О подборе родительских пар, закономерности доминирования и природе гетерозиса в сроках вегетации
Предварительно исследовав закономерности индивидуального развития растительных форм, исследовав необходимые условия и характер индивидуального развития каждого из кандидатов для скрещивания, мы в известной мере узнаём скрытую сущность наследственной основы выбираемых кандидатов. Стоя на позициях развития наследственного основания (генотипа), мы получаем возможность сознательно выбирать родительские пары.
Этот сознательный выбор пар для скрещивания включает в себя возможность:
а) заранее, ещё до скрещивания, предсказывать картину доминирования стадий развития того или иного родителя — отсюда предвидеть и длину вегетационного периода F1 в сравнении с обоими родителями;
б) преднамеренно изменять картину доминирования сроков вегетации;
в) по длине вегетационного периода F1 предвидеть срок вегетации наиболее скороспелых из возможных выщепенцев и соответственно этому производить браковку комбинаций, начиная с F1 ;
г) предсказывать общую картину возможного расщепления гетерозиготы по длине вегетации по данным любого поколения;
д) не ограничиваться случайным появлением гетерозиса и случайным выведением раннеспелых сортов от скрещивания позднеспелых, а преднамеренно создавать их в данных условиях при помощи сознательного выбора родительских пар.
Обычно селекция, оперируя с комбинаторикой отдельных признаков и не зная непосредственной основы развития признаков — закономерностей стадийного развития, лишь случайно и очень редко набредает на нужные комбинации, для получения сорта. И с вегетационным периодом селекция также оперирует просто как с «признаком», упуская, что этот «признак» есть итоговый результат всего развития, непосредственной базой которого является закономерность стадийного развития. Ведь именно длительность течения тех или иных стадий развития, при наличии требуемых стадиями условий существования, равно как задержка перехода данной стадии в последующую, вследствие отсутствия требуемых условий существования новой стадии, определяют в основном длительность вегетационного периода, определяют, будет ли сорт скороспелым яровым, позднеспелым яровым, полуозимым, озимым и т. д. Различнейшие факторы воздействия, неизбежно вмешивающиеся в жизнь растения в поле, конечно, несколько отклоняют в ту или иную сторону срок созревания. Но эти отклонения незначительны и остаются в пределах меры, определяемой закономерностью стадийного развития.
Оперируя со сроками вегетационного периода как с признаком, не зная причин, его определяющих, селекционер не может заведомо знать, какие пары надо ему брать для скрещивания, чтобы получить радикальный перелом в сроке вегетации. Если селекционер из наличного ассортимента сортов погонится за наиболее коротким по сроку вегетации, то он неизбежно потащит в гетерозиготу вместе с коротким сроком ещё «и» плохих показателей. Обычно селекционер так и вынужден делать.
Он берёт на учёт у одного сорта его иммунность к головне, к ржавчине и т. д., а у другого сорта берёт более короткий срок созревания, и в результате, погнавшись за коротким сроком созревания, селекционер втащит в гетерозиготу и подверженность тем или иным болезням или подверженность повреждению гессенской и шведской мушкой, малую устойчивость к засухе или плохие мукомольно-хлебопекарные свойства и т. д. Поэтому селекционер и не может знать заранее, когда получится и получится ли из взятой им комбинации лучший или худший сорт по урожаю. И лишь последующая апробация раскрывает ему картину полученного результата.
Не зная другого выхода и желая охранить зиготу от занесения в неё ухудшения, желая иметь дело с меньшим количеством неизвестных при расщеплении гетерозиготы, селекционеры и выдвинули принцип выведения сорта путём инцухта, но при этом они отказываются от возможностей обогащения наследственной основы, отказываются, например, ликвидировать плохое (длительное) течение стадии яровизации при помощи создания гетерозиготы из скрещивания с другим сортом с хорошим (более коротким) течением той же стадии. Поэтому инцухт в тех случаях, когда нужно создать сорт с более коротким вегетационным периодом, является не только беспомощным, но и вредным.
Мы же исходим из совершенно других принципов при выведении сорта, относительно гарантированного по урожайности в данном районе.
Среди наличного ассортимента сортов надо выбрать для скрещивания такие сорта, которые имеют возможность дать все хорошие показатели по всем стойкостям и урожаю при условии устранения в их наследственной основе невозможности прохождения или длительного прохождения в данных условиях района первой стадии (яровизации) или же второй (световой).
Но как раскрыть эти возможности? Как раскрыть возможную урожайность сорта, если он обычно не созревает или же поздно созревает и даёг плохого качества зерно?
Обычная селекция и формальная генетика, игнорирующие исследование закономерностей индивидуального развития, бессильны проникнуть в скрытые возможности наследственного основания, обеднённо трактуя эти возможности, сводя эти возможности лишь к тем признакам, которые обычно сами собой выносятся на поверхность явлений при географических посевах или же при гибридологическом анализе. Эти скрытые возможности частично можно выявить, заставив их соответственно осуществляться в индивидуальном развитии растений. Это можно сделать искусственно, создав для каждого из родителей отсутствующие в полевых условиях района необходимые условия их стадийного развития. Например, одного из родителей надо яровизировать, а другому создать соответствующие условия освещения. В результате каждый из родителей иначе разовьёт весь свой цикл в репродукции, что даст возможность узнать как весь ход и срок его развития, так и урожайность. Это и будет исследование возможностей наследственного основания родителя, проникновение в скрытые возможности генотипа. Раскрыв возможность сроков созревания и урожайную возможность родителей, при условии непрерывного, без задержки идущего развития их наследственной основы через все стадии к репродукции, установив тем самым единственное в каждом из родителей, но различное узкое место в наследственной основе для данных условий, можно преднамеренно создать гетерозиготу, в которой будут заведомо ликвидированы оба эти узкие места. И если один из родителей является представителем сорта, обладающего всеми хозяйственно ценными свойствами, при условии преодоления его единственного узкого места — плохого приспособления к условиям данного района течения стадии яровизации, то это узкое место можно ликвидировать соответствующим включением в зиготу гаметы другого родителя, обладающего хорошей приспособленностью стадии яровизации и также обладающего всеми хозяйственно ценными показателями, при условии ликвидации его собственного узкого места — плохого приспособления для данных условий района, течения стадии световой. Таким образом, путём скрещивания этих двух родителей будет создана гетерозигота. которая обладает реальной возможностью в условиях данного района развить, и притом быстро, как первую, так и вторую стадии, обладая в то же время всеми другими, в условиях данного района, хорошими показателями родительских гамет. Этот свой подход к выбору родительских пар лаборатория физиологии развития растений Генетико-селекционного института кратко формулирует следующим образом.
Выбирать родительские пары для скрещивания надо не по наибольшему количеству положительных признаков у родителей, а по наименьшему количеству отрицательных, лимитирующих в данном районе урожаи, узких мест задатков приспособления наследственного основания с тем, чтобы в гетерозиготе создать возможности взаимопреодоления узких мест родителей.
В частности, у таких растений, как хлебные злаки, соя, хлопчатник и ряд других, урожай которых в огромном количестве районов лимитируется сроком созревания, нужно выбирать родительские пары, пользуясь всей мировой коллекцией, так, чтобы родители имели не одноимённые, а различные единственные «узкие места» возможности развития наследственного основания (у одного родителя — яровизация, у другого — световая стадия).
Эти единственные узкие места и будут преодолены в процессе расщепления гетерозиготы путём взаимного замещения в наследственной основе плохого показателя одной формы аналогичным хорошим показателем второй, и наоборот.
Создавая зиготу из преднамеренно подбираемых пар, в потомстве которых благодаря расщеплению окажутся формы с прёодолённым «узким местом» их родителей, можно заранее, ещё до всякого скрещивания, знать и предсказывать, каков будет общий характер развития возможностей наследственного основания, какая из возможностей будет осуществлена в действительности и, следовательно, каков будет характер развития первого поколения (F1) по срокам бутонизации у хлопчатника, срокам созревания пшениц и т. д., при этом с большой долей вероятности зная также заранее все прочие хозяйственные показатели.
Если скрестить две формы, взяв одну с коротким вегетационным периодом, а другую с более длинным, взять в скрещивание озимую и яровую пару, яровую и полуозимую, яровую позднеспелую и полуозимую и т. д., то что здесь будет доминировать? Ответить на этот существеннейший вопрос правильно можно лишь с позиций развития, поставив и разрешив вопрос об общей закономерности доминирования.
Ещё Грегором Менделем было подмечено (и это одно из крупнейших завоеваний генетики), что в первом поколении гибридов (F1) из альтернативной пары признаков появляются признаки или от одного родителя, или же от другого. Происходит как бы временное «поглощение» тех или иных признаков родителя «аллелями» другого. Соответственно этому Мендель установил правило доминирования, правило, во многих случаях подтверждаемое последующими наблюдениями и изысканиями в генетике. Но ни сам Мендель, ни все последующие направления в современной генетике не вскрывали, в чём здесь дело, и ограничивались простым констатированием самого факта доминирования.
С позиций же теории развития, указывающей на необходимую роль в развитии внешних условий существования, подчёркивающей роль приспособления в биологическом развитии, можно уяснить необходимость доминирования, понять его биологическую суть и тем самым существенно продвинуться к овладению законом доминирования.
В зиготе, образовавшейся при гибридизации, соединяются два родовых начала, два наследственных основания, внося в зиготу все свои стороны как основу для возможностей развития стадий, которые в свою очередь определяют возможность конкретного образования признаков организма. Но какая же из двух парных возможностей (аллеломорф), полученных от разных родителей, будет осуществлена, перейдёт в действительность, разовьётся?
Это отнюдь не преформируется однозначно в самой зиготе, а решается в зависимости от соответствия внешних условий биологическим требованиям приспособления той или иной стороны зиготы к условиям существования своего развития. Поэтому в F1 разовьётся та из аллеломорфных сторон гетерозиготы, которая найдёт для своего развития соответствующие условия существования. Именно поэтому в данных условиях будет развиваться лишь та сторона зиготы, реализуется лишь та возможность наследственного основания, которая найдёт для себя необходимые условия, окажется лучше приспособленной к наличным условиям существования. Именно это и решает, какая из двух аллеломорфных сторон зиготы разовьётся, то есть окажется доминирующей.
Возражение, которое может быть нам сделано, что в ряде случаев доминируют совершенно определённые признаки, независимо от того, что организмы развиваются в разных средах, мы должны сразу же отвести. Это возражение базируется на неразличении «среды» и «условий существования» в ходе развития. Нужно спросить у растения, а не просто гадать, «как нам кажется», является ли перенос его в другую среду обитания переносом его и в другие «условия существования». Могут быть большие различия в среде, которые оказываются отнюдь не существенными для растения, и в то же время могут быть, казалось бы, незначительнейшие изменения в среде, но если эти изменения являются условиями существования на том или ином этапе жизни растения, то эти изменения обусловят крупнейшие сдвиги в растении.
Рисунок 27 иллюстрирует отличие одних растений ячменя от других. Одни растения колосятся, другие ползут по земле, не имея даже стебля. И в тоже время это один и тот же сорт растения, выращенный на одном и том же поле из обычных неяровизированных семян, и разница лишь в том, что одни растения были высеяны в поле 12 марта 1928 г., а другие тут же рядом на два дня позже (14 марта). Разница в два дня коренным образом Изменила всё поведение растения. Теперь совершенно понятно, в чём здесь дело: одни растения (высеянные 12 марта) попали в требуемые ими условия существования стадии яровизации, так как в течение этих двух дней за весь период вегетации температура была на 2–3° более низкая. Последующие же гораздо большие изменения в температуре остались безучастными элементами среды для развития стадии яровизации.

Рис. 27. Ячмень Паллидум 419.
Посев в поле через каждые два дня, начиная с 1 марта до осени 1928 г. в Гандже. Растения всех посевов, включая 12 марта, оказались яровыми. Растения всех посевов, начиная с 14 марта (растения посева 14 марта на рисунке не представлены), оказались озимыми и выколашивания не дали. Разница в сроке посева на 2 дня (12 и 14 марта) сыграла решающую роль в развитии органов и признаков растений.
Мы же утверждаем, что во всех случаях, когда гибридное растение в ходе развития попадает действительно в другие условия существования это обусловливает и соответствующие сдвиги в доминировании: доминировать будет то, что имеет более благоприятные условия приспособления для своего развития. Ещё раз повторяем, что доминирование отнюдь не преформировано однозначно в зиготе. Зигота, объединяющая обе родительские наследственные основы, содержит в себе возможность развития и одного и другого члена аллеломорфы. Решает же вопрос доминирования приспособленность развития того или иного члена аллеломорфы к наличным условиям существования.
Следовательно, то, что будет доминировать в одних условиях, будет рецессивно в других.
Взгляд на доминирование с позиций развития давно был высказан и претворялся в работу Иваном Владимировичем Мичуриным.
«… качества каждого гибрида, — писал Иван Владимирович, — выращиваемого из семян плода, полученного от скрещивания двух производителей, состоят из комбинации лишь той части наследственно переданных ему свойств от растений-производителей, т. е. отца, матери и их родичей, развитию которых в самой ранней стадии роста гибрида благоприятствовали условия внешней окружающей среды (т. о. температура окружающего воздуха и почвы, степень насыщенности атмосферы электричеством, того или другого направления и силы господствующие ветры, степень освещения, состав почвы, степень ее влажности и т. д.)»[16].
Именно этим взглядом на зиготу, как на нечто развивающееся, и на доминирование, как на развитие лишь тех сторон зиготы, которым благоприятствовали наличные условия, и руководствовался во всей своей огромной и плодотворнейшей работе Иван Владимирович: именно эти теоретические предпосылки дали ему возможность вывести огромное количество новых сортов и небывалых форм растений. Исходя из этих теоретических предпосылок, Иван Владимирович и выдвинул как один из своих принципов работы принцип отдалённой гибридизации. Но этот принцип Мичурина в корне отличается от установок теперешних отдалённых гибридизаторов. Мичурин выдвигал отдалённую гибридизацию не просто ради увеличения многообразия растительных форм независимо от того, что собой эти формы будут представлять, занимательную ли игрушку или же дело, рассчитывая лишь, что «авось» получится когда-либо и что-либо полезное. Он вовсе не предлагал скрещивать просто любое с любым, лишь бы пары были подальше в видовом и географическом отношениях.
И. В. Мичурин сознательно подбирал родительские пары, в том числе в значительной степени отдалённые как друг от друга, так и от места будущего назначения сорта, исходя из глубочайшего учёта различия их условий существования. И делал он это совершенно конкретно для того и в тех случаях, когда ему необходимо было парализовать развитие определённых свойств, доминирующее благодаря приспособленности к местным условиям развития, и создать условия доминирующего развития планово намеченных Иваном Владимировичем свойств выводимого им сорта. Подбирая для скрещивания наследственные основы, Мичурин всё время держал в голове учёт их исторически сложившихся биологических требований приспособления, прикидывая заранее, как пойдёт развитие наследственной основы в определённых условиях существования и при определённых факторах воздействия, и подбирая заранее эти условия для получения нужной картины доминирования, то есть заранее рассчитывая свою работу в отношении всего пути многопротиворечивого развития наследственной основы в признаки. В пояснение своих установок Иван Владимирович приводит следующий пример его работы как один из огромного количества подобных случаев:
«При скрещивании иностранных сортов зимних груш с нашими Тонковетками, Лимонками и другими выносливыми сортами получались гибриды, хотя и с лучшими вкусовыми качествами, но все с летним созреванием и мелкой величиной плодов, что произошло от доминирующего развития признаков наших местных сортов вследствие подходящих и привычных для них климатических и других условий нашей местности. Напротив, когда сделано было мною скрещивание иностранных зимних груш с дикой уссурийской грушей (Pyrus ussuriensis Max.), которая была у меня выращена из семян, полученных из Северной Манчжурии, тогда получились гибриды в одной половине своего количества с крупными плодами прекрасного вкусового качества со свойством зимнего созревания в лежке и с полной выносливостью к нашим морозам всех надземных частей деревьев»[17].
Иван Владимирович формулирует эту свою теоретическую установку следующим образом:
«Чем дальше отстоят между собой пары скрещиваемых растений-производителей по месту их родины и условиям их среды, тем легче приспособляются к условиям среды в новой местности гибридные сеянцы. Я объясняю это тем, что в данном случае наследственно переданные гибридам свойства отца или матери и их ближайших родичей, не встречая привычных для них как на родине условий среды, не будут в состоянии слишком сильно доминировать односторонней передачей этих свойств в развитии организма гибридов, что имеет огромное значение в деле»[18].
Взгляд на доминирование как на развитие, обусловленное большей приспособленностью к условиям, — вот что выдвигал и чем руководился Иван Владимирович и вот что явилось одной из его теоретических установок, давших ему могучее оружие и орудие в борьбе и действии.
Эту свою установку Иван Владимирович выдвигал очень давно, противопоставляя её формальному пониманию доминирования, как однозначно и вне зависимости от условий приспособления решаемому в самой зиготе. Ещё в ранних его работах он выступает как борец против такого формализма.
«Все наши менделисты, — писал И. В.[19], — как кажется, не желают принимать в расчет громадную силу влияния таких факторов (И. В. ранее в этой же статье перечисляет главнейшие из них: «… давление воздуха, состояние температуры, количество влаги, силы света и электричества; как каждый в отдельности, так и в различных комбинациях.») на сложение формы построения организма растения гибрида, начиная с момента образования семени от скрещивания двух особей до истечения нескольких лет роста сеянца гибрида, т. е. до его полной возмужалости. Примером чего приведу следующий факт: мною было произведено оплодотворение цветов Pyrus elaeagnifolia пыльцой с известного садового сорта груши Бессемянки. При культуре всех этих сеянцев мною замечено, что как только сеянцы попадали в лучшие условия питания, так наружный вид всех частей этих растений неизменно уклонялся в сторону Бессемянки. Листовые пластинки получали более широкую форму и блестящую поверхность, побеги утолщались, и кора их получала окраску, приближающуюся к виду побегов Бессемянки. Напротив, если сеянцы подвергались какому-либо страданию, например, при пересадке или при засушливом лете от недостатка влаги в начале вегетационного периода, то листья гибридов вырастали узкой и длинной формы. Те же явления наблюдались и на гибридах других комбинаций пар скрещивания культурных сортов с дикими видами растений. В испытаниях влияния состава почвы на сложение выращиваемых гибридов я также делал многочисленные опыты и всегда убеждался в том, что сила влияния этого фактора довольно значительна. В особенности это резко проявлялось в тех случаях, когда удавалось подставить гибридным сеянцам почву, близкую по составу к той, на которой успешно развивалась в течение долгого периода времени или, так сказать, выработалась форма одного из растений — производителей скрещенной пары, между тем как форма другого растения из этой же пары создавалась на почве совершенно иного состава. Во всех почти таких случаях гибридные сеянцы уклоняются своей формой в сторону первого производителя. Например, для воспитания гибридов, полученных от скрещивания нашей степной самарской вишни Prunus chamaecerasus с владимирской Родителевой вишней, я нарочно выписывал из окрестностей г. Владимира несколько пудов земли, на которой там растут всем известные Родителевские сладкие вишни. И несмотря на то, что этой подставкой почвы я мог только отчасти приблизить условия среды воспитания гибридов к таковым же условиям роста Родителевой вишни на ее родине, но тем не менее, те немногие экземпляры из общего числа гибридных сеянцев, которым дана была почва в сильной смеси с полученной из Владимира землей, сильно уклонились в своей форме в сторону Родителевой вишни и резко отличались от остальных сеянцев, воспитанных на нашей обыкновенной почве. Но ведь в этом опыте многого недоставало, — воспитывать сеянцы нужно было не в Козлове, а во Владимире, именно на родине Родителевой вишни, потому что в данном случае, кроме определенного состава почвы, большую роль играют и состав подпочвы, состав подпочвенной воды, расстояние уровня последней от поверхности почвы, положение местности, разница климатических условий и т. п. И если при отсутствии влияния таких существенно важных деятелей и при лишь одной подставке родной почвы сеянцы могли так заметно отклониться в сторону того или другого материнского растения, то как же можно определять величину или численность их отклонения в сторону родительского растения на основании только одной наследственной передачи свойств последней?»
Мы видим, что Иван Владимирович направлял развитие доминирования как подбором пар с определёнными требованиями приспособления к условиям развития, так и путём создания условий существования, требуемых развитием в нужном направлении. И именно практическая реализация теоретических позиций, исходящих из развития наследственной основы в условиях существования, привела к таким огромным достижениям в деле выведения сортов Иваном Владимировичем.
Мы здесь вынужденно бегло, лишь по ходу нашего изложения, остановились на одном из моментов теоретических установок и предпосылок работы Мичурина. Исчерпывающего же изложения всего огромного богатства их мы здесь, понятно, не можем дать.
Одно только можно сказать: Мичурин руководился поисками закономерности развития растений на всём протяжении жизни растения и нашёл путь к овладению многими из этих закономерностей.
Понимание доминирования как развития в противовес всей официальной нормальной науке и сознательное направление доминирования — этого одного уже достаточно, чтобы отсечь рассуждения некоторых «теоретиков» о научной «незаконнорожденности» работ Мичурина.
Настоящая наука о гибридизации — у Мичурина. Но не каждому дано его понять. Для этого надо подлинно стоять на позициях материалистического развития.
Из однотипных по своей природе и по требованиям к условиям существования, но различных по конкретному характеру требований к однотипным условиям существования возможностей развития наследственного основания (аллеломорф), действительно может развиться лишь одна из этих однотипных возможностей (короткая или длинная стадия — «яровизации», короткая или длинная стадия — «световая» и т. п.). Ведь речь идёт не о «доминировании» чуждых по своей природе возможностей развития, не о «доминировании» длинного над чёрным, жёлтого над коротким и т. п.; речь идёт о парных и однотипных возможностях (длинное — короткое, круглое — морщинистое и т. д.), поэтому понятно, что в этих случаях реальное развитие этих возможностей может быть лишь взаимоисключающим. И разовьётся именно та из взаимоисключающих друг друга возможностей, которая найдёт в окружающих условиях большее благоприятствование требованиям осуществления её развития, окажется лучше приспособленной к развитию в наличных условиях существования. Поэтому приспособленность или неприспособленность гетерозиготы к развитию в данных условиях обязательно скажется уже в F1; если гетерозигота имеет возможность полноценно развиться в данных условиях района, то эта возможность осуществится в первом поколении. Это и обусловит то или иное доминирование в развитии возможностей наследственного основания.
Поняв доминирование как развитие одной из парных возможностей гетерозиготы, благодаря большему соответствию требованиям, большей приспособленности развития этой возможности зиготы к наличным условиям существования, можно заранее, ещё до скрещивания, знать картину доминирования в отношении тех или иных «аллеломорфных» задатков (возможностей). Для этого надо заранее изучить возможности развития наследственного основания родителей; установить эти возможности, найдя условия, требуемые каждым из родителей для своего стадийного развития и для развития на этой базе органов и признаков растения; установить, какому же из этих требований более соответствуют наличные условия места высева (условия района).
Стадийный анализ, таким образом, должен предшествовать как гибридологическому анализу, так и самой гибридизации. Лишь тогда гибридологический анализ не будет строиться на слепых находках выщепления и можно будет управлять доминированием «признаков», как теперь уже стало возможным в основном управлять теми из них, которые являются непосредственным результативным итогом стадийного развития. Именно таким признаком и является срок вегетации. При продвижении дальше в анализ условий существования процесса формирования органов и их признаков, развитие которых является частной формой существования общих закономерностей стадийного развития, создаётся всё большая и большая возможность управления доминированием.
Общую закономерность доминирования можно сформулировать следующим образом.
Доминирование есть развитие той или иной аллеломорфной стороны наследственного основания (гетерозиготы), требованиям которой соответствуют наличные условия существования при невозможности развития, из-за отсутствия требуемых условий или меньшего благоприятствования их для развития другой парной стороны гетерозиготы.
Но так как та или иная сторона наследственного основания является лишь реальным носителем возможности развития, то эта же закономерность доминирования может быть сформулирована следующим образом.
Доминирование есть превращение в действительность одной из парных (аллеломорфных) и взаимоисключающих друг друга возможностей развития наследственной основы благодаря наличию соответствующих условий или меньшей приспособленности к ним осуществления развития другой аллеломорфной возможности.
Исходя из этой закономерности, можно заранее, ещё до скрещивания, знать, каково будет доминирование сроков созревания при скрещивании пары озимой и яровой, пары с коротким и длинным вегетационным периодом, полуозимой и яровой и т. д.

Рис. 28. На рисунке представлены растения двух позднеспелых (в условиях района Одессы) яровых сортов пшеницы Hordeiforme 2508 и Melanopus 069.
Предпосевная яровизация семян Hordeiforme 2508 в условиях района Одессы ускоряет выколашивание растений. Следовательно, данный сорт в условиях района Одессы позднеспелый из-за медленного прохождения стадии яровизации. Первый снопик слева из яровизированных семян, растения выколосились, второй — из обычных. Предпосевная яровизация семян сорта Melanopus 069 не ускоряет выколашивания его растений. Растения этого сорта ускоряют своё выколашивание при удлинении дня. Следовательно, этот сорт в условиях района Одессы позднеспелый из-за медленного прохождения световой стадии. Путём гибридизации обеих этих форм создаётся наследственная основа, в которой ликвидируются оба узких места этих родителей. Стадийный анализ обеих родительских форм дал возможность ещё до скрещивания предвидеть характер развития их потомства как скороспелого. См. выколосившиеся растения первого гибридного поколения (F1) — второй снопик справа.

Рис. 29. Стадийный анализ Hordeiforme 2506 и Melanopus 069 показал, что из этих двух позднеспелок получится раннеспелый гибрид (см. снопик второй справа); у Hordeiforme длительное (в условиях района Одессы) течение стадии яровизации, у Melanopus — длительное течение стадии световой.
Путём гибридизации создаётся наследственная основа, в которой есть возможность быстрого прохождения обеих этих стадий (в условиях района Одессы).
Для этого надо произвести стадийный анализ, то есть уяснить, по какой причине, благодаря каким стадиям родители развиваются как озимые, яровые, раннеспелые и т. д. формы. Выяснив, какие условия требуются развитием каждой из этих стадий у каждого из родителей для того, чтобы их развитие шло более быстро и непрерывно из стадии в стадию, можно, изучив условия района, заранее знать, как разовьётся гетерозигота в F1 в озимую, яровую форму, полуозимую, позднюю, раннюю форму и т. д., и соответственно действовать как в подборе пар для скрещивания, так и условий существования (выбор района) для получения нужной картины доминирования по сроку созревания первого поколения (рис. 28 и 29).
При скрещивании двух сортов, из которых один в данных условиях существования (условия района)развивается как озимый, а другой как яровой, получается гетерозиготное наследственное основание, в котором есть реальная возможность ярового развития. И гибридное семечко, будучи высеяно весной, найдёт необходимые условия существования для осуществления развития наличествующей в наследственной основе «яровой» возможности.
«Яровость», таким образом, будет доминировать над «озимостью». Но при этом всегда надо помнить, что сама яровость любого из родителей будет сохраняться лишь в данных условиях существования, а в других она станет «озимостью». Поэтому первое поколение, от скрещивания тех же родителей, развиваясь в других условиях, может получиться озимым.
Избежать же такой ошибки можно лишь при помощи предварительного стадийного анализа скрещиваемых родителей. Необходимо ещё до скрещивания выявить, какие условия существования для стадии яровизации требует каждый из родителей. После этого можно будет предвидеть, будет ли первое поколение озимым или яровым при посеве в любом районе, а не только в том районе, где выращивались родители.
Всегда, скрещивая двух родителей, из которых хотя бы один в данных условиях развития будет яровым, мы заранее знаем, что в этих же условиях развитие гибрида F1 будет также яровым, то есть знаем картину доминирования заранее.
Если для скрещивания взять полуозимую форму и яровую, то по тем же соображениям наличия в гетерозиготе реальной возможности «ярового» развития в данных условиях существования, то есть при посеве в тёплое время года, F1 будет яровым, то есть «яровость» будет доминировать над «полуозимостью».
Если для скрещивания взять две формы, из которых одна скороспелая, а другая позднеспелая в данных условиях района, то так как в гетерозиготе будет налицо реальная возможность раннеспелого развития при наличных условиях и так как сами условия налицо, то и само развитие гибрида F1 пойдёт как раннеспелое. Таким образом, раннеспелость будет здесь доминировать над позднеспелостью.
Соответственно общей закономерности доминирования можно сформулировать следующую закономерность в сроках вегетации, являющуюся конкретизированием общей закономерности доминирования.
У первого поколения гибридов (F1) от двух родителей, из которых один в данных условиях будет рано созревать, а другой в этих условиях будет более позднеспелым, доминировать в этих же условиях будет раннеспелость. Первое поколение всегда будет столь же раннеспелым, как наиболее раннеспелый из родителей, или же ещё более раннеспелым (табл. 1 и 2 на стр. 80–81).
Случаи большей раннеспелости F1 нежели оба гомозиготных родителя, давно известны современной генетике и, по нашему мнению, имеют общую природу со случаями более мощного роста у F1, нежели у обоих родителей, являясь одной из форм гетерозиса. Но формальная генетика, не анализирующая закономерностей развития признаков на основе стадий развития, бессильна этот гетерозис объяснить.
С позиций же понимания стадийности развития гетерозис скороспелости становится понятным, и это понимание даёт нам возможность предсказывать такой гетерозис и преднамеренно его создавать. Мы исходим из того, что гетерозис скороспелости — это суммарное результативное выражение доминирования в развитии нескольких более быстро протекающих стадий при гибридизации.

Рис. 30. Создание гетерозиса скороспелости путём подбора пар на основе стадийного анализа у пшениц.

Рис. 31. Создание гетерозиса скороспелости путём подбора пар на основе стадийного анализа у ячменя.
Паллидум 883/6 Азербайджанской станции не приспособлен в условиях Одессы пройти при весеннем посеве стадию яровизации (см. снопик слева). Паллидум 046 Одесской станции (см. снопик справа) плохо приспособлен к этим условиям весеннего посева в отношении световой стадии. Гибридизация этих форм даёт гетерозис раннеспелости.
Гибридизируя две наследственные основы — одну с более благоприятной (быстро протекающей) в данных условиях стадией яровизации при плохой (затяжной) стадии световой и вторую с более благоприятной в этих же условиях развития световой стадией при плохом (затяжном) течении стадии яровизации, мы, вследствие закономерности доминирования, с необходимостью будем иметь в F1 гетерозис скороспелости.
Так, если взять две позднеспелые формы, произвести стадийный анализ каждой из них, и если они окажутся позднеспелыми благодаря разным стадиям, то при скрещивании их образуется наследственное основание, в котором есть в наличии реальная возможность более быстрого развития и одной и другой стадии. И в данных условиях существования развитие гибрида от двух позднеспелок пойдёт как раннеспелое.
Таким образом, подбирая родителей с учётом различия условий существования и развития их стаций, можно преднамеренно создавать гетерозис раннеспелости (рис. 30–33).

Рис. 32. Кунжут.
Оба родителя с длинным вегетационным периодом. Стадийный анализ показал, что их позднеспелость обусловлена разными стадиями. Подбор пары на основе стадийного анализа показал возможность из этих двух позднеспелых форм создавать раннеспелую форму.
Стадии развития, как мы уже указывали, образуют общую биологическую закономерность в индивидуальном развитии растения от семени к семени, закономерность, отражающую в требованиях развития стадий к своим условиям существования историческое действие естественного отбора приспособлений.
Эти стадии, различная природа требований которых к условиям своего существования у различных органических форм имеет своей базой исходное различив наследственных основ этих форм, являются сами базой, из которой развиваются органы с их конкретными признаками. Но эти последние, равно как и их база (стадии), в свою очередь развиваясь при своих условиях существования, конкретно развиваются при неизбежном влиянии различнейших, иногда не сразу поддающихся учёту, внешних воздействующих факторов, которые являются далеко не безучастными в формообразовании признаков у каждого отдельного организма как единичного представителя сорта, обусловливая те или иные конкретные отличия в формообразовательном процессе.
Быстрота и срок прохождения стадий хоть и определяют быстроту и срок наступления выхода в трубку, бутонизацию, цветение, плодоношение и т. д., но далеко не нацело. Поэтому изменения в сроках прохождения стадий и изменения сроков бутонизации, цветения, плодоношения и т. д. с необходимостью будут несколько расходиться, причём у различных растений это расхождение будет различно, определяясь специфической природой каждой растительной формы и спецификой развития его органов, а также различным воздействием внешних факторов. Но при всём этом общая закономерность зависимости сроков бутонизации, цветения и плодоношения от сроков прохождения стадий развития сохраняется.
Имея всё вышеизложенное в виде теоретических предпосылок, лабораторией физиологии растений Генетико-селекционного института и были поставлены опыты как в полевых, так и в лабораторных условиях. Полученные результаты целиком подтверждают правильность этих теоретических позиций, целиком подтверждают правильность сформулированной нами закономерности доминирования в сроках вегетации (табл. 1 и 2).
Опираясь на вышеизложенную закономерность доминирования сроков вегетации, стало возможным производить браковку комбинаций уже по первому гибридному поколению, обычно игнорируемому селекционерами. Из этой закономерности доминирования вытекает, что наилучшая картина развития (в смысле скороспелости) будет в F1, имеющем наиболее благоприятное основание по наибольшему богатству возможностей развития, нежели все последующие поколения.

Рис. 33. Создание преднамеренного «гетерозиса» раннеспелости путём подбора стадийно проанализированной пары для скрещивания (из мировой коллекции).
Приспособленность той или иной возможности наследственного основания к наличным условиям, если она в действительности имеется, неизбежно должна сказаться уже в F1 и если F1 не смогло в данных условиях развития полноценно развиться, то лучшего течения развития (в данном случае более короткого срока созревания) у последующих поколений с обеднёнными, в следствие расщепления, возможностями наследственного основания ожидать не приходится.
Так что, если в F1 получилась форма, не удовлетворяющая лимиту требований к сорту для данного района по сроку созревания, то дальнейшая селекционная работа по получению выщепенцев из данной комбинации становится ненужной.
Ни в одном из следующих поколений не может развиться более скороспелая форма, нежели само F1.
Произведённые опытные работы по гибридизации, доведённые сейчас уже до F7, целиком, без одного исключения, подтверждают это.
Из этого же понимания наследственного основания как носителя возможностей развития вытекает, что во всех последующих поколениях может происходить обеднение возможностей раннего развития благодаря выщеплению, но никогда не может происходить их обогащение (без мутаций). Поэтому F2 не может быть более скороспелым, нежели F1; F3 не будет более скороспелым, нежели F2, и т. д.
Таблица 1. Посев 26 марта 1934 г. в поле
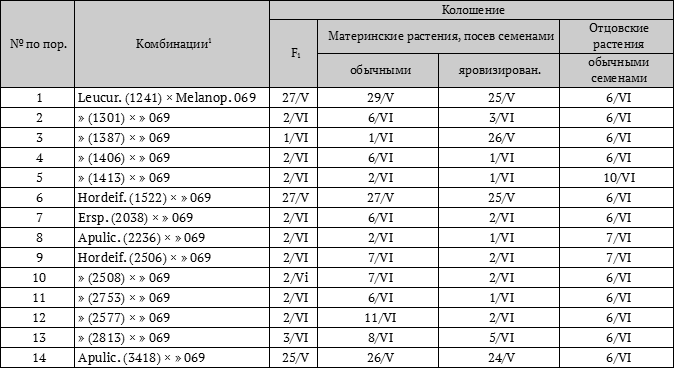
1 Номера в скобках указаны по каталогу ВИРа.
Таблица 2. Посев 24 августа 1933 г. в вегетационном домике
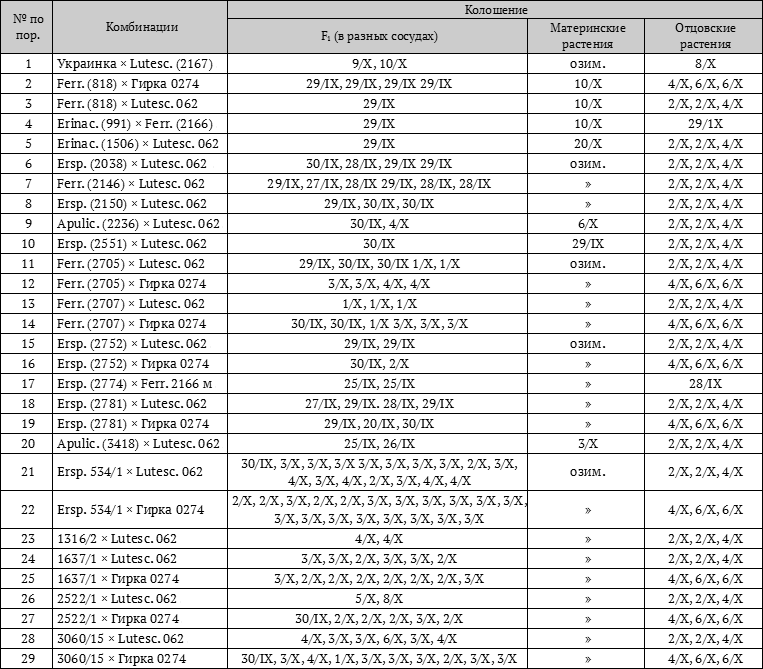
Все результаты опытов лаборатории физиологии развития растений и просмотренные в этих целях литературные данные целиком подтверждают это положение.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК