Основные положения традиционной неодарвинистской теории эволюции
Основные положения традиционной неодарвинистской теории эволюции
• Наследственность
Генетический материал (ДНК) может передаваться неизменным от поколения к поколению.
• Мутации
Изредка в ДНК возникают стойкие изменения - мутации. Чарлз Дарвин называл такие изменения наследственной информации спортами. Например, замена А основания на G приводит к появлению новой последовательности нуклеотидов в данном участке хромосомы, и, соответственно, в информационной РНК. Это приводит к изменению последовательности аминокислот в белке и, следовательно, к изменению его структуры и/или функции. Мутации служат постоянным источником новой генетической информации, на которую может действовать отбор. Полагают, что подобные изменения последовательностей нуклеотидов редки.
• Случайная передача отцовских и материнских хромосом потомку
Оплодотворенная яйцеклетка, из которой развивается человеческий эмбрион, имеет 46 хромосом. В репродуктивных органах (семенниках и яичниках) половозрелых людей происходит особое деление клеток (мейоз), в результате которого число хромосом уменьшается вдвое (гаплоидное число 23). Гаплоидный набор хромосом упаковывается в сперматозоиды у мужчин и в яйцеклетки у женщин. При оплодотворении число хромосом восстанавливается до 46 (диплоидное число). Потомок получает от каждого из родителей 23 случайно выбранные хромосомы, которые родители в свою очередь унаследовали от своих родителей. Все яйцеклетки содержат половую хромосому одного типа, а именно Х-хромосому. В сперматозоидах содержатся либо Х-, либо Y- хромосома. Набор половых хромосом XX определяет женский пол плода, XY - мужской. Все остальные признаки детерминируются случайным сочетанием хромосом. Таким образом, наши обычные житейские наблюдения над детьми справедливы, и если нам кажется, что у «Тома мамино лицо, но папины руки!», значит так оно и есть.
• Рекомбинация
У видов, размножающихся половым путем, физический обмен (кроссинговер, или рекомбинация) между отцовскими и материнскими хромосомами, который происходит во время образования гамет (сперматозоидов и яйцеклеток), вызывает перетасовку существующих последовательностей ДНК. Этот процесс приводит к появлению новых комбинаций наследственных признаков у потомков.
• Естественный отбор и эволюция
Не все генетически разные организмы размножаются одинаково, т.е. с одинаковой скоростью. Условия среды отбирают более приспособленные организмы и дают им селективные преимущества. Если бы не было отбора, не было бы и общих генетических изменений. Эволюция, следовательно, является результатом взаимодействия генетического материала с внутренними (клеточными, внутри организма) и с внешними условиями развития организма и может быть представлена как отбор комбинаций генов, имеющих наибольшую приспособленность.
• Изоляция
В больших популяциях свободное скрещивание между разными генотипами, расширяющее пул генов, в котором действует естественный отбор, невозможно из-за внутренних и внешних изолирующих факторов. Самым простым является географическое отделение потомков от общего предка. Например, некогда единая популяция вьюрков на островах Галапагосского архипелага со временем разделилась на изолированные, небольшие, инбредные популяции. Дарвин заметил, что именно это оказалось важнейшим условием появления новых видов.
• Дрейф
Последовательности ДНК, различаются ли они по большому участку или по одному нуклеотиду, могут быть потеряны по случайным причинам. Генетический дрейф является частью случайного фонового шума, изменяющего сложившийся состав последовательностей ДНК в популяции.
• Барьер Вейсмана
Приобретенные соматические модификации у многоклеточных организмов не могут наследоваться.
Итак, мы представили вам две, отнюдь не несовместимые, концепции. Первая — традиционная неодарвинистская теория о том, что важная для эволюции генетическая изменчивость существует до того, как подействует селективная сила (естественный отбор). И вторая — традиционно отвергаемая ламарков-ская теория о том, что генетическая изменчивость возникает одновременно с отбором. Последняя концепция особенно важна для процессов, протекающих в иммунной системе, для которых селективная сила, или стимул внешней среды (инфекционное заболевание), действует одновременно с появлением новых генетических вариантов.
Таким образом, исторически всегда существовал альтернативный (ламаркисткий) взгляд на механизм эволюционных изменений. Этот механизм легко объясняет, почему некоторые виды смогли очень быстро генетически измениться при внезапных изменениях среды, во время катастроф; также легко объяснить быстрое создание разных пород домашних животных. В этой книге мы рассмотрим, насколько реальны такие изменения с позиций молекулярной генетики. Мы ни в коем случае не утверждаем, что идея Дарвина о естественном отборе случайной предсуществующей изменчивости неверна. Наоборот, мы стремимся доказать, что дарвиновская идея чрезвычайно важна для ламаркистской концепции обратной связи генов сомы и зародышевой линии. По-видимому, правы и Дарвин, и Ламарк. Их взгляды дополняют друг друга. Именно это мы попытаемся продемонстрировать с помощью создаваемой нами теории эво-люционно-генетических изменений иммунной системы.
Это же исторический факт, что сам Чарлз Дарвин, а до него его дед, были ламаркистами. Дарвин разделял идею Ламарка о наследования приобретенных признаков. Ламаркистские доводы об упражнении и неупражнении органов появляются во многих местах его Происхождения видов (1859). Через десять лет после выхода этой книги он публикует теорию пангенезиса [З]. Этот его вклад в науку часто вычеркивается из научной литературы неодарвинистами, которых, по-видимому, сильно задевает тот факт, что основатель теории естественного отбора нашел необходимым воспользоваться ламаркистской «ересью» для объяснения причин генетической изменчивости. Основой для этой теории послужили прекрасные наблюдения Дарвина над изменчивостью и приспособленностью растений и особенно животных в ходе одомашнивания: «Измененные условия вызывают последствия, передающиеся по наследству, например, изменение периода цветения растений, перенесенных из одного климата в другой. У животных усиленная работа или неиспользование каких-то органов оказывают существенное влияние; так, я заметил, что у домашней утки кости крыла весят меньше, а кости ног больше по отношению ко всему скелету, чем те же кости у дикой утки, и это отличие можно с уверенностью приписать тому, что домашняя утка гораздо меньше летает и больше ходит, чем ее дикие предки... .Значительное наследуемое развитие вымени у коров и коз в тех странах, где этих животных обычно доят, по сравнению с животными в других странах, представляет, вероятно, другой пример последствий активной работы, органа»
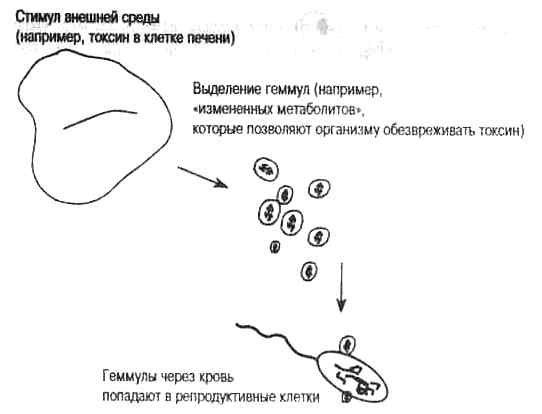
Рис. 1.1. Теория пангенезиса Чарльза Дарвина, 1868. Теория пангенезиса предложена Чарльзом Дарвином для объяснения причин биологической изменчивости, на которую действует естественный отбор. Основная идея его теории «упражнения и неупражнения» заключается в повышении активности органа-мишени, вызванной стимулами внешней среды (например, токсинами в изменившейся пище). Измененная метаболическая активность ткани-мишени (в данном случае, печени) приводит к приспособлению, далее, согласно теории, из этого органа выделяются «геммулы», или «пангены», которые попадают в кровеносную систему и из нее в половые клетки. Таким образом, измененная функция органа фиксируется в половых клетках и наследуется потомками. Следовательно, Дарвин предположил ламаркистский механизм генетической передачи приобретенных признаков.
В 1868 г. Дарвин пришел к заключению, что соматические изменения, появляющиеся в результате специфического приспособления, стимулируют клетки органа-мишени к выделению некоего наследственного материала в форме, которую он назвал геммулами (или пангенами). Геммулы — это представители каждой нормальной или измененной части тела. Онивыделяются активным органом в кровоток, и, циркулируя по телу, могут попадать в половые клетки и передаваться следующему поколению. Рис. 1.1 иллюстрирует теорию пангенезиса Дарвина. Как мы покажем в следующих главах, Дарвин предвидел не только естественный отбор случайных изменений, но и роль выделяемых телом геммул, записывающих наследуемый эффект, как мы говорим сейчас, на ДНК зародышевой линии.
В 1977 г. Тед Стил предложил гипотезу соматического отбора, которая представляет собой современную молекулярную точку зрения на идею пангенезиса (Steele, 1977). Согласно этой гипотезе, эволюция генов вариабельных участков антител происходит через преодоление вейсмановского барьера, т. е. в результате обратной связи генов сомы (тела) и зародышевой линии. Этот механизм допускает появление новых генетических вариантов в ответ на внедрение микробов из внешней среды. Таким образом, гипотеза соматического отбора представляет собой версию ламарковского наследования, основанную на современных молекулярных данных. Вначале эта идея многими была признана как «еретическая». Однако появившиеся за последние двадцать лет данные изменили отношение к ней. Сейчас нет сомнений в том, что в результате активации антигенами вторгнувшегося инфекционного агента гены, кодирующие белки (антитела), необходимые для распознавания чужеродного агента, подвергаются в соматических клетках быстрому мутиро-ванию (гипермутированию). Самые последние исследования подтверждают то, что мутантные гены антител попадают в ДНК зародышевой линии в результате процесса обратной транскрипции. На рис. 1.2 показано, как соматические изменения могут включаться в ДНК половых клеток.
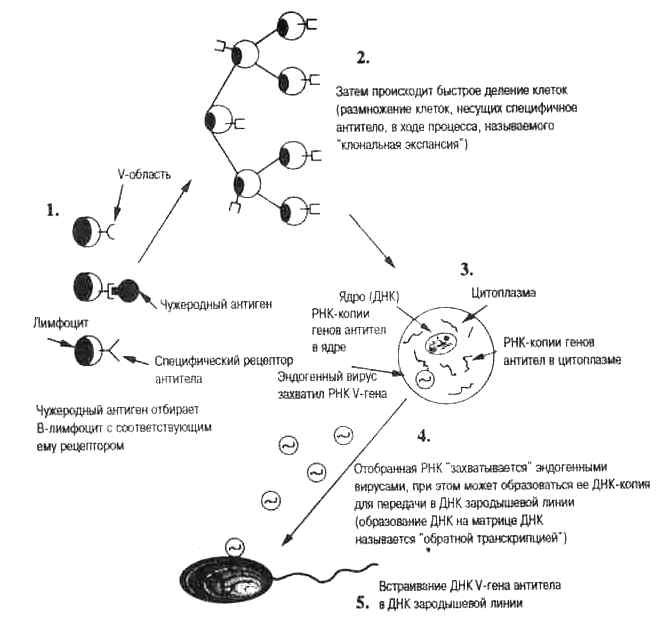
Рис. 1.2. Предполагаемый механизм соматического отбора в иммунной системе.
1. Множество разнообразных В-лимфоцитов существует до того, как чужеродный антиген попадает в организм. Каждая клетка экспесси-рует на своей поверхностной мембране антитела одной специфичности. Гены вариабельной V-области кодируют те участки антитела, которые образуют антигенсвязывающий центр (как показано на рисунке). Чужеродный антиген связывается с В-клетками, имеющими комплементарное антитело — таким образом, эти клетки «отбираются» в дарвиновском смысле («клональная селекция»).
2. После связывания антигена В-клетка активируется и делится, давая потомков, которые в свою очередь тоже делятся. В результате образуется множество идентичных дочерних клеток — клон. Все клетки клона экспрессируют одинаковые антитела («клональная экспансия»). В отдельных клетках этого клона гены вариабельной области могут мутировать (соматические мутации). Эти клетки в свою очередь могут быть отобраны антигеном для образования нового клона.
3. Внутри клеток клона создаются РНК-копии генов вариабельной V-области. Зрелые молекулы информационной РНК переходят в цитоплазму, где они транслируются в последовательности аминокислот, составляющие белковые цепи антитела
4. Молекулы РНК V-генов (ядерные или цитоплазматические) могут «захватываться» безвредными эндогенными РНК-ретровирусами (продуцируемыми клеткой). Вирусный фермент обратная транс-криптаза может синтезировать ДНК-копию этой РНК (эта копия ДНК называется ретротранскриптом, или кДНК).
5. Копии V-генов антител с помощью вирусов переносятся в сперматозоиды или яйцеклетки, встраиваются в ДНК половых клеток (зародышевой линии) и передаются потомкам, (см. гл. 6)
Открытие в конце 1950-х г. Говардом Теминым (Temin) феномена обратной транскрипции поначалу вызвало сомнение. Однако после присуждения в 1975 г. Темину и Балтимору (Baltimore) Нобелевской премии этот процесс признан всеми как важный элемент репликации ретровирусов (таких, как ВИЧ) и некоторых других событий в клетке. (Свое название ре-тровирусы получили вследствие того, что у них поток генетической информации направлен от РНК к ДНК, т. е. в направлении, обратном существующему во всех живых клетках — от ДНК к РНК.) Главная цель этой книги — показать, как новые данные молекулярной генетики разрушают построения неодарвинистов, рассматривающих отбор случайных генетических вариантов в качестве единственного фактора эволюционных изменений. Мы хотим доказать актуальность новой теории эволюции иммунной системы, основанной на объединении концепций Дарвина и Ламарка.
Почему же точка зрения Ламарка вызвала столько споров? В наши планы не входит полный анализ этого вопроса, однако некоторые исторические моменты борьбы дарвинизма и ламаркизма все же следует отметить. Во-первых, в 1885 г., через три года после смерти Дарвина, немецкий биолог Август Вейсман (Weismann), отвечая на вызов, брошенный теорией пангенезиса, провозгласил существование барьера между соматическими и половыми клетками (рис. 1.3), защищающего половые клетки от любого изменения тела. Вейсман пытался проверить, могут ли наследоваться приобретенные родителями увечья. Например, он вырезал аппендикс или другую ткань и показывал, что потомство не наследует этих нарушений. Но любой мыслящий человек и без этого знает, что иудейская традиция обрезания ни разу не привела к рождению мальчика без крайней плоти. Вейсман провел и другие эксперименты, в частности, с гидрой (Hydra). Однако наиболее известна его работа по отрубанию хвостов у только что родившихся крысят. В длившихся много поколений экспериментах Вейсман показал, что отрубание хвостов никогда не приводило к появлению бесхвостого потомства. Ученые, критиковавшие этот эксперимент, указывали, что такими опытами нельзя проверить идею Ламарка. Короткий обрубленный хвост — это модификация, индуцированная не крысой. А по Ламарку наследоваться могут только модификации, индуцированные ответом организма на условия среды. Если бы Вейсман изучал поведение потомков от крыс, лишенных хвостов, он вполне мог бы наблюдать мелкие наследственные модификации поведения (например, при кормлении, чистке, передвижении).
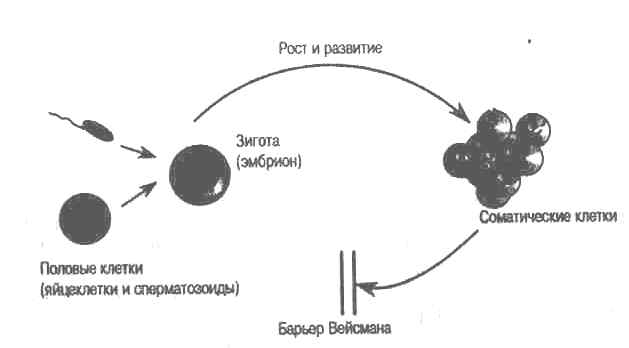
Рис. 1.3. Барьер Вейсмана. В 1885 г Август Вейсман провозгласил существование тканевого барьера, защищающего половые клетки от любого влияния сомы. На языке современных терминов это можно сформулировать так: мутации в соматических клетках (клетках тела) никогда не передаются в клетки зародышевой линии (репродуктивные клетки).
Существование вейсмановского барьера было якобы неопровержимо доказано в 1911 г. работой Кастла и Филлипса (Castle, Phillips) из Гарвардского университета. Они удалили яичники у белой морской свинки (альбиноса) и пересадили ей яичники черной морской свинки [4]. Белая мать-реципиент при скрещивании с нормальным белым самцом дала несколько черных потомков в течение шести-двенадцати месяцев после операции. Точно такой же результат наблюдался бы при скрещивании белого самца с черной самкой. Таким образом, новая «белая» сома не оказала никакого влияния на половые клетки в пересаженных «черных» яичниках. Однако подобные «острые» эксперименты можно критиковать по тем же причинам, что и отрубание хвостов. Нет никаких причин предполагать, что адаптивный ответ самок с пересаженными яичниками должен быть связан с генетическим контролем окраски шерсти.
Гипотезу Ламарка пытался доказать австрийский биолог Пауль Каммерер (Kammerer). Эта работа имела трагические последствия. Каммерер сообщил, что изменение полового поведения и некоторых других инстинктов у морских животных и земноводных может приводить к появлению потомства, имеющего те же поведенческие или морфологические черты, которые приобрели их родители в течение жизни. Его наиболее известные эксперименты проведены на жабах-повитухах Alytes obstetricians. Большинство видов жаб и лягушек спариваются в воде. Самцы этих видов крепко захватывают самку и долго (дни и недели) удерживают ее до тех пор, пока она не отложит икру.
Для того чтобы удерживать скользкую самку, у самцов на ладонях и пальцах имеются мозолистые и ороговевшие брачные бугорки. Alytes спариваются на суше, у них нет таких бугорков, так как кожа самок сухая и грубая. Пауль Каммерер обнаружил, что, если Alytes заставить спариваться в воде, как это делают другие жабы, то через несколько поколений у них появляются брачные бугорки. Это и есть приобретенный наследственный признак.
В начале двадцатого века эти эксперименты вызвали острую полемику, которая, в конце концов, завершилась тем, что профессор Кембриджского университета Вильям Бэтсон (Bateson) обвинил Каммерера в фальсификации. Артур Кестлер в своей книге Case of Midwife Toad («Дело жабы-повитухи») приводит документы по этому трагическому делу и утверждает, что обвинения не были в достаточной мере доказаны. Но удар был нанесен, и не выдержав оскорбления, в 1926 г. Каммерер покончил жизнь самоубийством. Недавно Марк Гиллман (Gillman) проанализировал этот исторический эпизод в книге Envy as a Retarding Force in Science, 1996 («Зависть как тормоз в науке»).
Еще одной исторической причиной, по которой ламаркистское мышление стало «закрытой зоной» в науке, можно считать разгром советской генетики в 40-е гг. Иосиф Сталин поручил селекционеру Т. Д. Лысенко улучшить сельское хозяйство в СССР. К сожалению, пытаясь продемонстрировать увеличение урожая зерновых с помощью приема, названного яровизацией, Лысенко применил неапробированную процедуру. Этот прием заключался в различных способах обработки семян, например, изменении температуры и питания во время прорастания. Он использовался для получения более продуктивных растений, из семян которых, по утверждению Лысенко, вырастают растения лучшего качества. Но лысенковский подход к возрождению идей Ламарка заставил отвернуться от него западньк ученых и генетиков, работающих в Советском Союзе. Проблема противостояния была решена печально известными жестокими преследованиями научных оппонентов Лысенко [5].
Мы считаем, что все эти обстоятельства истории науки и общества надолго затормозили рациональное развитие идей Ламарка, которые сейчас выливаются в представления об обратной связи генов.
Важно упомянуть еще два момента, оказавших влияние на развитие биологической науки. Первое — это, не побоимся сказать, обожествление Дарвина, особенно в Британии. Дарвина превратили в икону, однако мы полагаем, что на определенном этапе это было действительно необходимо для того, чтобы внедрить в умы человечества ключевую идею о естественном отборе случайных изменений. Без этой концепции было бы трудно объяснить многие биологические явления, будь то Ц структура популяции, изменчивость генов или работа иммунной системы. Это было необходимо для противостояния наивному креационизму, утверждавшему, что все виды возникли одновременно и относительно недавно.
Согласно дарвиновской теории эволюции, для появления новых форм и видов необходимо длительное время. Этот факт согласуется с данными современных палеонтологических и геологических исследований. Действительно, между всеми существующими в настоящее время живыми организмами установлено молекулярно-эволюционное родство. Получение данных, позволивших сделать столь важные заключения, стало возможным благодаря появлению в конце 80-х гг. приборов для автоматического определения последовательности нуклеотидов в ДНК (ДНК-секвенаторы). Новая технология дала возможность генетикам и молекулярным биологам получать точную информацию о большом числе генов (о последовательности нуклеотидов в ДНК). Большая часть этих данных собрана в обширных общедоступных базах данных в Интернете, например в Genbank.
Присуждение в 1993 г. Нобелевской премии по химии Кэри Маллису (Mullis) за открытие и разработку метода полимеразной цепной реакции (ПЦР) подчеркивает важность новых технологий в получении научного знания. Метод ПЦР используется с конца 1980-х годов. Он дает возможность увеличивать число копий отдельного участка ДНК в миллионы раз. После этого с помощью секвенатора можно легко определить порядок нуклеотидов A, G, С и Т в этом фрагменте (определения терминов даны в табл. 1.2). Метод ПЦР стал для генетиков новым мощным «телескопическим» средством, позволяющим увидеть молекулярное строение и информационное содержание различных последовательностей нуклеотидов. Именно метод ПЦР, который можно назвать «генетическим копированием», побудил к созданию книги и фильма «Парк юрского периода», показав возможность (пока нереальную) того, что сохранившиеся древние ископаемые останки ДНК можно размножить, а затем с помощью клонирования «воскресить» вымерших животных.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
2.1. Становление и основные положения синтетической теории эволюции
2.1. Становление и основные положения синтетической теории эволюции Эволюционизм возник как альтернатива учению о неизменности видов. Вопросы, связанные с возникновением и развитием жизни, прошли через всю интеллектуальную историю человечества. Количество литературы,
2.2. Альтернативные теории эволюции
2.2. Альтернативные теории эволюции Многообразие альтернативных концепций эволюции обычно группируют в три ветви: ламаркизм, теории направленной эволюции и сальтационизм. Каждая ветвь имеет свою богатую историю. В настоящее время эти названия представляют скорее
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции Пер. А. НестеровойВ этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись
Основные положения канцеро-лейкозогенеза
Основные положения канцеро-лейкозогенеза Ниже изложены основные заключения, следующие из приведенного выше биофизического анализа предраковых и особенно предлейкозных состояний и анализа возможных последствий функционирования выявленных порочных потенциально
ТЕОРИИ О ТЕОРИИ ЭВОЛЮЦИИ
ТЕОРИИ О ТЕОРИИ ЭВОЛЮЦИИ Немало недоразумений возникает из-за неумения отличить общеэволюционный подход от частных метаэволюционных проблем и эти последние друг от друга.На вопрос, в чем различие между теориями Ж. Б. Ламарка и Ч. Дарвина, большинство отвечает: Ламарк
НАУЧНОСТЬ ТЕОРИИ ЭВОЛЮЦИИ
НАУЧНОСТЬ ТЕОРИИ ЭВОЛЮЦИИ Каждому, вероятно, приходилось слышать время от времени, что в биологии нет настоящих теорий. В частности, эволюционизму отказывают в статусе подлинной научной теории по следующим соображениям.1. Это в основном описание всевозможных событий, а
2.3. Основные постулаты теории сбалансированного питания
2.3. Основные постулаты теории сбалансированного питания Теория сбалансированного питания возникла вместе с классическим экспериментальным естествознанием и в сущности остается господствующим мировоззрением и в настоящее время. Основы этой теории изложены в ряде
2.6. Основные следствия теории сбалансированного питания
2.6. Основные следствия теории сбалансированного питания Теория сбалансированного питания замечательна не только стройностью и ясностью логики и экспериментальной обоснованностью, но и способностью к предсказанию неизвестных явлений, которые были затем открыты, или
3.2. Основные постулаты теории адекватного питания
3.2. Основные постулаты теории адекватного питания Кризис теории сбалансированного питания и открытие некоторых витальных, неизвестных ранее механизмов (лизосомального и мембранного пищеварения, различных видов транспорта пищевых веществ, общих эффектов кишечной
2.3. Основные постулаты теории сбалансированного питания
2.3. Основные постулаты теории сбалансированного питания Теория сбалансированного питания возникла вместе с классическим экспериментальным естествознанием и в сущности остается господствующим мировоззрением и в настоящее время. Основы этой теории изложены в ряде
2.6. Основные следствия теории сбалансированного питания
2.6. Основные следствия теории сбалансированного питания Теория сбалансированного питания замечательна не только стройностью и ясностью логики и экспериментальной обоснованностью, но и способностью к предсказанию неизвестных явлений, которые были затем открыты, или
3.2. Основные постулаты теории адекватного питания
3.2. Основные постулаты теории адекватного питания Кризис теории сбалансированного питания и открытие некоторых витальных, неизвестных ранее механизмов (лизосомального и мембранного пищеварения, различных видов транспорта пищевых веществ, общих эффектов кишечной
Глава 1. Теории прогрессивной эволюции
Глава 1. Теории прогрессивной эволюции Кто знает, что дух человека возносится ввысь, А дух скота — тот вниз уходит в землю? Экклесиаст, III, 21* Бытовой антропоцентризмЧеловек может оставаться равнодушным ко многому, только не к собственной персоне. В себе его интересует