Глава 4 КЛОНАЛЬНО-СЕЛЕКЦИОННАЯ ТЕОРИЯ
Глава 4
КЛОНАЛЬНО-СЕЛЕКЦИОННАЯ ТЕОРИЯ
Первую «селекционную» теорию образования антител предложил в 1900 г. Пауль Эрлих (Ehrlich). Согласно его теории, существуют клетки (по-видимому, В-лимфоциты), на поверхностной мембране которых расположено много разных молекул антител. Эти клетки способны синтезировать любое из них. После того как происходит связывание чужеродного антигена с каким-то одним антителом, клетка начинает производить антитела только этой специфичности. Поскольку этот «селективный» процесс происходит одновременно в большом числе клеток, образуется много антител, специфичных к данному антигену. Сейчас известно, что идея Эрлиха не верна. Современные селекционные теории, основанные на представлениях о том, что одна клетка может продуцировать только антитела одного типа (а не многих), начали появляться только в 1950-х гг.
Первой среди них была теория, предложенная Нильсом Ерне (Jerne) в 1955 г. Несмотря на ошибочные представления об антителах как основных единицах отбора, его вклад в проблему был плодотворен. Его теория переместила интересы иммунологов с инструктивных теорий, по которым антитела принимают любую форму в зависимости от формы антигена, на селекционные. В 1957 г. Макфарлейн Бернет предположил, что основной единицей отбора антигеном является клетка, и что одна клетка отвечает за образование антител только одного типа. Именно Бернет придумал термин «клональная селекция» [I]. В основе этой теории лежит представление о том, что «одна клетка производит только одно антитело» (точнее, антитела одной специфичности). Существует много разных клеток (лимфоцитов), образующих и несущих на поверхности разные антитела. Клетка «отбирается» антигеном, форма которого соответствует антителам, производимым данной клеткой. Именно она начинает размножаться и дает клон идентичных клеток, причем все клетки клона продуцируют антитела одной специфичности. Согласно этой теории, должен существовать механизм, который обеспечивает проявление на поверхности клетки антител только одной специфичности и исключает все другие антитела. Теперь мы знаем, что «решение», какое антитело будет синтезироваться в В-клетке, принимается на ранних стадиях развития лимфоцита. Бернет не только нашел экспериментальные свидетельства в пользу этой теории, но и сформулировал ее следствия для проблемы различения «своего» и «не-своего».
В краткой форме мысль об отборе клеток была высказана Дэвидом Талмейджем (Talmage) в 1957 г., но Бернету мы обязаны всесторонней разработкой этой теории. Далее клонально-селекционная теория развивалась усилиями таких исследователей, как Мелвил Кон и Элистэр Каннингем (Cunningham). До сих пор основные представления этой теории объясняют, как иммунная система приспосабливается к разнообразным и постоянно меняющимся антигенам внешней среды. Основные положения клонально-селекционной теории подтверждены экспериментально (рис. 4.1).
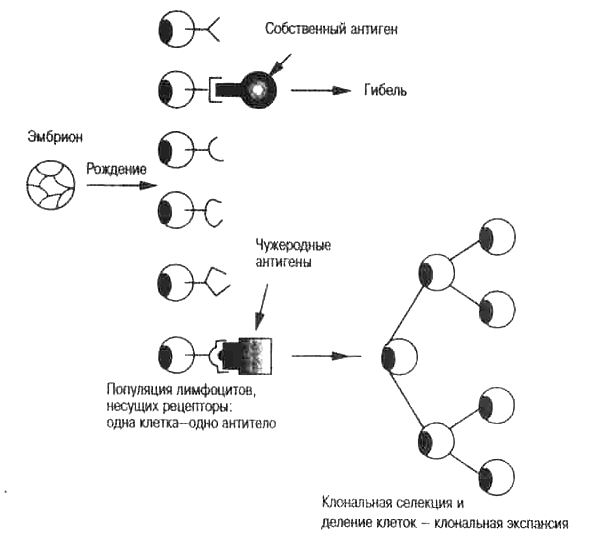
Рис. 4.1. Клональная селекция. Эта идея иллюстрируется также на рис. 1.2 и в табл. 3.1.
Привлекательность этой теории состояла в том, что она давала разумные объяснения механизма аутотолерантности. Если рецептор на поверхности развивающегося незрелого лимфоцита связывается с собственным антигеном, клетка получает «отрицательный» сигнал и уничтожается. (В отличие от этого, зрелые лимфоциты должны размножаться, продуцировать и секре-тировать антитела, если их рецептор связан с антигеном.) Так как собственные антигены — это первые молекулы, которые встречаются незрелым лимфоцитам, этот процесс должен происходить в местах развития лимфоцитов. Бернет назвал его уничтожением «запрещенных» клонов. Это очень элегантное теоретическое объяснение проблемы аутотолерантности. Только лимфоциты, прошедшие этот селекционный фильтр (уничтожение запрещенных клонов), достигают зрелости и приобретают способность связываться с чужеродными антигенами. С тех пор, как в 1957 г. была сформулирована клонально-селекци-онная теория, проведено множество иммунологических исследований, выявлены разные классы лимфоцитов (В- и Т-клетки) и роль каждого из них в иммунном ответе, однако основные положения этой теории до сих пор справедливы.
Итак, мы видим, что в основе клональной селекции лежит простой дарвиновский отбор. Предполагается, что до встречи с антигеном уже существует множество разнообразных клеток, несущих рецепторы. Свои и чужие антигены играют только селективную роль (соответственно отрицательную или положительную) в формировании репертуара лимфоцитов в крови и лимфатической системе. Предполагается, что есть единственное событие, от которого зависит отбор, — это реакция связывания поверхностного антитела В-клеток (или рецепторов Т-клеток) с антигеном определенной формы. Популяции клеток, образовавшиеся в результате «клонального роста», продуцируют специфические антитела и Т-клетки, необходимые для борьбы с инфекцией. Клональный рост может увеличивать число клеток в тысячи раз. Это объясняет, почему концентрация антител в крови растет в экспоненциальной зависимости от времени после заражения или искусственного введения чужеродного антигена (рис. 3.8). Увеличение концентрации антител отражает экспоненциальный рост популяции продуцирующих их клеток, численность которых удваивается при каждом делении. После пятнадцати делений должно появиться около 30 тысяч клеток, возникших из одной исходной клетки, так как число клеток описывается 2"-степенньш рядом, где п равно числу делений. Такое увеличение численности клеток, секретирую-щих антитела, дает возможность быстро достичь эффективной концентрации антител, достаточной для борьбы с острой инфекцией. Как правило, это занимает от трех до пяти дней.
Легко представить себе, как клонально-селекционная теория объясняет иммунологическую память. Она является следствием (по крайней мере, частично) размножения стимулированных антигеном клеток (клональная экспансия). Если антигенспецифичные клетки размножились, некоторые из их потомков становятся долгоживущими. Эти клетки «памяти» могут оставаться в кровеносной системе и в лимфоидных тканях, дожидаясь следующей встречи с тем же самым антигеном спустя много лет.
Эта теория дает также объяснение феномена созревания аффинности (сродства антитела к антигену). Образующиеся на поздних стадиях иммунного ответа антитела имеют большую аффинность, чем те, что образовались ранее (рис. 3.8). По мере того, как концентрация антигена в лимфоидной ткани падает, конкуренция за связывание с редким антигеном приводит к тому, что для размножения «отбираются» более успешные В-клет-ки. В-лимфоциты с поверхностным рецептором самой высокой аффинности (антиген связывается более прочно) будут побеждать в соревновании, и синтезируемый ими тип антител станет, в конечном счете, преобладающим. В следующей главе (рис. 5.4) мы рассмотрим этот процесс отбора, основанного на сродстве, более подробно.
Если изучить каждое антитело в образце крови, то можно обнаружить тысячи разных молекул антител, причем каждое способно распознать специфичный антиген внешней среды. Если определить аминокислотную последовательность каждого из этих антител, окажется, что одно антитело отличается от другого последовательностью аминокислот в вариабельной V-области, которая образует антигенсвязывающий центр (рис. 3.2). Этот факт был установлен в 1960-х гг., и он дал решающие доказательства истинности клонально-селекционной теории Бернета.
Важная экспериментальная проверка клонально-селекционной теории была предпринята в конце 1960-х гг. нашим коллегой Гордоном Эйда (Ada), работавшим с Полин Берт (Byrt) в Институте медицинских исследований Уолтера и Элайзы Холл. В основе их эксперимента лежало предположение, что если популяцию лимфоцитов подвергнуть воздействию антигенов одного типа, с ними будет связываться только клон лимфоцитов с антителами, реагирующими на этот антиген, тогда как огромное большинство клеток не будет связываться, т. к. они продуцируют антитела, реагирующие на другие антигены. Создавая радиоактивные антигены, Эйда и Берт рассуждали так: лимфоциты, которые свяжут их, будут облучены и погибнут. Таким образом, популяция лимфоцитов окажется неспособной к иммунному ответу на проверяемый радиоактивный антиген, но будет нормально реагировать на другие антигены. Именно такой результат они и получили. Этот эксперимент стал известен как «эксперимент по самоубийству антигеном». Он был первым экспериментальным доказательством клонально-селекционной теории.
Отталкиваясь от клонально-селекционной теории, Тед Стил и Боб Бландэн стали изучать механизм эволюции генов антител. Эта новая область иммунологии исследует генетические особенности процессов, обеспечивающих разнообразие антител и созревание аффинности. С конца 1970-х гг. становится все больше известно об уникальной системе генов Ig и ТкР. Сейчас мы уже много знаем о структуре последовательностей ДНК этих генов. Мы можем объяснить, почему одна клетка производит антитела одной специфичности, и как мутация порождает новое антитело. Именно это делает иммунную систему способной продуцировать антитела высокой аффинности ко многим тысячам новых антигенов.
Клонально-селекционная теория поставила вопрос о том, подвергаются ли Ig-гены изменениям в отдельных В-лимфоци-тах после стимуляции антигеном. В общей форме этот вопрос обсуждал еще Бернет в 1957 г. Однако он полагал, что разные Ig-гены, кодирующие антитела различной специфичности, существуют уже до того, как чужеродные антигены внедряются в систему. (Дарвин в предьвдущем столетии также считал доказанным предсуществование генетической изменчивости животных и растений, на которую действует естественный отбор.) Таким образом, Бернет предполагал, что гены антител экспрес-сируются в популяции клеток, последние затем могут отбираться антигеном и клонально размножаться. (Современные методы молекулярной биологии, необходимые для ответа на вопрос о соматическом мутировании в то время были недоступны.)
Тем не менее в конце 1960-х-начале 1970-х гг. Мелвин Кон, а затем Элистэр Каннингем привели доводы в пользу того, что иммунная система имеет способность генерировать соматические мутации генов антител в ответ на внедрение чужеродных антигенов. По их мнению, по-видимому, выгодно, чтобы с ДНК зародышевой линии наследовалось только небольшое число необходимых Ig генов, а новые могли бы возникать в течение жизни животного в виде соматических мутаций, вызванных антигеном. В то время, когда эта теория соматического мутирования была высказана впервые, она была оценена по достоинству только небольшим числом иммунологов. Большинство продолжало придерживаться мнения, что все антитела закодированы в генах половых клеток, то есть они предсущест-вуют до рождения животного. Это предполагало, что изменчивость является результатом случайных (редких) мутаций в генах клеток зародышевой линии, которые передаются потомкам. Без преувеличения можно сказать, что споры между приверженцами идеи «соматических мутаций» и «мутаций в половых клетках» были в центре внимания большинства наиболее важных работ. Они сыграли ту же роль, что и дискуссия в физике о кор-пускулярно-волновой двойственности в квантовой механике в 1920-х годах. Поэтому мы вынуждены объяснять основные молекулярные механизмы в контексте их исторических корней.
Почему антитела специфичны и как приобретается аутотолерантность
Вернемся к проблеме различения своего и не-своего. Мы уже говорили, что для различения своего и не-своего антитела должны быть специфичными. Таким образом, для того чтобы отличать огромный внутренний мир молекул от внешнего мира (а некоторые химические характеристики этих миров одинаковы), эволюция обеспечила ошеломляющее разнообразие молекул, распознающих антигены. Антитела каждой специфичности клонально экспрессируются в популяции тысяч клеток, и простой механизм удаления клона выбраковывает те клетки, которые реагируют на самого себя. Это придает особое значение специфичности и разнообразию антител. В качестве простого, но не строго корректного сравнения [2] можно привести четкость изображения на экране телевизора, которая зависит от числа минимальных элементов изображения (пикселов) в строке экрана. Чем плотнее строка, тем четче изображение. Для того чтобы легко различать разные участки и, следовательно, идентифицировать изображение, необходимо иметь набор разных пикселов (светлые, темные, красные, зеленые и так далее). По аналогии, чем меньше специфичных антител и рецепторов Т-лимфоцитов, тем большему риску уничтожения они подвергаются в процессе установления аутотолерантности. Если бы все они были неспецифичны, было бы два возможных исхода. Первый — после того, как установилась аутотолерантность, у нас не осталось бы иммунной системы. И второй — если бы аутотолерантности не существовало, наша собственная иммунная система атаковала бы и разрушала организм, которому она принадлежит.
Поскольку мы приняли необходимость существования аутотолерантности, мы должны считать аксиомой утверждение, что аутотолерантность каждого индивида должна «обучаться» в дарвиновском смысле этого слова. Поясним это. Если самцы инб-редной линии А мышей скрещиваются с самками генетически другой инбредной линии В, то появляется потомство, которое унаследовало разные гены от разных родителей. Мыши линии А генетически запрограммированы производить в клетках и тканях молекулы А-антигена, тогда как мыши линии В запрограммированы производить В-антигены. В некотором смысле это напоминает правила переливания крови у человека для системы групп крови АВО. Взрослые особи линии А всегда будут развивать анти-В иммунную реактивность (естественно развивающиеся антитела и Т-клетки), а особи В будут развивать анти-А реактивность. Таким образом, ткань, пересаженная от А к В и, наоборот, от В к А, всегда будет отторгаться. Однако потомки генетически различающихся родителей будут по своему антигенному составу АВ, и это представляет для них серьезную проблему. Если бы аутотолерантность не приобреталась в ходе развития, АВ потомки этого скрещивания не могли бы выжить. Они должны были бы саморазрушиться, как только их иммунная система приобрела бы способность реагировать на свои собственные клетки. Однако мы знаем, что популяции человека и большинство популяций диких животных чрезвычайно разнообразны по генам, кодирующим антигены гистосовмести-мости. Следовательно, каждый из нас несет разные комбинации этих антигенов, унаследованных от родителей. То есть, все мы похожи на гибридов АхВ, изображенных на рис. 4.2.
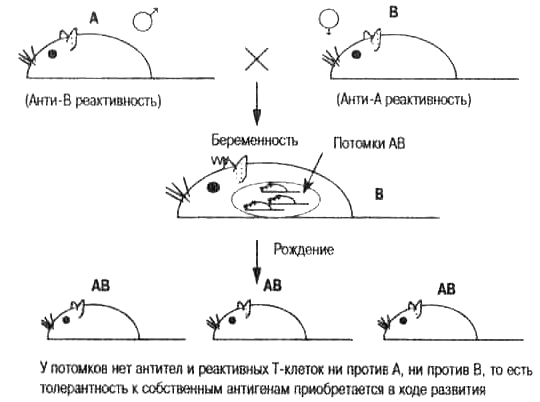
Рис. 4.2. Толерантность к собственным антигенам не детерминирована генетически. Мелвин Кон и его коллеги Петер Бретчер и Род Лангман неоднократно подчеркивали этот ключевой момент. На рисунке показан результат эксперимента, поставленного природой (случайные скрещивания генетически различающихся индивидов). Поскольку взрослое потомство АВ не разрушает само себя аутоиммунной реакцией, это со всей очевидностью свидетельствует о том, что толерантность приобретается в ходе развития организма. Клонально-селекционная теория Макфарлейна Бернета объясняет, как это достигается путем дарвиновского отрицательного отбора в иммунной системе — уничтожением «запрещенных» клонов, направленных против собственных антигенов (рис. 4.1).
Очевидно, что индивид АВ на какой-то стадии своего развития должен «научиться» быть толерантным и к А-, и к В-антигенам. То есть, механизм аутотолерантности должен приобретаться «соматически». В начале 1950-х годов Питер Медавар и его коллеги экспериментально подтвердили эти теоретические предположения, впервые сформулированные Макфарлейном Бернетом и Фрэнком Феннером (Fenner) в 1949 г. Группа Медавара показала, что, если новорожденным мышам линии В ввести клетки линии А, они вырастают толерантными к трансплантатам кожи линии А. Таким образом, мышей линии В можно было соматически модифицировать так, что они приобретали способность не отторгать кожные трансплантаты линии А, которые в норме у взрослых отторгаются (рис. 4.3). Вернет и Медавар в 1960 г. получили Нобелевскую премию за открытие приобретенной иммунной толерантности.
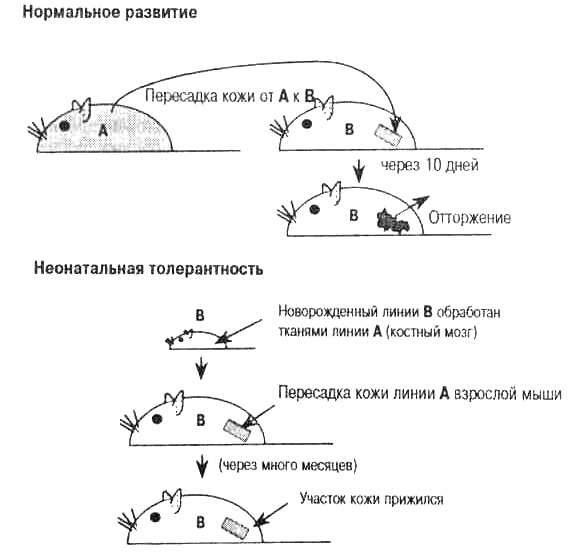
Рис. 4.3. Эксперимент Питера Медавара по приобретенной неонатальной толерантности. Питер Медавар, Лесли Брент и Руперт Биллингем провели свои эксперименты в 1940-х-начале 1950-х годов. Результаты экспериментов согласуются с клонально-селекционной теорией Макфарлейна Бернета. Они показали, что введение в организм на ранних стадиях неонатально-го развития чужих клеток из костного мозга может индуцировать приобретенную иммунологическую толерантность. Введение на этих стадиях чужеродного антигена вызывает уничтожение клонов, направленных против чужих. В результате иммунная система начинает относиться к чужим тканям как к своим.
Итак, мы приходим к заключению, что необходимая для приобретения и иммунности, и толерантности соматически обучающаяся программа основана на ключевом взаимодействии (или связывании) антигенов с клонально экспрессирующи-мися поверхностными молекулами, распознающими антигены (антитела, ТкР). Это достигается в процессе дарвиновского отбора, действующего в популяции лимфоцитов. Это — основной принцип функционирования иммунной системы. Мы объясним в дальнейшем, что обратная связь между генами сомы и половых клеток дает логически последовательное объяснение всех уникальных молекулярно-генетических свойств иммунной системы. Но, прежде чем вести читателя дальше, мы должны потратить немного времени на описание деталей строения ДНК-последовательностей генов Ig и ТкР и некоторых необычных свойств этих генов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ГЛАВА 1. Только лишь теория?
ГЛАВА 1. Только лишь теория? ПРЕДСТАВЬТЕ, что вы преподаватель Римской истории и Латинского языка, стремящийся передать свою увлеченность древним миром — элегиями Овидия и одами Горация, ораторским мастерством Цицерона, стратегическими находками полководцев пунических
ГЛАВА 14 Четвертичный период (антропоген): Великое оледенение. Ледниковая теория. Перигляциальные сообщества и мамонтовая фауна
ГЛАВА 14 Четвертичный период (антропоген): Великое оледенение. Ледниковая теория. Перигляциальные сообщества и мамонтовая фауна Четвертичный период, или антропоген, — самый последний отрезок кайнозоя, начавшийся около 2 млн лет назад. Наиболее существенная черта этого
Глава 1 Только теория?
Глава 1 Только теория? Вообразите, что вы — преподаватель латыни и римской истории. Вы стремитесь привить своим ученикам любовь и уважение к античности: от элегий Овидия и од Горация до латинской грамматики и речей Цицерона, от стратегических находок Пунических войн до
7 Расовая теория
7 Расовая теория Из всех способов уклонения от разбора характера социальных и моральных воздействий на разум человека самый распространенный — объяснение разнообразия поведения и характера человека наследственными природными признаками. Джон Стюарт Милл Совершенно
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции Пер. А. НадирянВ этой и следующей главах дается краткое описание современного состояния эволюционной биологии, какой она была до 1995 года, когда возникло новое направление науки — сравнительная геномика.
Глава 2 Палеобиология и эволюционная теория. Время и изменение
Глава 2 Палеобиология и эволюционная теория. Время и изменение ...Слыша доносящийся издалека шум великих морей, омывающих берега, давным-давно ими же разрушенные, и крики морских птиц, что исчезли с лица земли. Дж. Р. Р. Толкиен «Братство кольца» Абсолютное и относительное
ТЕОРИЯ, ЭВОЛЮЦИЯ И ТЕОРИЯ ЭВОЛЮЦИИ
ТЕОРИЯ, ЭВОЛЮЦИЯ И ТЕОРИЯ ЭВОЛЮЦИИ Термины эволюционируют помимо и вопреки нашей воле. Поэтому даже самое лучшее определение не может считаться окончательным. В прошлом теории (с оттенком пренебрежения) противопоставляли точному знанию, почерпнутому из Библии или, на
Глава II ЭКОСИСТЕМНАЯ ТЕОРИЯ ЭВОЛЮЦИИ
Глава II ЭКОСИСТЕМНАЯ ТЕОРИЯ ЭВОЛЮЦИИ Одним он (Эпиметей) дал силу без быстроты, других же, более слабых, наделил быстротой; одних он вооружил, другим, по природе безоружным, измыслил какое-нибудь другое средство во спасение… Все это он измыслил из осторожности, чтобы не
Глава 3. ЭКЗОТРОФИЯ. НОВАЯ ТЕОРИЯ АДЕКВАТНОГО ПИТАНИЯ
Глава 3. ЭКЗОТРОФИЯ. НОВАЯ ТЕОРИЯ АДЕКВАТНОГО ПИТАНИЯ Питание — одна из центральных проблем, решение которой составляет предмет постоянных забот человечества. Эта проблема актуальна для большинства биологических наук. Однако первоначально наука о питании имела
ОСТРОВНАЯБИОГЕОГРАФИЧЕСКАЯ ТЕОРИЯ
ОСТРОВНАЯБИОГЕОГРАФИЧЕСКАЯ ТЕОРИЯ Хотите узнать, как размер острова влияет на количество видов, обитающих на нем? Это легко сделать: поделите его пополам и наблюдайте за последующими переменами. Хотите узнать, как скорость иммиграции связана с расстоянием до материка?
Глава 2. Классическая теория сбалансированного питания
Глава 2. Классическая теория сбалансированного питания 2.1. Вводные замечания Нет необходимости доказывать, что питание — одна из центральных проблем, решение которой составляет предмет постоянных забот человечества. По-видимому, едва ли не самое большое заблуждение —
Глава 3. Теория адекватного питания
Глава 3. Теория адекватного питания 3.1. Вводные замечания Экспериментальная проверка ряда важных положений и следствий теории сбалансированного питания с учетом ряда новых фундаментальных открытий в биологии привела к их пересмотру. Было обнаружено, что следствием
Глава 2. Классическая теория сбалансированного питания
Глава 2. Классическая теория сбалансированного питания 2.1. Вводные замечания Нет необходимости доказывать, что питание — одна из центральных проблем, решение которой составляет предмет постоянных забот человечества. По-видимому, едва ли не самое большое заблуждение —
Глава 3. Теория адекватного питания
Глава 3. Теория адекватного питания 3.1. Вводные замечания Экспериментальная проверка ряда важных положений и следствий теории сбалансированного питания с учетом ряда новых фундаментальных открытий в биологии привела к их пересмотру. Было обнаружено, что следствием
Химическая теория
Химическая теория И тут на научном горизонте появилась химическая теория. Организм — это фабрика ядов. Таков был её главный тезис. «Мы отравлены с головы до ног продуктами наших собственных органов. Это — общий результат обмена веществ…» — писал в книге «Сон» И. И.
Теория Пенелопы
Теория Пенелопы У племени чероки существует обычай, пишет Леви-Брюль в своей книге «Первобытное мышление», по которому человек, видевший во сне, что он был укушен змеей, должен быть подвергнут тому же лечению, которое применяется при настоящем укусе змеи. В прошлом веке