, что, даже если эволюция и может быть описана как квазидетерминированный процесс, такое описание не имеет ничего общего с телеологическими представлениями. Тем не менее канализация в смысле Уоддингтона (см. гл. 2) представляется интересной аналогией.
Сложное и неоднозначное соответствие генома и фенотипа
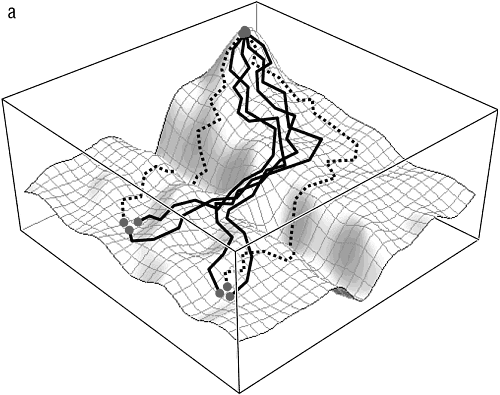
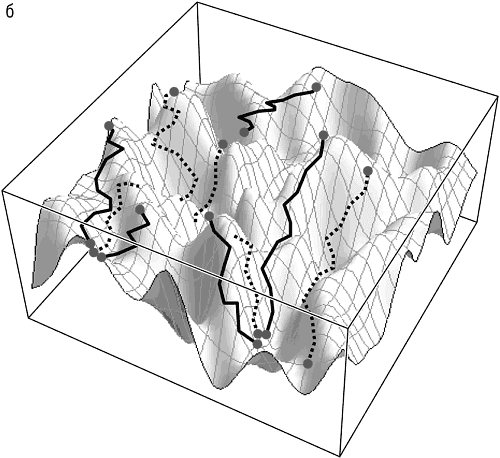
Рис. 13-2. Пересеченный адаптивный ландшафт и доступные эволюционные траектории: а — квазидетерминированная эволюция: канализация доступных траекторий; б — стохастическая эволюция: случайное распределение доступных траекторий. Сплошные линии — монотонно восходящие траектории, доступные для эволюции, движимой исключительно отбором. Пунктирные линии — немонотонные траектории, доступные только с участием генетического дрейфа.
Принято считать, что геном (генотип) определяет фенотип организма (с некоторым участием эпигенеза), фенотип жестко контролируется отбором, а фенотипические изменения не имеют эволюционных последствий. Сравнительная геномика и системная биология показывают, что все эти утверждения не являются истиной в последней инстанции, и такие упрощенные обобщения оставляют в стороне ключевые биологические явления. Отсутствие простой детерминированной связи между фенотипом и генотипом выражается по крайней мере в двух взаимодополнительных аспектах их взаимоотношений:
1. Фенотипические мутации и другие формы шума, такие как случайная транскрипция практически всех геномных последовательностей у животных, неотъемлемо присущи биологическим системам и вносят вклад в их эволюцию (см. гл. 9)[140]. Эти эволюционно важные фенотипические изменения частично контролируются геномом, но связь между геномом и шумом стохастическая по своей природе.
2. Отображение генома на фенотип является неизоморфным и сложным (говоря проще, это многозначное отображение, то есть не отображение одного элемента в один, но многих в многие); все гены плейотропны, и все фенотипические свойства («функции» или «антрвольты») зависят от активности многих генов. В целом геномфенотипическое отображение является исключительно сложным графом (см. гл. 5, в основном рис. 5-9). Ребра этого графа имеют различные веса, которые отображают разный вклад разных генов в один и тот же признак.
Повсеместное распространение и эволюционное значение фенотипической изменчивости делают связь генома с фенотипом принципиально недетерминированной. Многозначное отображение ограничивает эволюцию, возможно являясь даже ee главным ограничителем (см. предыдущий раздел), но при этом делает связь генома с фенотипом невероятно сложной, a реконструкцию фенотипа по геному крайне затруднительной. Некоторые простые фенотипические признаки, конечно, предсказуемы. Например, если бактерия не имеет лактозного оперона, она не сможет употреблять лактозу. Тем не менее даже такие простые признаки часто могут реализовываться несколькими разными путями. Любой сложный фенотип крайне трудно поддается предсказанию, например термофилия и устойчивость к радиации у прокариот, как мы видели в главе 5. Сложность связи генома с фенотипом и, как следствие, трудность построения надежных представлений о фенотипе организма только на основании анализа генома резко усугубляется для эукариотических и особенно многоклеточных организмов. Удивительное и труднообъяснимое, но на данный момент несомненное отсутствие сильной связи между очевидной биологической важностью генов и скоростью их эволюции (см. гл. 4) особенно подчеркивает возникающее понимание того, что фенотипические последствия эволюции генома нетривиальны и, в общем случае, трудно предсказуемы. Недооценка этой сложности может привести к нереалистичным надеждам на быстрый успех в проектах, направленных на исследование и манипуляции со сложными фенотипами, таких как изучение полногеномных ассоциаций, «война с раком» или индивидуализированные лекарства[141].
Заря экспериментальной эволюционной биологии
Эта книга прежде всего о концепциях, идеях и моделях, а не о методах. Тем не менее, прежде чем закончить эту последнюю главу, необходимо сказать несколько слов о новом поколении подходов, которые не только позволили поразительно глубоко проникнуть в ключевые эволюционные процессы, но и должны начать менять сам облик эволюционной биологии в следующем десятилетии (или близко к тому). Эти исследовательские стратегии подпадают под категорию «экспериментальной эволюционистики». В сегодняшних эволюционных экспериментах ход эволюции популяции организмов или молекул можно проследить непосредственно, обрабатывая, за счет применения нового поколения методов секвенирования, тысячи и даже миллионы молекул ДНК или РНК. Эксперименты Ричарда Ленски и его коллег по долгосрочной лабораторной эволюции популяций E. coli, о которых мы не раз говорили в этой книге, являются замечательным примером такого рода опытов (Ostrowski et al., 2008; Barrick et al., 2009; Woods et al., 2011). Эти эксперименты уже дали бесценную информацию о различных режимах отбора и дрейфа, распространенности параллельных мутаций, эволюции способности к эволюции и многом другом. Однако, благодаря появившейся в настоящее время возможности секвенирования тысяч полных бактериальных геномов, самые многообещающие результаты ожидаются в не столь отдаленном будущем, когда будут всесторонне изучены эволюционные траектории популяций в различных условиях окружающей среды и при разных давлениях отбора. Концептуально эти эксперименты продолжают направление исследований, начатое в пророческой работе Шпигельмана и коллег с РНК-бактериофагами в 1960-х годах (см. гл. 8). Эксперименты Шпигельмана почти на полвека опередили свое время и оказали сравнительно небольшое влияние на развитие науки, но в первые десятилетия XXI века статус экспериментального исследования эволюции быстро меняется.
Другое направление эволюционного экспериментирования включает в себя изучение ландшафтов отбора эволюционирующих белков или молекул РНК, которые мы кратко обсудили ранее в этой главе. Имеющиеся в настоящее время экспериментальные данные описывают только крошечные доли ландшафтов, но возможность исследовать большие области уже представляется реалистичной (Kogenaru et al., 2009; Loewe, 2009)[142]. В конечном итоге подробная реконструкция полных ландшафтов отбора изменит наши представления о том, что значит «понимать» эволюционный процесс.
Дивный новый мир вирусов и прокариот
СТЭ ориентируется в основном на изучение эволюции животных и растений — многоклеточных эукариот, размножающихся чаще всего половым путем. Одноклеточные эукариоты и прокариоты, не говоря уж о вирусах, не считались важными для эволюционной биологии. Пожалуй, включение огромного мира микробов в систему эволюционных представлений следует считать главным толчком для перехода от СТЭ к «постсовременному» состоянию эволюционной биологии. Первые попытки разобраться в эволюционных связях между бактериями были в высшей степени обескураживающими, но последующее сравнение последовательностей консервативных генов таких как рибосомные РНК, а затем и полный анализ геномов привели к замечательным успехам. Сравнительная геномика бактерий и архей изменила центральные концепции эволюционной биологии, включая древо жизни (см. гл. 6), и показала крайне динамичный характер геномов и пангеномов (см. гл. 5).
Изучение мира вирусов привело к не менее, а может быть, и более драматическому изменению наших взглядов на эволюцию жизни на Земле. Крошечные паразиты, вирусы являются наиболее многочисленными и генетически разнообразными биологическими объектами на всей планете. Мир вирусов, по всей вероятности, существует с самых ранних, доклеточных стадий эволюции и постоянно взаимодействует с миром клеточных организмов, внося существенный вклад в его эволюцию, но сохраняя при этом собственную автономию.
Империи и домены жизни
В свое время Карл Вёзе, используя древо рРНК, классифицировал живое на три домена, что являлось огромным концептуальным скачком как для эволюционной биологии, так и биологии в целом. Положение вещей, однако, радикально изменилось за последние 30 лет, и в рамках этой классификации живых организмов не могут быть объяснены новые сложные реалии эволюции, открытые сравнительной геномикой. Первым важным открытием, противоречащим трехдоменной схеме, была демонстрация химерной природы генома эукариот. Трехдоменное древо отображает эволюцию только подмножества генов, вовлеченных в информационные процессы и количественно составляющих незначительное меньшинство эукариотических генов, даже внутри группы высококонсервативных генов, прослеженных до последнего общего предка всех эукариот. Конечно, доменная классификация живых организмов является лишь соглашением среди исследователей, поэтому классификация эукариот как отдельного домена не является правильной или неправильной сама по себе. Тем не менее такая классификация способна ввести в заблуждение, особенно в сопровождении тройственной схемы древа жизни, поскольку объединение двух организмов и их изначально отдельных геномов, которое, по всей видимости, дало начало эукариотам, в этой схеме не отражено. Схема же, включающая такое объединение в явном виде, с математической точки зрения деревом не является (см. рис. 13-3).
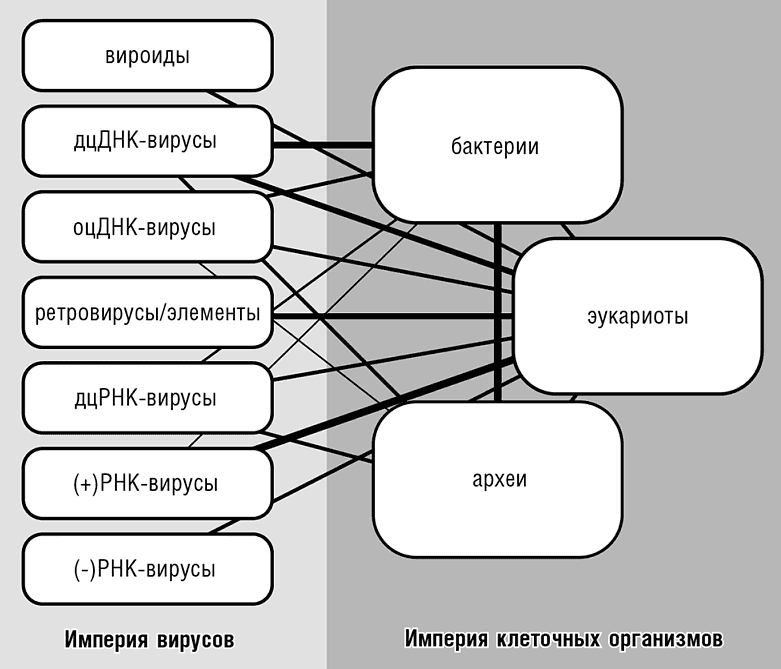
Рис. 13-3. Империи и домены жизни. Соединительные линии показывают потоки генетической информации между доменами, включая как вирусные инфекции, так и все пути горизонтального переноса генов.
Возможно, даже более фундаментальным отходом от трехдоменной схемы стало открытие мира вирусов с его неожиданным и ошеломляющим простором и удивительной эволюционной связностью[143]. Как обсуждалось в главе 10, вирусоподобные паразиты неизбежно возникают в любой репликаторной системе, таким образом, не будет преувеличением утверждение, что жизнь невозможна без присутствия вирусов. Более того, кажется почти неизбежным, что доклеточная эволюция жизни прошла через вирусоподобную стадию развития. Также важно заметить, что главные группы вирусов раличаются между собой не меньше (а возможно, и больше), чем три (или два) домена клеточных форм жизни. Эти различия обусловлены тем, что разные группы вирусов используют различные циклы репликации/экспрессии, в отличие от клеточных организмов, которые в этом отношении устроены идентично (подробности см. в гл. 10).
Любая классификация по сути всего лишь соглашение, в то время как эволюционные сценарии стремятся реконструировать, по возможности, историю, которая на самом деле произошла. Очевидно, наилучшей классификацией живых организмов будет та, что наиболее точно отражает лучший из существующих эволюционных сценариев. С этой точки зрения наибольший смысл имеет разделение всех известных форм жизни на две «империи»[144]: вирусов и клеточных форм (Koonin, 2010d). Эти две империи сильно отличаются друг от друга, но постоянно обмениваются генами (см. рис. 13-3). Можно говорить о трех или только о двух доменах клеточной жизни, но архео-бактериальное объединение должно быть необходимой частью классификационной схемы. Несмотря на то что нашей целью не является высказывать какие-либо формальные предложения относительно новых «вирусных доменов», отдельные большие группы вирусов, характеризующиеся общими наборами консервативных генов, несомненно, сравнимы по своему статусу с доменами клеточной жизни.
Парадокс биологической сложности, обманчивость прогресса и значение неадаптивных храповиков
Клетки и организмы как устройства для самовоспроизведения генов
По наблюдениям автора этой книги, многие исследователи и просто образованные читатели с пренебрежением относятся к концепции эгоистичного гена Ричарда Докинза (Dawkins, 2006) — во всяком случае, в ее крайних формах — возможно, потому, что она противоречит здравому смыслу и «унижает наше достоинство». Тем не менее, если мы принимаем тезис о том, что репликация генетического материала — основное свойство живых систем (см. гл. 2), у нас нет логического выхода, кроме как принять и концепцию эгоистичного гена. В частности, Докинз весьма решительно называет организмы транспортными средствами для репликации и эволюции генов, и я думаю, что эта простая идея описывает ключевой аспект биологической эволюции. Конечно, это не подразумевает никакого метафизического или телеологического контекста, например будто клетки и организмы существуют «с целью» репликации генов. «Целеориентированный» подход вообще неконструктивен (см. прил. I). Таким образом, предположение, будто фенотип существует с целью репликации, столь же бессмысленно, как и противоположное ему. Тем не менее между геномом и фенотипом существует фундаментальная, логически неизбежная асимметрия: все фенотипические признаки организмов, как и сами клетки и организмы, рассматриваемые как сложные физические системы, возникают и эволюционируют лишь ввиду того, что они способствуют процессу самовоспроизведения генома — либо увеличивая эффективность процесса, либо, по крайней мере, не уменьшая ее.
Как обсуждалось в главе 10, «сердцевину» фенотипа всех организмов в значительной мере составляют антиэнтропийные устройства, уменьшающие уровень ошибок при передаче информации во время самого процесса репликации и подчиненных ему процессов транскрипции, трансляции, укладки РНК и белков, а также снижающие вредные эффекты таких ошибок, когда они все-таки случаются. В остальном большая часть «сердцевины» фенотипа — это вспомогательные устройства, которые производят и транспортируют строительные блоки для репликации и сохранения фенотипа. Таким образом, невозможно отрицать, что эволюция фенотипа направлена на обеспечение репликации генов (генома).
С точки зрения одной лишь репликации, возникновение сложности являет собою загадку: почему существует так много живых форм, неизмеримо более сложных, чем это необходимо для минимального, простейшего приспособления для репликации? Мы не знаем точно, каким должно быть самое простое устройство для самовоспроизведения, но у нас есть замечательные кандидаты на это звание — простейшие автотрофные бактерии и археи, такие как Pelagibacter ubique или Prochlorococcus sp. Эти организмы обходятся приблизительно 1300 генами, не потребляют органических молекул и полностью независимы от других живых форм[145]. Кстати, эти организмы также являются наиболее «успешными» на Земле. Они имеют самые большие популяции, которые эволюционировали под сильнейшим давлением отбора и в результате этого обладают наиболее «оптимизированными» геномами. Биосфера, состоящая из одних таких высокоэффективных одноклеточных организмов, легко представима, более того, земная биота до появления эукариот (то есть первые 2 миллиарда лет эволюции или около того) должна была быть очень похожа на такой мир, гораздо более, чем сегодняшняя биосфера (хотя более сложные прокариоты, несомненно, существовали и в то время).
Так почему же организмы столь сложно устроены?
Один из ответов, казалось бы, самоочевидный и принимавшийся биологами и всеми интересующимися эволюцией, состоит в том, что более сложные организмы являются также более приспособленными. Эта точка зрения, несомненно, ошибочна. Более того, и в этом состоит парадокс сложности, общая тенденция прямо противоположна: чем сложнее организм, тем меньший эффективный размер популяции он имеет и, по единственно разумному определению эволюционного успеха, тем менее успешным он является. Понимание этого может подсказать нам, что решение загадки появления сложности может быть поразительно простым: взглянув на ту же тенденцию с противоположной стороны, мы заметим, что чем меньше эффективный размер популяции, тем ниже интенсивность отбора и, следовательно, больше вероятность неадаптивного развития сложности. В этом и состоит суть популяционно-генетической неадаптивной концепции эволюции генома, предложенной Линчем.
Теперь мы можем сформулировать более конкретный ответ на вопрос в заголовке этого раздела — или, скорее, несколько взаимодополняющих ответов.
1. В самом общем смысле сложность появляется просто «потому, что может»: принимая во внимание универсальный процесс «беспорядочного блуждания», при наличии достаточного времени, стабильно возрастает вероятность случайного усложнения биологической организации.
2. Другое, более конкретное описание роли случайности проявления как определяющего фактора эволюции дается популяционной генетикой: сложность может возникнуть в результате случайной фиксации фактически нейтральных (слегка вредных) мутаций через генетический дрейф в популяциях с малым эффективным размером. Таким образом, из комбинации (1) и (2) получается, что сложность возникает, «потому что может», при условии слабого давления очищающего отбора, который не может отбраковать незначительно вредные изменения, такие как дупликация генов, включение мобильных элементов в нескольких сайтах генома и т. д.
3. На фоне этих случайных факторов эволюция сложной организации стала возможной благодаря храповикам конструктивной нейтральной эволюции: как только два или более гена делаются зависимыми друг от друга, в результате дифференциального накопления слегка вредных мутаций в обоих, оба они фиксируются в эволюции и оказываются неразрывно связанными, что приводит к повышенной сложности. Таков механизм эволюции генетических дупликаций, а также многих горизонтально перенесенных генов при субфункционализации; по-видимому, это один из основных путей эволюции.
4. Сложные формы обычно не более приспособлены, чем простые, однако сложность может облегчить адаптацию к новым нишам, как, например, в случае наземных растений. Появление сложности, таким образом, может нести в себе и адаптивную компоненту, в дополнение к основным неадаптивным факторам, упомянутым ранее.
Необходимо подчеркнуть существенную роль нейтральных храповиков в придании эволюции очевидной направленности без фактического повышения приспособленности. Храповик конструктивной нейтральной эволюции может быть ключевым элементом в появлении разнообразных сложных биологических свойств: храповики переноса гена от эндосимбионта к хозяину внесли существенный вклад в эукариогенез, а храповики необратимой потери генов явились ведущим фактором в редуктивной эволюции паразитов и симбионтов. Храповики можно рассматривать как узкие, крутые горные хребты на адаптивном ландшафте: как только эволюционирующая популяция оказывается на таком гребне, она начинает следовать квазидетерминированному пути, поскольку падение с хребта приводит к резкому снижению приспособленности и неизбежному вымиранию. В этом процессе сложность может возникать без вклада адаптации.
Величайшая загадка происхождения жизни
Благодаря достижениям в областях геномики и системной биологии, за первое десятилетие XXI века мы накопили много больше знаний о ключевых аспектах эволюции, чем за предыдущие полтора века. Хоть крупные скачки эволюции, такие как происхождение эукариот или происхождение животных, остаются чрезвычайно сложными проблемами, появляется все больше и больше зацепок для их решения. Вне всякого сомнения, был достигнут значительный прогресс даже в этих трудных областях эволюционной биологии[146]. Мы даже начали разрабатывать сценарии возникновения клетки, выходящие за рамки чистых спекуляций.
Тем не менее происхождение жизни, или, если быть более точным, происхождение первых репликаторных систем и происхождение трансляции, остается великой загадкой, и прогресс в решении этих проблем очень скромен — и даже, если говорить о трансляции, незначителен. Существует несколько потенциально плодотворных наблюдений и идей, таких как открытие вероятных инкубаторов жизни — сетей неорганических ячеек в гидротермальных источниках — и химическая многосторонность рибозимов, поддерживающая гипотезу мира РНК. Однако эти достижения остаются только предварительными, пусть и важными, поскольку мы даже близко не подошли к убедительному сценарию добиологической эволюции от первых органических молекул до первой репликаторной системы и от них до собственно биологических объектов, в которых хранение информации и функция разделены между различными классами молекул (нуклеиновых кислот и белков соответственно).
На мой взгляд, несмотря на все достижения, эволюционная биология является и будет оставаться крайне неполной, пока нет хотя бы правдоподобного, пусть не полностью убедительного, сценария происхождения жизни. Поиск решения этой великой загадки может вести нас в неожиданных (и глубоко противоречащих, с точки зрения биологов, здравому смыслу) направлениях, в частности к полной переоценке важнейших понятий случайности, вероятности и возможного вклада чрезвычайно редких событий, что может быть проиллюстрировано на примере космологического подхода, рассматриваемого в главе 12.
Нужна ли и полезна ли новая теория биологической эволюции? Появится ли постсовременная синтетическая теория?
Принцип дополнительности Бора, как представляется автору этой книги, занимает центральное место в нашем понимании эволюции, прежде всего в смысле взаимной дополнительности между случайностью и детерминированными факторами (необходимостью) — той самой дополнительности, которая является лейтмотивом этой книги. Дополнительность проявляется на всех уровнях и во всех аспектах эволюции и может стать основным руководящим принципом на пути к новой теоретической биологии. Очевидные случаи, которые мы обсудили в этой книге, включают дополнительность между:
• случайными и (квази)направленными мутациями;
• отбором и дрейфом;
• эгоистичным и альтруистичным поведением разнообразных генетических элементов (персистентных вирусов, ретроэлементов, систем токсин — антитоксин и рестрикции — модификации и др.);
• устойчивостью и способностью к эволюции.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава 1. О прогрессивном развитии в биологии
Глава 1. О прогрессивном развитии в биологии Мы, разумные существа, не должны забывать, что наша цивилизация — лишь одно из замечательных явлений природы, зависящих от постоянного притока концентрированной энергии солнечного излучения. Ю. Одум Вопрос о движущих силах
Глава 7. Похвала биологии
Глава 7. Похвала биологии Каждый из нас, к сожалению, хорошо знаком с неприятным явлением, которое все еще бытует кое-где, несмотря на строгие приказы министерств торговли, культуры и Госкомиздата СССР. Речь идет о так называемой «продаже с нагрузкой». Согласитесь, крайне
Глава 3 Рождение современной биологии
Глава 3 Рождение современной биологии Новая анатомияЗнаменующим началом научной революции принято считать 1543 г. В этом году польский астроном Николай Коперник опубликовал книгу, где была изложена новая точка зрения на Солнечную систему, центром которой было Солнце,
Введение На пути к новому синтезу эволюционной биологии[1]
Введение На пути к новому синтезу эволюционной биологии[1] Пер. А. ЧеркасовойНазвание настоящей работы связано с четырьмя замечательными книгами: романом Пола Остера «Музыка случая» (Auster, 1991), знаменитым трактатом Жака Моно по молекулярной биологии, эволюции и философии
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции Пер. А. НестеровойВ этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись
Глава 23. Обмен аминокислот. Динамическое состояние белков организма
Глава 23. Обмен аминокислот. Динамическое состояние белков организма Значение аминокислот для организма в первую очередь заключается в том, что они используются для синтеза белков, метаболизм которых занимает особое место в процессах обмена веществ между организмом и
Глава вторая СОСТОЯНИЕ СОБАК ПРИ 7-, 12- И 19-ЧАСОВОМ ПРЕБЫВАНИИ В КАМЕРЕ
Глава вторая СОСТОЯНИЕ СОБАК ПРИ 7-, 12- И 19-ЧАСОВОМ ПРЕБЫВАНИИ В КАМЕРЕ Влияние на собак 7-часового пребывания в камере Вегетативные показатели: после трех-четырехразового пребывания в 7-часовых опытах наблюдалось снижение пульса (на 5–11 ударов в минуту), числа
Глава третья СОСТОЯНИЕ СОБАК ПРИ ПЕРВОМ СПОСОБЕ ТРЕНИРОВКИ
Глава третья СОСТОЯНИЕ СОБАК ПРИ ПЕРВОМ СПОСОБЕ ТРЕНИРОВКИ Первый способ тренировки являлся окончательным этапом 7-, 12- и 19-часовых опытов. Животные после этих экспериментов фиксировались в камере на 15–20 суток и, в отличие от предыдущих этапов лишенные прогулок,
Глава четвертая СОСТОЯНИЕ СОБАК ПРИ ВТОРОМ СПОСОБЕ ТРЕНИРОВКИ
Глава четвертая СОСТОЯНИЕ СОБАК ПРИ ВТОРОМ СПОСОБЕ ТРЕНИРОВКИ Большой процент суток опытов с аномальным ритмом выделений (57,3%) от общего числа суток, возникновение других неблагоприятных сдвигов, тот факт, что только одна собака оказалась по-настоящему пригодной к
Глава пятая СОСТОЯНИЕ СОБАК ПРИ ДВУХМЕСЯЧНОЙ ФИКСАЦИИ
Глава пятая СОСТОЯНИЕ СОБАК ПРИ ДВУХМЕСЯЧНОЙ ФИКСАЦИИ В начале двухмесячных опытов состояние собак, несмотря на многие индивидуальные отклонения, изменялось по закономерностям, отмеченным при 20-суточных
Глава 1. Нерешенные проблемы биологии
Глава 1. Нерешенные проблемы биологии 1.1. Предпосылки успеха Цель механистического подхода в биологическом исследовании особенно ясно была выражена более ста лет назад Томасом Хаксли в следующем определении:«Зоологическая физиология — это доктрина функций или
Глава вторая ДНК Рационализация биологии
Глава вторая ДНК Рационализация биологии Почти все проявления жизни заложены на молекулярном уровне, и без понимания молекул мы можем иметь лишь весьма поверхностное понимание самой жизни. Френсис Крик Великая идея: наследственность закодирована в ДНККаждый из нас
Глава 5 Современное состояние биосферы Земли
Глава 5 Современное состояние биосферы Земли 5.1. Границы биосферы Биосфера является одним из трех (гидросфера, атмосфера и литосфера) компонентов климатической системы. Ее можно уподобить тонкой пленке, покрывающей поверхность нашей планеты. Плотность органического
Глава 10 Состояние почв и лесов Приморья
Глава 10 Состояние почв и лесов Приморья 10.1. Общая характеристика По данным разнообразных исследований, доминирующими загрязнителями атмосферы являются автомобильный транспорт, предприятия электроэнергетики и многочисленные котельные. Из основных веществ в списке
Глава I. Основы мичуринской биологии Кандидат сельскохозяйственных наук Е. К. Меркурьева
Глава I. Основы мичуринской биологии Кандидат сельскохозяйственных наук Е. К. Меркурьева 1. Понятие о биологии Биология — это наука о законах развития живой природы. Она изучает жизнь как животных, так и растений. Основная задача биологии состоит в том, что она должна
Глава 12. Гомосексуализм в эволюционной перспективе
Глава 12. Гомосексуализм в эволюционной перспективе Гомосексуализм как культурное явление Эволюционная теория предполагает, что адаптивные признаки должны воспроизводиться более успешно, чем неадаптивные. Важным индикатором адаптивности признака является его