Насколько велик репертуар антител?
Насколько велик репертуар антител?
Этот ключевой вопрос был поставлен работами Ландштейнера. Его эксперименты в этой области менее известны неспециалистам, чем открытие системы АВО, хотя они и привели к далеко идущим выводам. Эксперименты Ландштейнера показали гигантский размах разнообразия антител, которые можно получить у лабораторных животных. Превосходный химик, он присоединял мелкие искусственно синтезированные молекулы сложных углеродных колец (например, производных бензола) к белковым антигенам разных типов и демонстрировал образование специфических антител к ним у лабораторных кроликов и мышей. Эти мелкие молекулы (гаптены) сами по себе обычно не вызывали образования антител. Однако будучи присоединенными к белковому носителю, они образовывали гаптен-бел-ковые комплексы, которые становились мощными антигенами, вызывающими образование антител, специфичных и к гаптеновой части, и — еще больше — к белковой.
Само по себе образование антител против таких антигенов не было сколько-нибудь удивительным, так как иммунологи давно получали антитела путем искусственной вакцинации крыс, мышей, кроликов, морских свинок, коз и лошадей. Поразительным было другое, Ландштейнер смог выявить антитела против всех новых химических соединений и лекарств, только что созданных зарождающейся в те годы фармацевтической промышленностью. Этих новых антигенов никогда раньше не было в природе! Этот результат поставил нас перед следующей научной — или даже философской — проблемой. Почему химические вещества, созданные в лабораториях современных фармацевтов, которых до этого никогда не было на Земле, могут вызывать иммунный ответ? Не могло быть никакого давления естественного отбора, направленного на образование антител против субстрата, которого никогда не существовало! Однако Ландштейнер смог показать, что лабораторные животные создают антитела, специфически связывающиеся с любой уникальной молекулярной структурой.
Истинную специфичность антител, полученных Ландштейнером, можно продемонстрировать простым лабораторным опытом. Например, было обнаружено, что антитела к гаптенуА не могут быть удалены из антисыворотки реакциями с гаптена-ми В, С, D, Е, G, Н... и т. д. Другими словами, антитела к гапте-ну А специфичны и не имеют сродства (или имеют очень слабое сродство) ко всему ряду рассматриваемых гаптенов. Однако в более чувствительных тестах — реакциях преципитации — можно наблюдать картину перекрестной реактивности (рис. 3.3). На рисунке знак минус (—) означает, что нет заметного осаждения; один или несколько знаков плюс (от + до +++) указывают на интенсивность осаждения и означают присутствие нерастворимого комплекса антиген—антитело; очень слабое осаждение обозначается (±). По-видимому, антигены D и G как-то связаны с А. И в самом деле, Ландштейнер смог показать, что D и G имеют с А общую молекулярную структуру.
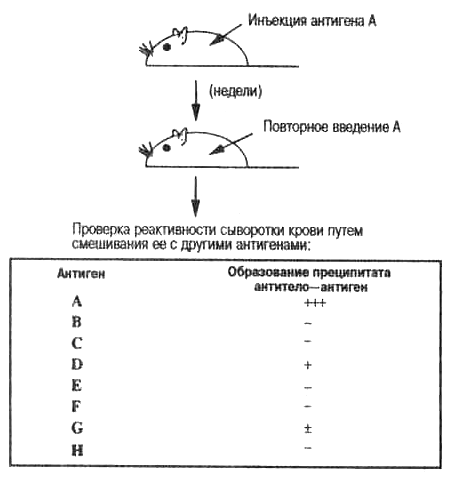
Рис. 3.3. Основа специфичности антител. Иммунизация определенным антигеном (А) вызывает образование антител, которые связываются с А, но не с В, С и т. д. Простейший способ проверки антител состоит в смешивании сыворотки крови с антигенами. Антитела связывают антигены, и на дне пробирки образуется осадок (преципитат). Преципитат в пробирке свидетельствует о наличии антитела, которое может связываться с данным антигеном (- нет преципитата; + следы, +++ сильный преципитат). В данном примере антигены D и G имеют формы (эпитопы, или детерминанты), похожие на А; говорят, что они перекрестно реагируют с А. (также см. табл. 3.1 и рис. 3.6)
Эксперименты Ландштейнера послужили основой первой оценки размера потенциального репертуара антител. Тридцать лет назад был обычным такой комментарий генетических и эволюционных следствий, вытекающих из результатов Ландштейнера: «Тем не менее, трудно понять, какие преимущества дает существование генов для синтеза антител против многих недавно синтезированных органических соединений, таких как п-амино-бензоат, 2,4-динитробензол и т. д., которые совершенно не похожи на микробные патогены. Сохранение такого избытка генетического багажа в течение бесконечного времени до тех пор, пока не появится мишень для них, кажется неправдоподобным»
На самом деле это неправдоподобно в рамках дарвиновского естественного отбора генов, кодирующих определенные специфичные антитела.
Итак, к 1930-м годам стало ясно, что потенциальный репертуар антител должен быть гигантским и насчитывать миллионы или больше специфичностей. Ответ на другой фундаментальный вопрос все еще не был ясен: как могла в ходе эволюции появиться система, способная производить антитела, которые никогда не были частью эволюционной истории данного вида? Если инфекционные болезни были орудием эволюции современной иммунной системы, то, несомненно, частные и специфические молекулярные характеристики множества возможных антигенов никогда не были ее движущей силой. Правильнее, по-видимому, считать, что в ходе эволюции возникла биологическая стратегия, способная производить иммунный ответ на неожиданное.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Структура антител
Структура антител Теперь кратко рассмотрим структуру молекулы антитела. Все антитела имеют общий план строения — это белки, состоящие из нескольких субъединиц. На рис. 3.2 показано строение наиболее обычного антитела, называемого IgG. Эта структура была впервые описана в
В крови плавают десятки тысяч различных антител, но они имеют общий план строения.
В крови плавают десятки тысяч различных антител, но они имеют общий план строения. — Как устроены молекулы антител? Это, наверное, белки, которые циркулируют в крови. — Да, это самые крупные белковые молекулы кровяной сыворотки — гамма–глобулины. Поскольку все
Как велик возраст Вселенной и на основе каких данных он определен?
Как велик возраст Вселенной и на основе каких данных он определен? В 2003 году с помощью запущенного NASA (Национальным управлением США по аэронавтике и исследованию космического пространства) космического зонда, оснащенного специальной аппаратурой, были проведены
Как велик суточный перепад температуры на поверхности Луны?
Как велик суточный перепад температуры на поверхности Луны? Суточный перепад температуры на поверхности Луны весьма велик: температура опускается до минус 170 градусов Цельсия в ночное время и поднимается до плюс 130 градусов Цельсия, когда Солнце в лунном зените. Тем не
Насколько опасны астероиды?
Насколько опасны астероиды? Шкала опасности астероидов, принятая Международным астрономическим союзом, градуирована от 0 до 10 баллов. Ноль получает астероид, орбита которого хотя и пересекается с орбитой Земли, но у него нет никаких шансов на столкновение. Десяткой
Как велик Большой Каньон?
Как велик Большой Каньон? Каньонами называют глубокие речные долины с очень крутыми, нередко отвесными склонами и узким дном, обычно полностью занятым руслом реки. Один из крупнейших каньонов мира – Большой Каньон реки Колорадо в США – имеет длину более 320 километров,
Как велик самый маленький участок суши, которым одновременно владеют две страны?
Как велик самый маленький участок суши, которым одновременно владеют две страны? Самый маленький участок суши, которым владеют сразу две страны, – это островок Сен-Мартен в Карибском море. Его площадь всего 96 квадратных километров, и вот уже почти 350 лет им без ссор
Как велик самый большой в мире розовый куст?
Как велик самый большой в мире розовый куст? По данным на 2001 год, самый большой в мире розовый куст растет в городке Томбстоун (Аризона, США). Он вырос из посаженного в конце XVIII века черенка, и сейчас его ствол имеет периметр у основания более полутора метров. В сезон на этом
Насколько распространен в мире диабет?
Насколько распространен в мире диабет? По оценкам Всемирной организации здравоохранения, в 2001 году в мире было около 150 миллионов диабетиков, а к 2025 году их количество вырастет
Я велик и могуч
Я велик и могуч Большинство животных растут всю жизнь. У таких видов встреча двух взрослых особей, одна из которых много больше другой, — обычное дело. Тот, кто старше, много крупнее, сильнее и опытнее. Драку с ним маленький противник заведомо проиграет. Поэтому у рыб,