Глава вторая ДНК Рационализация биологии
Глава вторая
ДНК
Рационализация биологии
Почти все проявления жизни заложены на молекулярном уровне, и без понимания молекул мы можем иметь лишь весьма поверхностное понимание самой жизни.
Френсис Крик
Великая идея: наследственность закодирована в ДНК
Каждый из нас состоит почти из сотни триллионов самих себя. Каждая из наших клеток — а их примерно сто триллионов, и большинство их так малы, что понадобится около двух сотен, чтобы покрыть точку над i — содержит полную схему нашего тела. В принципе (всегда угрожающе подозрительный оборот речи) ваше тело, рассыпанное на свои сто триллионов клеток, могло бы породить сто триллионов вас, каждый из этих новых вас, рассыпанный снова, мог бы стать еще одной сотней триллионов, и вы с вашими клонами очень быстро достигли бы абсолютного доминирования во Вселенной. К счастью, имеются физические и биологические ограничения, делающие невозможным воплощение этой фантазии. Но даже рассмотрение такой возможности заставляет предположить, что мы в беспрецедентной степени осведомлены о клеточной природе жизни.
Мы осведомлены. Дарвин и его современники, возможно, за исключением одного монаха, ничего не знали о природе наследственности. Несмотря на то что они проницательно смотрели на мир природы и хорошо осознавали результаты конкуренции, крылья их понимания были подрезаны неосведомленностью о механизмах наследования. Наиболее почитаемым механизмом в то время была смешанная наследственность, при которой каждый из родителей сливает свои наследуемые характеристики в общий котел, которому предстоит стать их ребенком, и ребенок возникает из этой смеси. Тот факт, что подобное смешивание не смогло бы поддерживать естественный отбор, поскольку новые адаптационные свойства быстро размывались бы, использовался как сильный аргумент против точки зрения Дарвина и задерживал полное принятие его теорий. Аристотель, хотя и был замечательным искателем вопросов, давал обычно неверные ответы, снова и снова демонстрируя бесполезность размышлений в кресле, не подкрепленных экспериментом.[5] Замечая, что кровь омывает все органы тела, Аристотель назначил носителем наследственности кровь, и этот взгляд сохранился до сих пор в качестве метафоры. Он считал семя очищенной кровью, которая при копуляции смешивается с менструальной кровью и порождает следующее поколение.
Монахом, владевшим ключом, конечно, был Грегог Мендель (1822-84), названный Иоганном при рождении в семье фермера в Хайнцендорфе, на севере Моравии, провинции в Австрийской Силезии, позднее вошедшей в Чехословакию, ныне Чешскую Республику. Отец Менделя, Антон, был незадачливым малым, чье здоровье и средства к существованию были подорваны ботаникой в виде упавшего на него дерева. Антон продал ферму своему племяннику, так что смог вносить плату за сына, которому предстояло посвятить жизнь ботанике в школе в Троппау и затем в университете в Олмютце. Единственным способом получить дешевое образование для Менделя было поступление в монастырь Св. Томаша в Брюнне (теперь Брно) в возрасте двадцати двух лет, где он принял имя Грегор и был возведен в сан священника в 1847 г. Шагом, подготовившим его ум к малой арифметике наследственности, которую ему суждено было разработать позже, было направление в Вену для изучения наук и математики, чтобы стать учителем. Однако его успехи в учении были незначительны, особенно в биологии, и через два года он возвращается в монастырь, чтобы позднее стать его аббатом (1868).
Мендель был священником прихода в прелатстве имперского и королевского Австрийского ордена императора Франца-Иосифа, заслуживающим похвал директором Моравского ипотечного банка, основателем Австрийской метеорологической ассоциации, членом Моравского и Силезского общества поддержки агрокультуры, естественных наук и краеведения, и, что наиболее важно, садовником. В 1850-х гг., примерно в то же время, когда Дарвин записывал свои мысли, он начал исследования, сделавшие его посмертно знаменитым. Множество вопросов о достоверности его работы или работы его ассистентов поднималось — и энергично отводилось, — когда выдающийся статистик и генетик Рональд Элмер Фишер (1890-1962) объявил, что цифры, приводимые Менделем, вызывают подозрения. Позднее понимались вопросы, знал ли действительно Мендель, что он делает, и не является ли миф, выросший вокруг его достижений, скорее следствием нашей подслеповатости, чем его прозрения. Так, толчком к работам Менделя послужило желание понять скорее правила гибридизации, чем механизм наследственности. Мотивацией же была попытка реализовать преобладающую в то время точку зрения, что новые виды возникают из гибридизации, причем «устойчивые» гибриды и становятся новыми видами. Его безрассудной целью было создание новых видов, и он потерпел в этом оглушительное фиаско.
Мендель представил свои результаты — по существу, мрачный отчет о неудаче — в собрании общества естествоиспытателей Брюнна на заседаниях 8 февраля и 8 марта 1865 г. и опубликовал их как «Опыты над растительными гибридами» (Versuche ?ber Planzenhybriden) в трудах общества в 1866 г. Эти результаты были полностью проигнорированы, если не считать вводящей в заблуждение цитаты в Die Pflanzen Mischlinge (1881) В.О. Фока, и пролежали незамеченными до 1900 г. Возможно, они были проигнорированы потому, что с современной им точки зрения описывали лишь неудачу в попытке выявить рациональные основы гибридизации, а дрейф Менделя в сторону административной работы также мог отражать его собственное разочарование в печальном исходе трудов всей его жизни. Затем три ботаника — Хуго де Врис в Голландии, Карл Эрих Корренс в Германии и Эрих Чермак фон Зейсенегг в Австрии — обнаружили, как они заявили, что сами того не зная, как они заявили, повторили его работу. В этих сообщениях имеется специфический привкус надувательства, поскольку было согласовано, что один из авторов (де Врис) отложит признание приоритета Менделя до той поры, когда окажется, что другой (Корренс) уже опубликовал подобную работу, так что де Врис, понимая, что так или иначе первенство придется уступить, объявил приоритет Менделя попыткой замутить блеск сообщения Корренса. Вся манера объяснений была выдержана в духе пренебрежения к работе Менделя тридцатипятилетней давности, и содержала утверждения, что он был вторгшимся в науку любителем, что он был слишком тесно связан с церковью, от которой ничего хорошего ждать не приходится, что его математические построения — даже в простой арифметике, которая ему требовалась — приводят в замешательство современных биологов. Истина может быть проще: до тех пор, пока де Врис, Корренс и фон Зейсенегг не вытащили на свет его работу и не взглянули на нее с более современной точки зрения, о механизме наследственности не появилось ни одной стоящей мысли.
Хотя Мендель провел свои исследования в девятнадцатом веке, их значение стало очевидным только в двадцатом. Теперь мы понимаем, что Мендель квантовал наследственность, подобно тому, как Планк квантовал энергию (см. главу 7). Теперь мы способны увидеть, что его достижением были свидетельства, которые привели к низвержению преобладающей тогда теории смешанной наследственности и к ее неизбежно последовавшей замене на теорию, в которой наследственную информацию несли дискретные единицы. В течение восьми лет его внимание было сфокусировано на садовом горохе (Pisum sativum), обладавшем множеством свойств, необходимых для проводимых им исследований. Во-первых, сама структура цветка довольно специфична и дает возможность либо легко скрестить два растения, либо, как это случается в природе, позволить им самоопылиться. Более того, это растение имеет ряд изменчивых характеристик: например, его лепестки могут быть белыми или пурпурными, его горошины могут быть округлыми или покрытыми морщинами, иметь зеленую или желтую внутренность, находиться в желтом или зеленом стручке, а его ростки могут быть кряжистыми или тонкими. Более того, и, возможно это было подлинной причиной, горошины имели достаточно низкую цену на рынке семян, занимали мало места и давали много ростков в относительно короткое время. Мы также можем подозревать, что гороховый суп удручающе часто появлялся в меню монастыря Св. Томаша. Единственным недостатком садового гороха было то, что он не слишком фотогеничен в пейзаже, и экспериментальный садик Менделя засадили, к удовольствию посетителей, более привлекательными бегониями (рис. 2.1).

Рис. 2.1. Сад Менделя в его монастыре. Мендель использовал в своей работе обычный горох, что оказалось удачным выбором, отчасти из соображений экономии, но также и потому, что многие характеристики гороха генетически независимы. В настоящее время этот сад засажен бегониями.
Мендель хотел знать, каким способом гибридизация декоративных растений производит изменения, повторяющиеся в следующих поколениях. Он решил поискать систематическую схему, которая, как он считал, могла скрыто присутствовать в наблюдениях. В первые два года он решил убедиться, что его растения дают правильное потомство, что кустики зеленого гороха порождают зеленый горох, а кустики желтого гороха порождают желтый, и то же происходит с другими признаками. Потом он начал серию перекрестных опылений и самоопылений. Например, когда он скрещивал зеленый горох с желтым, весь горох в потомстве первого поколения (в так называемых F1 гибридах) был желтым. Однако, когда гибриды самоопылялись, три четверти гороха в следующем, F2, поколении были желтыми, а одна четверть зеленой. Таинственным образом первоначальный зеленый опять появился. Подобная схема, с тем же численным отношением, возникала, когда он скрещивал и потом самоопылял растения, проявляющие другие характеристики. Ясно, что схема проявилась, а схемы вопиют, требуя объяснения.
На основании огромного числа наблюдений Мендель построил гипотезу. Первым ключом для него стал тот факт, что его эксперименты приводят к вариантам с простыми числовыми отношениями. Чтобы найти объяснение дискретным числам, которые получались в этих отношениях, он предположил, что различие внутри каждой пары характеристик (зеленый и желтый горох, например) обусловлено присутствием в растении различных дискретных единиц. Мендель использовал термин «элемент», чтобы обозначить дискретные целостности наследственности, и употреблял термин «характер», когда обсуждал внешний вид, фенотип своих растений. Большинство его рассуждений проводилось в терминах этих наблюдаемых характеров, и только более поздние интерпретаторы обратили внимание на роль лежащих в основании «элементов». Эти целостности тогда получали множество различных наименований, но теперь повсеместно известны под именем, которое предложил в 1909 г. датский биолог Вильгельм Людвиг Иогансен, гены. Более точно, различные версии генов, ответственные за частные фенотипы, например, ответственные за цвет гороха, называются аллелями. Так, зеленый горох и желтый горох соответствуют разным аллелям гена, ответственного за цвет гороха.
Чтобы объяснить простые числовые отношения, установленные Менделем, предположим, что гены — мы будем использовать современный термин — существуют парами, причем каждому характеру соответствует одна пара, и что каждая гамета (яйцеклетка и сперма у животных, семяпочка и пыльца у растений) содержит один из этих генов. Тогда при зачатии (опылении у растений) мужская и женская гаметы соединяются случайно и объединяют индивидуальные гены обратно в пары. Мендель разделил наследуемые характеристики на доминантные и рецессивные, и задним числом мы можем видеть, что это разделение приложимо также и к генам. Поэтому, если доминантный аллель объединится в пару с рецессивным, фенотип проявит характеристики доминантного аллеля. Например, эксперименты Менделя показывают, что аллель желтого гороха является доминантным по отношению к аллелю зеленого гороха, поскольку при скрещивании дающего правильное потомство желтого растения с дающим правильное потомство зеленым растением все потомки являются желтыми.
Можно проиллюстрировать эти идеи символически. Обозначим аллель желтого гороха буквой Y, а рецессивный аллель зеленого гороха буквой у (в элементарной генетике есть соглашение: доминантный аллель обозначается буквой, указывающей на соответствующее свойство, в данном случае на английское слово yellow, желтый, а его рецессивный двойник такой же, но маленькой, буквой). Дающие правильное потомство желтый и зеленый горох обозначаются соответственно как YY и yy. Гаметы каждого растения обозначаются соответственно как Y и y. Когда их скрещивают, потомство должно быть Yy, и весь горох будет желтым, потому что желтый (Y) доминантен. Теперь самоопылим эти гибриды. Поскольку гамета растения Yy может случайным образом оказаться Y или y, потомки растений Yy будут четырех видов: YY, Yy, yY и yy. Только последний из них, yy, соответствует зеленому гороху (поскольку Y доминантен в Yy и yY), так что растения являются желтыми и зелеными в отношении 3:1, в точности как и наблюдал Мендель. Он сумел распространить эту простую схему на другие характеристики и их комбинации (зеленый и карликовый горох, к примеру) и в каждом случае обнаружил, что ожидаемые отношения подтверждаются. (Именно здесь Фишер подверг его статистику атаке, поскольку отношения не были точными, а разброс результатов — который мог возникнуть из-за систематической ошибки, сдвига в желаемую сторону, при решении вопроса, является ли горошина со слегка неровной поверхностью гладкой или морщинистой — вызывал подозрения.)
Не всякая наследственность является менделевской, в смысле подчинения законам Менделя с простой статистикой. Возможно, наихудший совет в истории экспертных советов был дан немецким ботаником Карлом Вильгельмом фон Нэгели из Мюнхенского университета, который не понял аргументов Менделя и предложил ему переключить свое внимание с гороха на ястребинку (Hieracium). Но ястребинка размножается путем соматического партеногенеза (т.е. неполовым путем), и едва ли есть что-либо менее подходящее для демонстрации менделевской наследственности. Мендель, должно быть, несколько приуныл, когда его опыты с ястребинкой привели в никуда и определенно не подтверждали его идеи. Он также был подавлен результатами опытов с бобами (Phaseolus), в которых так много генов отвечают за характеристики, которые он наблюдал, что ожидаемые им простые отношения, такие ясные для гороха Pisum, оказались скрытыми.
Существуют и более тонкие причины, по которым не вся половая наследственность является менделевской, так как некоторые гены сцеплены с другими, и наследование определенных пар характеристик не является случайным. Более того, многие гены плейотропны, в том смысле, что они управляют более чем одной чертой фенотипа, и организм не является взаимно однозначным отображением характерных черт в гены. Например, мутация фруктовой мушки Drosophila, героини многих генетических штудий, приводит к недостатку пигментации ее сложных глаз и ее почек (Malpighian tubules); в другой мутации не только крылья вытягиваются в стороны, но мушка также теряет несколько волосков по бокам. Даже статистика правильной менделевской наследственности может затеняться вторичными эффектами. Например, бесхвостая кошка имеет ген, назовем его t, который мешает нормальному развитию позвоночника у Tt кошек и дает в результате знакомый бесхвостый фенотип; но если дать кошке двойную дозу этого аллеля, она становится нежизнеспособной, эмбрионы tt умирают. «Самоопыляющиеся» Tt кошки дадут поэтому в потомстве, способном к появлению на свет TT, Tt и tT в отношении 1:2, вместо ожидаемого 1:3.[6]
Эта работа отдыхала тридцать пять лет, пока ее не откопали и с неохотой признали при, возможно, несколько темных обстоятельствах, о которых мы упомянули выше. Но пока наблюдения Менделя крепко спали, биология путешествовала по другой дороге, которой суждено было раствориться в них.
Заслуживающий цитирования немецкий биолог Эрнст Геккель (1834-1919) придумал для нас термин филогения, означающий эволюционную историю вида, и предположил, что «филогения повторяет онтологию», где онтология есть развитие индивида. Он имел в виду, что превращения, которым подвергается эмбрион при развитии в матке, являются ускоренной версией эволюции вида. Он также сделал предположение, имевшее чудовищные последствия через двадцать лет после его смерти, что политика представляет собой прикладную биологию. Более уместно для текущего обсуждения предположение, сделанное им в 1868 г., о том, что ядра биологических клеток содержат информацию, которая управляет наследственностью. Немецкий эмбриолог Вальтер Флеминг дал новый импульс этому предположению, когда в 1882 г. обнаружил, что ядра клеток личинки саламандры содержат крошечные стержнеподобные структуры, которые могут окрашиваться путем поглощения определенных красителей. Основываясь На этих наблюдениях, Вильгельм фон Вальдейер в 1889 г. предложил название хромосома («окрашенное тело»).[7]
Число хромосом в ядрах клеток, как известно, трудно сосчитать, поскольку они сплетаются, расплетаются и расползаются по ядру, пока оно не подвергнется делению, и тогда они начинают удваиваться и делиться. Животные, которых мы считаем малыми, а заодно и растения, обычно имеют меньше хромосом, чем мы: у нас их двадцать три пары, а у домовой мыши только двенадцать. Томаты, однако, имеют двадцать две, а картофель, к нашему стыду, двадцать четыре. И действительно, подсчет так труден, что долгое время число хромосом у человека считали таким же, как у шимпанзе (двадцать четыре пары); и только проглотив свою гордыню и признав, что число хромосом не связано с самоуверенным восхищением собой, мы смогли принять правильное число, двадцать три.[8]
На рубеже веков биологи стали подозревать, что хромосомы являются инструментом наследственности. Эти хромосомы зашагали в ногу с менделевской наследственностью в 1902 г., когда Уолтер Саттон (1877-1916), выпускник и сотрудник Колумбийского университета в Нью-Йорке, изучавший сперму кузнечиков (а именно, большого равнинного кузнечика, Brachystola magna, который встречается повсеместно на равнинах запада Соединенных Штатов и Мексики и имеет большие клетки и сносно видимые хромосомы), обнаружил, что парные хромосомы действительно делятся, причем разные члены каждой пары попадают в разные клетки. Открытие Саттона обычно называют теорией Саттона-Бовери, поскольку Теодор Бовери (1862-1915), немецкий биолог, работавший над яичниками морских ежей, объявил в 1904 г., что он тоже пришел к этой мысли, и как раз в то же время, что и Саттон. Бовери действительно внес (наряду с другими) несколько центральных идей, но более важно, что у него были влиятельные друзья.
На этой стадии мы можем заключить, что менделевские гены заключены в саттоновских хромосомах. Мир был подготовлен к новой науке, и в 1905 г. немного странноватый Уильям Бейтсон предложил термин «генетика», сначала в письме к кембриджскому зоологу Адаму Седвику, а затем, в 1906 г., публично на третьей международной конференции по гибридизации. О тяжеловесности его стиля, а возможно, и о высотах, которых достигла связь науки с общественностью за сто лет, можно судить по его замечанию,
что этот термин в достаточной мере указывает на то, что наши труды посвящены разъяснению феномена наследственности и изменчивости: другими словами, физиологии наследования, со всеми вытекающими из нее аспектами теоретических проблем эволюционистов и систематиков, и приложениями к практическим проблемам размножения, будь то животных или растений.
Прежде чем сделать следующий шаг в генетику и внутренний мир наследственности, нам необходимо узнать, что включают в себя два решающих процесса: митоз, деление соматических клеток (обычных клеток тела), и мейоз, образование гамет (спермы и яйцеклетки, пыльцы и семяпочки) в гонадах (половых органах) животных и в пыльниковых мешочках и завязях растений. Сложность последнего процесса является одной из причин, объясняющих, почему эволюция полового размножения так трудна для понимания и почему она была столь грандиозным эволюционным подарком (глава 1). Тем не менее перед Природой возникла задача, и мейоз — а это логически гораздо более сложная задача, чем митоз — появляется там и тогда, где и когда он необходим. Тут не учебник биологии, поэтому я приведу лишь эскиз этих двух процессов, подробный настолько, насколько это необходимо для понимания дальнейшего.
Сначала рассмотрим митоз, копирование соматических клеток. Жизнь клетки циклична, и лишь около десяти процентов ее времени отведено митозу. Остальное время, однако, критически важно, поскольку на его протяжении приготовляются многие вещества, которые будут использованы в акте копирования. Большую часть этого лежащего под паром, но плодородного, времени все двадцать три пары наших хромосом вытягиваются и сложным образом распределяются по ядру клетки. При наступлении митоза (рис. 2.2) хромосомы стягиваются в спирали, становясь более подготовленными к движению в разных направлениях. На этой стадии становится также видно, что каждая хромосома уже подверглась копированию, поскольку она уже состоит из двух идентичных стержнеподобных единиц, называемых хроматидами, соединенных вместе областями, называемыми центромериями, принимая облик, похожий на вытянутое X. Затем оболочка ядра расходится, и компоненты ядра вместе с окружающей цитоплазмой, сложной смесью составов и структур, находящихся внутри стенок клетки, но вне ядра, сливаются в одно. Хроматиды теперь растаскиваются в стороны, и между двумя отрядами хромосом (которыми мы теперь считаем разделившиеся хроматиды) начинает формироваться клеточная мембрана, новая мембрана ядра начинает возникать вокруг каждой копии, спирали хромосом разворачиваются, и мы получаем уже две идентичных клетки вместо одной.

Рис. 2.2. Процесс митоза, деление соматической клетки на две копии. Первоначально хромосомы распределены по всему ядру (изображаемому здесь в виде внутренней сферы). Когда деление начинается, хромосомы свертываются в спирали, удваиваются и образуют протяженные объекты в форме буквы X (здесь мы показываем лишь два из них; в клетке человека имеются двадцать три таких пары), состоящие из двух хроматид, соединенных центромериями. Хромосомы располагаются в линию на центральной плоскости, мембрана ядра разжижается, хромосомы разделяются и по отдельности выталкиваются в цитоплазму клетки. Затем мембрана ядра преобразуется, а мембрана клетки начинает закрываться вокруг каждого из новых ядер. Наконец, хромосомы раскручиваются, и мы получаем две идентичные диплоидные клетки (клетки со спаренными хромосомами) там, где первоначально была одна.
Теперь рассмотрим мейоз, образование гамет. Этот процесс гораздо более тонок, чем митоз, поскольку конечным выходом в нем должно быть формирование четырех клеток, каждая с одной половиной от пары хромосом (которых у человека двадцать три). Этот процесс является довольно сложным, поэтому давайте проследим его шаги на рис. 2.3, где мы сосредоточились на паре хромосом. Первоначально хромосомы сплетены вместе и заполняют ядро, но при начале мейоза они расплетаются и сжимаются. На этой стадии через микроскоп становится видно, что каждая хромосома удвоилась и состоит из двух хроматид, соединенных центромериями в форме обычного вытянутого X, в точности как при митозе. Теперь, однако, пара материнских и пара отцовских хроматид движутся вместе и формируют продолговатый объект, похожий на две стороны застежки-молнии. Каждая хромосома прикрепляется к оболочке ядра своими концами, которые называются теломерами («удаленными частями»); такая постановка на якорь, возможно, помогает одной стороне «молнии» найти своего партнера. Пока две удвоенные хромосомы лежат вместе, вещество в хроматиде, представляющей отцовскую составляющую, заменяется на вещество соответствующей области хроматиды, предоставленной матерью. Это мгновение, когда в организме происходит генетическое изменение.
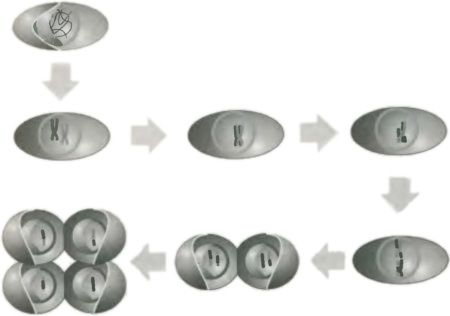
Рис. 2.3. Процесс мейоза, образования гамет. Стратегией мейоза является превращение диплоидной клетки в четыре гаплоидных клетки (клетки с одиночными версиями хромосом) и создание генетической композиции родительских хромосом. Мы снова показываем лишь одну пару хромосом в родительской клетке. Первоначально две хромосомы распределены по всему ядру. Однако, когда начинается мейоз, они свертываются в спирали и удваиваются, чтобы образовать две пары соединенных между собой хроматид, так же как при митозе. Однако соответствующие пары сопряженных хроматид перемещаются вместе и, находясь по соседству, обмениваются генетическим материалом. Затем они мигрируют к центральной плоскости, где происходит первое деление, подобное происходящему при митозе (в деталях мы его не показываем) и дающее в результате две клетки с двумя хромосомами в каждой. Затем следует второе митотическое деление, в котором две хромосомы каждого ядра разделяются снова. Процесс оканчивается появлением четырех гаплоидных клеток, каждая из которых содержит хромосому, представляющую собой генетическую смесь двух хромосом клеток родителей. Воспроизведение теперь, на понятийном уровне, но не механистически, является обращением мейоза, в котором одна хромосома в гамете, предоставленной одним из родителей, соединяется с другой хромосомой, предоставленной другим родителем.
После этого временного затора в истории организма, процесса кроссинговера (взаимного обмена между парами хромосом), две пары гибридных хромосом растаскиваются по двум областям — что довольно похоже на митоз, — чтобы образовать две клетки, каждая из которых содержит пару хроматид. Это «первое митотическое деление» на иллюстрации. Затем во «втором митотическом делении» каждая из пар хроматид растаскивается на индивидуальные хромосомы, которые теперь занимают индивидуальные клетки. В этой конечной точке процесса у нас оказалось четыре клетки там, где была одна, а исходный генетический материал от обоих родителей распределился по всем четырем клеткам. Хромосомы одной из этих клеток могут содержать доминантный аллель Y гена желтого гороха; в другой может находиться рецессивный аллель y зеленого гороха. Арифметика Менделя уже почти готова войти в его сад. Обратим здесь внимание на еще одну грань науки: за простотой арифметических наблюдений может лежать огромной глубины сложность, в нашем случае сложность биологической клетки.
Теперь пришло время развернуть хромосому. Что на самом деле является веществом наследственности? Каково физическое воплощение генетической информации?
Мысль о том, что наследуемая информация кодируется химически, возникала уже в девятнадцатом веке, ибо где же еще ей в конце концов находиться? Примерно с 1902 г. и была принята точка зрения, что белки представляют собой нитеподобные молекулы (обычно свернутые в шарики), построенные из набора примерно двадцати аминокислот в определенной последовательности (подробнее об этом мы скажем ниже), и возник всеобщий энтузиазм по поводу идеи о том, что генетическая информация закодирована в белках, и различные последовательности аминокислот передают различные послания от одного поколения другому. Удивляло, однако, загадочное присутствие в клеточных ядрах молекул другого типа, названного, чтобы отметить его происхождение из ядра, «нуклеиновой кислотой». Они состоят из нитей, в которые входят единицы другого типа, о них речь пойдет позже. Эти нуклеиновые кислоты находили скучными и структурно слишком простыми для того, чтобы переносить огромное количество информации, содержащейся в хромосомах. Было широко распространено предположение, что они просто входят в структуру клетки, подобно тому как целлюлоза входит в структуру растений.
Эту точку зрения пришлось переменить в 1944 г. Биохимик, игравший на корнет-а-пистоне, Освальд Эвери (1877-1955), родившийся в семье британских иммигрантов в Новой Шотландии (Канада), но сделавший свою основополагающую работу в Соединенных Штатах, исследовал различные типы пневмококков, находящихся в полости рта у пациентов, больных пневмонией, и у здоровых людей. С 1923 г. было известно, что пневмококки (бактерии, вызывающие пневмонию) появляются в нескольких разновидностях: невирулентные (незаразные) формы выглядят неровными, в то время как вирулентные штаммы выглядят гладкими. Фредерик Гриффите (1879-1941), работавший в Министерстве здравоохранения в Лондоне над Streptococcus pneumoniae, показал, что неровные и гладкие формы могут быть превращены друг в друга. Эвери и его коллеги принялись за работу в 1930 г. и вскоре обнаружили, что трансформация одного типа бактерий в другой может быть получена в экстракте из клеток и что «источник трансформации», являющийся ее эффективным агентом, может быть выделен. Эвери затем сосредоточил усилия на выяснении природы источника трансформации. Он обнаружил, что протеаза, которая является ферментом, дезактивирующим белки, не влияет на активность источника, так что источник не является белком. Он обнаружил также, что липаза, которая является ферментом, разрушающим липиды, жировые субстанции, составляющие стенки клетки, также не дает эффекта, поэтому источник не является липидом. Выяснив, какие вещества не являются источником трансформации, Эвери продолжил серию опытов, и они показали, что источником была старая, скучная нуклеиновая кислота. Это смешало все карты, и нуклеиновые кислоты встали на путь карьерного роста, как Кларк Кент на путь Супермена, чтобы вдруг оказаться самыми интересными и важными молекулами в мире.
Не всех удалось убедить. Некоторые очень привязались к белковой теории наследования и настаивали в своих публикациях, что источником трансформации является, возможно, еще не выявленный белок, ассоциированный с нуклеиновой кислотой. Эта точка зрения была решительно отвергнута в последующие несколько лет. В 1952 г. Альфред Херши (1908-97) и его ассистент, студентка последнего курса Марта Чейз обнародовали результаты своих опытов на бактериофагах, вирусах инфицирующих бактерий. Они обнаружили, что элементарный фосфор присутствует в нуклеиновых кислотах, но отсутствует в белках, а сера присутствует в белках, но отсутствует в нуклеиновых кислотах. Затем, используя радиоактивные версии каждого элемента, они проследили их путь и показали, что в процессе инфицирования в клетку бактерии попадает только нуклеиновая кислота фага, а не его белок. Этот эксперимент убедил научный мир в том, что наследуемая информация закодирована в нуклеиновой кислоте.
Тем временем был достигнут прогресс в изучении структуры одной из нуклеиновых кислот, дезоксирибонуклеиновой кислоты (ДНК). Это соединение было обнаружено в 1868 г. шведским врачом Фредериком Мишером в немецком городе Тюбингене в клетках из пропитанных гноем повязок, снятых с раненых солдат. Гной представляет собой в основном скопление белых кровяных телец, которые накапливаются для борьбы с инфекцией; хотя красные кровяные тельца млекопитающих не имеют ядер, у белых они есть, и они явились источником нуклеиновых кислот.
Чтобы понять все, что за этим последовало, нам нужно кое-что узнать о химическом устройстве ДНК. Лучше всего сделать это, разложив на части ее полное наименование, дезоксирибонуклеиновая кислота. Эта молекула подобна длинной нити, к которой регулярно по всей ее длине прикреплены другие молекулы. Сама нить построена попеременно из молекул сахара и фосфатных групп. Молекулой сахара является рибоза, близкая родственница глюкозы, из которой удален один атом кислорода (отсюда части «дезокси» и «рибо» в названии). Как можно видеть на рис. 2.4, рибоза состоит из простого кольца, содержащего четыре атома углерода и один атом кислорода, и всяких кусочков, прикрепленных к кольцу. Фосфатные группы, связывающие вместе кольца дезоксирибозы состоят из атома фосфора (вспомним опыты Херши), к которому прикреплены четыре атома кислорода. Позвоночником для ДНК служит чередование фосфатных и дезоксирибозных групп, достигающее сотен и тысяч повторений, подобное хрупкой жемчужной нити.
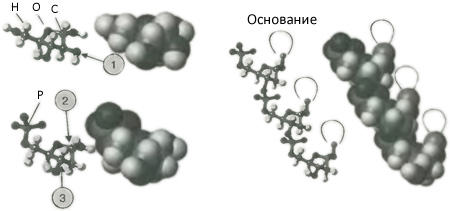
Рис. 2.4. Структура дезоксирибонуклеиновой кислоты (ДНК). Мы можем понять структуру этой сложной молекулы, взглянув, как она слагается из простых компонент. Слева вверху мы видим молекулу сахара рибозы. Эта молекула является кольцом из четырех атомов углерода (C) и одного атома кислорода (O), с различными частицами и кусочками, прикрепленными к нему. Теперь вообразите, что один атом кислорода, прикрепленный к атому углерода на юго-востоке кольца (стрелка 1), удален, чтобы получилась дезоксирибоза, а к другому концу молекулы прицеплена фосфатная группа. Теперь представьте, что молекулярная группа — нуклеотидное основание (смотрите на рис. 2.5, но представляйте комочек здесь) — прикреплена к одному из углеродных атомов кольца (стрелка 2), а фосфатная группа связана с другим углеродным атомом кольца (стрелка 3), чтобы образовать цепь, как показано справа. Это цепь ДНК.
Это позвоночник. К каждому кольцу дезоксирибозы прикреплены другие молекулы, называемые нуклеотидными основаниями. Термин «основание» в этом названии имеет техническое происхождение, так как в химии основными называют соединения, вступающие в реакции с кислотой: основными эти соединения делает присутствие атомов азота в их молекулах, верный признак основных соединений в химии. В ДНК встречаются только четыре нуклеотидных основания, а именно, аденин (обычно обозначаемый буквой A), гуанин (G), цитозин (C) и тимин (T). Аденин и гуанин имеют во многом похожие формы, с двумя углеродными кольцами и атомами азота, сцепленными вместе. Эти структуры характерны для класса соединений, которые химики называют «пуринами». Напротив, цитозин и тимин имеют лишь одно углеродное кольцо с атомами азота. Эти структуры характерны для класса соединений, называемых «пиримидинами». Чтобы вообразить молекулу ДНК, представьте себе, что одно из этих четырех оснований прикреплено к каждому кольцу рибозы на позвоночнике, причем выбор основания в каждом положении с виду случаен. Возможно, вы начнете понимать, почему люди считали ДНК скучной.
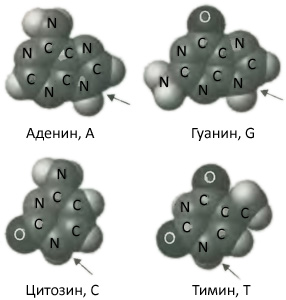
Рис. 2.5. Четыре основания, образующие буквы генетического кода. Аденин (A) и гуанин (G) являются пуринами, цитозин (C) и тимин (T) — пиримидины. (Не помеченные маленькие светло-серые атомы являются водородом.) Стрелками указаны атомы азота, формирующие связи с молекулами рибозы в ДНК.
Как только в ДНК увидели генетическое вещество, возник огромный интерес к деталям ее структуры. Туман над этой структурой начал развеиваться, когда австро-американский биохимик Эрвин Чаргафф (р. 1905), родившийся в городе Черновцы в западной Украине (входившем в Австрию под именем Черновиц) и эмигрировавший в Соединенные Штаты для работы в Колумбийском университете в Нью-Йорке, обратил свое внимание на эту проблему. В 1950, используя новую технику «бумажной хроматографии», которая позволяла разделять и идентифицировать тесно связанные виды соединений путем нанесения их смесей на полоску бумаги, Чаргафф обнаружил равные количества аденина и тимина и равные количества гуанина и цитозина, независимо от вида ткани, из которой он экстрагировал ДНК. Из этого следовало предположение, что аденин каким-то образом всегда ассоциирован с тимином, а гуанин всегда ассоциирован с цитозином. Он также обнаружил, что пропорции долей каждой пары оснований различаются от вида к виду, но являются одинаковыми для различных клеток одного и того же животного. Наблюдения показали, что существует не одна, а много ДНК, и что состав каждой ДНК специфичен для данного организма, в точности как если бы это была его светокопия. Чаргафф также обнаружил, что какие бы виды он ни использовал в качестве источника ДНК, полное количество пуринов (двойных колец аденина и гуанина) является таким же, как полное количество пиримидинов (одиночных колец цитозина и тимина). Вся эта информация оказалась, безусловно, решающей для распознавания структуры ДНК и, как выяснилось задним числом, является почти достаточной, чтобы понять структуру молекулы.
Роль ветра, который окончательно унес остатки тумана, сыграла информация, полученная с помощью экспериментов по диффракции рентгеновских лучей, которые провели новозеландец Морис Уилкинс (р. 1916) и Розалинда Франклин (1920-1958) в Королевском колледже в Лондоне, и дальнейшее развитие их результатов Френсисом Криком (р. 1916 в Нортгемптоне) и Джеймсом Уотсоном (р. 1928 в Чикаго) в Кембридже. Как уже в тысячу раз было пересказано, это была история надувательства, соперничества, напористости, рвения, враждебности, трагедий, женоненавистничества, мошеннических трюков, в большей мере, чем можно вообразить. Вероятно, нельзя считать слишком большим сюрпризом тот факт, что одно из наиболее важных открытий двадцатого века с неизбежностью вызвало к жизни наиболее человеческие эмоции и взаимоотношения.
Трагической фигурой, безусловно, была Франклин, умершая в тридцать семь лет от рака яичников, почти наверняка вызванного облучением рентгеновскими лучами, которыми она пользовалась в своей работе: жизнь не выдает своих секретов, не требуя жизни взамен. Соблазнительно было бы произвести Франклин из трагической фигуры в трагическую героиню и поставить ее в центр всей истории, но это не соответствует фактам. Факты этой очень человеческой истории выглядят следующим образом. Они должны дать абрис обстановки в Британии середины двадцатого века, когда с сегодняшней точки зрения отношение мужчин к женщинам было… неразвитым.
Уилкинс работал над ДНК в Королевском колледже, когда глава лаборатории, намереваясь построить рентгеновский аппарат, пригласил Франклин поработать в колледже и вложить в дело свои специальные познания в рентгеновской кристаллографии. Она приобрела эти познания, изучая микроструктуру угля в парижской лаборатории и была живо заинтересована в том, чтобы переключить свое внимание на живую жизнь в большей степени, чем на ископаемую. Было не вполне ясно, удастся ли ей совершить эту перемену места работы, поскольку Королевский колледж в то время запрещал женщинам находиться в его общей комнате.[9] Уилкинс отсутствовал в момент ее появления и, возвратясь, был приведен в замешательство ролью новой сотрудницы. Немедленно произошло столкновение темпераментов, и каждый из соперников создал свою лабораторию для работы над ДНК. Обе группы вскоре получили весьма неплохие рентгеновские фотографии нитей, образующих эту молекулу. На конференции в Неаполе Уилкинс встретил молодого американского биолога Джеймса Уотсона и показал ему свои изображения. Это побудило Уотсона начать работу над структурой ДНК, и в сентябре 1951 г. он отправился в Кембридж, чтобы изучить дифракцию рентгеновских лучей в лаборатории, которой заведовал сэр Лоуренс Брэгг, один из основателей рентгеновской кристаллографии. Здесь Уотсон встретил Френсиса Крика, как раз заканчивавшего докторскую диссертацию.
В ноябре 1951 г. эти два потока усилий столкнулись, один, имевший тщательно проделанные измерения, но лишенный отваги (или способности) предложить собственную их интерпретацию, другой со смелыми умозаключениями, но без ресурсов (или терпения) для проведения измерений. Уотсон приехал в Лондон и выслушал сообщение Франклин о ее работе. Он поторопился назад в Кембридж, где вместе с Криком они построили модель, которую считали соответствующей тому, что Уотсон смог запомнить из данных Франклин, и пригласили Лондонскую команду приехать и посмотреть на нее. Построение моделей — реальных физических моделей, собранных из проволоки и кусков металла — демонстрировало могущество техники в деле прояснения структуры белков, и Крик и Уотсон просто следовали моде своего времени. Лондонская команда приехала и немедленно отвергла модель как несогласующуюся с их данными. Они также отвергли и сам метод сооружения моделей, метод потенциально (а как оказалось, и реально) продуктивный. Более того, Брэгг приказал Крику и Уотсону прекратить работу над ДНК, оставив ее Лондонской команде, которой и принадлежал весь проект. Отношение к собственности в науке, так же как отношение к женщине, изменилось с тех пор: возможно, следующий шаг и отмечает поворотный пункт к будущему.
В 1952 г. Крик и Уотсон узнали, что Линус Полинг, весьма успешно исследовавший структуру белков, в которой Брэгг не разбирался, работает над той же проблемой. Если работает Полинг, решили они, значит, собственность на проблему уже ускользнула из рук лондонцев, и они имеют право работать над ней, как и любой другой. Далее случилось нечто немного странное. В этот момент Уилкинс без ведома Франклин показал Уотсону одну из ее рентгеновских фотографий (рис. 2.6), а Макс Перуц предоставил ему и Крику неопубликованный доклад в Совете медицинских исследований, в котором Франклин сводила вместе свои последние данные. Наконец-то они получили некоторые определенные числа, характеризующие размеры спиральной молекулы, и смогли подогнать к ней свою модель. Через несколько недель они уже имели возможность с триумфом отослать Уилкинсу свою знаменитую модель, и он ее получил. Трио публикаций, одна Крика и Уотсона, одна группы Уилкинса и одна группы Франклин (Франклин так никогда и не узнала, что Уилкинс воспользовался ее данными), появилось в Nature 25 апреля 1953 г. Две последних предоставили данные эксперимента, подтверждающие умозрения первой. Эта дата, 25 апреля 1953 г., является днем рождения современной биологии.

Рис. 2.6. Этот снимок дифракции рентгеновских лучей, полученный Розалиндой Франклин, был решающей для понимания детальной структуры ДНК частью экспериментальных данных. Он подтверждает, что эта молекула имеет форму двойной спирали, а детали фотографии могут быть использованы для определения размеров этой спирали.
Структура ДНК теперь повсеместно известна как знаменитая символическая правосторонняя двойная спираль, в которой одна длинная нить нуклеиновой кислоты обернута вокруг другой, образуя сплетенную пару (рис. 2.7), которая весьма похожа на сплетенные лестницы входа для публики в музее Ватикана[10], что выглядит немного иронично. Ключевым моментом, однако, оказывается то, что нуклеотидные основания одной нити являются парными к нуклеотидам другой (рис. 2.8), так что аденин всегда в паре с тимином (что мы обозначаем как A…T), а гуанин всегда вместе с цитозином (что обозначаем как G…C). Эта парность соответствует наблюдению Чарграффа, показавшему, что количество аденина в его образцах таково же, как количество тимина, а количество гуанина равно количеству цитозина: комплементарность гарантирует равенство их количеств. Можно также отметить, что относительно маленький пурин (аденин и гуанин) всегда спарен с более крупным пиримидином (тимин и цитозин), поскольку таким способом поддерживается форма двойной спирали: два больших пурина попадают в выпуклость, а два маленьких пиримидина в снижающуюся часть витка спирали. Парность соответствует и другому наблюдению Чарграффа: количество пуринов (A+G) в образце равно количеству пиримидинов (T+C).
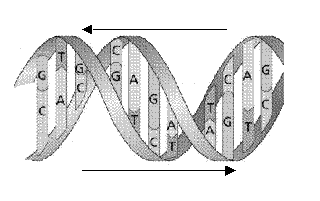
Рис. 2.7 Двойная спираль ДНК. Две нити нуклеиновой кислоты накручены друг на друга, образуя сплетенную двойную спираль с одним узким и одним широким желобом. Две нити удерживаются вместе водородными связями между основаниями, причем пуриновые основания (A, G), представленные длинными стержнями, связаны с пиримидиновыми основаниями (C, T), представленными короткими стержнями. Парными всегда являются А…T и G…C.

Рис. 2.8. Пары оснований, которые удерживаются вместе на нити ДНК, создавая двойную спираль. Водородные связи между молекулами представлены линиями. Заметьте, что пурин связывается с пиримидином и что объемы обеих пар приблизительно одинаковы.
Две нити нуклеиновой кислоты сцеплены между собой благодаря весьма специфическому виду химической связи, известному как водородная связь. Когда я говорю «специфический вид связи», я не имею в виду его малую распространенность, поскольку почти каждая молекула воды в каждом океане связана со своими соседями посредством этого вида, так что только в океанах есть 1044 таких случаев, не считая гораздо большего их числа в других местах. Водородная связь является специфической в том смысле, что она формируется необычным путем и только между несколькими определенными атомами, среди которых кислород и азот. При образовании этой связи атом водорода (который настолько мал, насколько вообще это возможно для атома) попадает между двумя другими атомами и, действуя подобно клею, связывает их вместе. Одним из ключей к пониманию двойной спирали является то, что, как видно на рис. 2.8, форма и расположение атомов азота, кислорода и водорода в тимине и аденине как раз подходят для формирования двух очень удобных водородных связей. Цитозин и гуанин тоже очень удачно подходят друг к другу, но образуют три водородных связи. Тот факт, что водородные связи много слабее, чем обычные химические связи, держащие вместе атомы устойчивых молекул, означает, что две составляющих двойной спирали могут довольно легко отрываться друг от друга, оставляя сами нити нуклеиновой кислоты неповрежденными, в точности так же, как испаряется вода без разрушения ее индивидуальных молекул.
Теперь мы способны понять, почему Уотсон и Крик смогли заключить свою короткую, но удивительную статью скромным замечанием:
От нашего внимания не ускользнуло, что специфический вид образования пар, который мы постулировали, немедленно предлагает возможный механизм копирования генетического материала.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава вторая. Предвидение Галена
Глава вторая. Предвидение Галена И тот, кто даст удовлетворительное объяснение этих явлений, должен быть поистине Изобретателем и человеком, очень сведущим в Управлении и Внутреннем Устройстве таких Анатомических Машин. Г. Пауэр Экспериментальная философия, 1664
ГЛАВА ВТОРАЯ. ЛЕТО КРАСНОЕ
ГЛАВА ВТОРАЯ. ЛЕТО КРАСНОЕ Борщевик, дудник,дягиль лекарственный,земляника, зверобой, марьбелая, душица, звезчаткасредняя, кипрей, ежевика,боярышник, смородина, черемуха, жимолость, малина, голубика,черника, вишнядикая, кизили другие Глава втораяВ северных районах
Глава 1. О прогрессивном развитии в биологии
Глава 1. О прогрессивном развитии в биологии Мы, разумные существа, не должны забывать, что наша цивилизация — лишь одно из замечательных явлений природы, зависящих от постоянного притока концентрированной энергии солнечного излучения. Ю. Одум Вопрос о движущих силах
Глава 11. Что дает энергетический подход для теоретической биологии, или попытка ответить на вопросы «почему»?
Глава 11. Что дает энергетический подход для теоретической биологии, или попытка ответить на вопросы «почему»? Многие вещи нам не понятны не потому, что наши понятия слабы, но потому, что сии вещи не входят в круг наших понятий. Козьма
ГЛАВА ВТОРАЯ. В ГЛУБИНЫ КЛЕТКИ
ГЛАВА ВТОРАЯ. В ГЛУБИНЫ КЛЕТКИ Труден путь в неизведанное, и хорошо, когда есть карты пути... Мала клетка, еще меньше ее ядро, и уж совсем малы крошечные тельца в ядре — хромосомы. А ведь в каждой из них тысячи генов. И совсем немного знали бы люди о них, если б сквозь череду
Глава 7. Похвала биологии
Глава 7. Похвала биологии Каждый из нас, к сожалению, хорошо знаком с неприятным явлением, которое все еще бытует кое-где, несмотря на строгие приказы министерств торговли, культуры и Госкомиздата СССР. Речь идет о так называемой «продаже с нагрузкой». Согласитесь, крайне
Глава 3 Рождение современной биологии
Глава 3 Рождение современной биологии Новая анатомияЗнаменующим началом научной революции принято считать 1543 г. В этом году польский астроном Николай Коперник опубликовал книгу, где была изложена новая точка зрения на Солнечную систему, центром которой было Солнце,
Глава 13 Постсовременное состояние эволюционной биологии
Глава 13 Постсовременное состояние эволюционной биологии Пер. А. СамородоваВ предыдущих двенадцати главах мы рассмотрели многообразие аспектов эволюции жизни. Подобное рассмотрение, конечно, никоим образом не может быть всеобъемлющим, но это и не было моей целью. Так или
Глава вторая: Живые ископаемые
Глава вторая: Живые ископаемые Брат динозавра — крокодил Для жителей северных стран крокодил был “коркодилом”, фантастическим существом со “змиевым хоботом” и “глазами василиска”, проливающим слезы над своею жертвой. Для жителей тех стран, где водились крокодилы,
Глава 1. Нерешенные проблемы биологии
Глава 1. Нерешенные проблемы биологии 1.1. Предпосылки успеха Цель механистического подхода в биологическом исследовании особенно ясно была выражена более ста лет назад Томасом Хаксли в следующем определении:«Зоологическая физиология — это доктрина функций или
Глава вторая Об отражении
Глава вторая Об отражении Объяснив явления световых волн, распространяющихся в однородной среде, мы исследуем затем, что происходит с ними при встрече с другими телами. Сперва мы покажем, как этими же волнами объясняется отражение света и почему при нем сохраняется
Глава I. Основы мичуринской биологии Кандидат сельскохозяйственных наук Е. К. Меркурьева
Глава I. Основы мичуринской биологии Кандидат сельскохозяйственных наук Е. К. Меркурьева 1. Понятие о биологии Биология — это наука о законах развития живой природы. Она изучает жизнь как животных, так и растений. Основная задача биологии состоит в том, что она должна
Глава вторая
Глава вторая Прибытие в тот же день в Борнмут явилось ослепительным контрастом всему, что мы оставили в Манчестере, и для меня было великим облегчением очутиться в кругу людей, всецело симпатизирующих нам и предстоящей женитьбе. Вся семья собралась встретить нас,