О двух направлениях в генетике (1937)
Доклад на IV сессии Академии сельскохозяйственных наук им. В. И. Ленина 23 декабря 1936 г.
Товарищи, в заслушанных здесь докладах селекционеров, работающих в разных местах нашего великого Советского Союза, была показана огромная творческая селекционная работа, ведущаяся с самыми разнообразными растительными культурами и породами животных. Успехи этих работ колоссальны. Нет никакого сомнения также и в том, что здесь, на сессии Сельскохозяйственной Академии, была представлена лишь ничтожная доля всей той огромной селекционно-генетической работы, которая ведётся у нас в Союзе ССР.
В нашей стране наукой овладевают широчайшие слои трудящихся, начиная с работников многих тысяч колхозных хат-лабораторий и кончая работниками исследовательских институтов и академиками. Сравнивать наши достижения с исследовательской работой в царской России никому даже не придёт в голову.
Во многих разделах наша сельскохозяйственная наука уже вышла на передовые позиции в мире.
Исходя из безусловных успехов селекционно-генетической науки, некоторым товарищам, в том числе и некоторым присутствующим здесь на сессии Академии, непонятны причины и корни той дискуссии, которая ведётся сейчас на страницах журналов «Социалистическая реконструкция сельского хозяйства» и «Яровизация». Некоторые из дискуссирующих в журналах выступают в довольно приподнятых тонах, с нередкими, на мой взгляд, перегибами, со стремлением подтасовать факты в выгодном для себя направлении. Лично к себе я этого отнести не могу. Я думаю, что тот, кто следил за печатью, должен придти к заключению, что мои статьи хотя и являются страстными, но во всяком случае беспристрастны (аплодисменты). Статьи же Дончо Костова, академиков Константинова, Лисицына, М. М. Завадовского и некоторых других, мне кажется, действительно, нестрастны, хладнокровно размеренны, но зато сугубо пристрастны.
Дискуссия, которая у нас велась и ведётся, не является простым столкновением, мнений отдельных учёных, — она затрагивает, важнейшие интересы исследовательской работы. Этим только я и объясняю, почему эта дискуссия, ведущаяся как будто в узко научной области — в селекции и генетике, — вызвала такой большой интерес у широкой советской общественности, в том числе и у колхозников-опытников. Дело здесь идет не о частных мелких вопросах; дело идёт о главной линии направления работ в агробиологической науке. Основным, на чём сейчас заостряется внимание в нашей дискуссии, являются взгляды на процесс эволюции растительного и животного мира.
Чем лучше будут поняты закономерности развития растительных и животных форм, тем легче и быстрее мы сможем по своему желанию и заданию создавать нужные нам формы.
Материалистическое ядро эволюционного учения Дарвина в своей основе является революционным, действенным. Естественным и искусственным отбором Дарвин блестяще объяснил природные целесообразности животного и растительного мира.
Дикая растительность, главным образом, отличается признаками и свойствами, полезными не для удовлетворения потребностей человека, а признаками и свойствами, полезными для самого вида, рода растения с точки зрения его приспособленности, лучшей его выживаемости в тех условиях, в которых данное растение произрастает. Растения культурных сортов создаются людьми, поэтому они и пригоняются соответственно к потребностям человека.
Человек отбирает на семена (на племя) только те растения, которые наиболее соответствуют поставленной им задаче. Конечно, те изменения, благодаря которым культурное растение в данных условиях но может выживать, приводят его к гибели, то есть в этих случаях и у культурных растений действует естественный отбор. Но мы знаем, что, наряду с отбором растений на семена с желательными для человека изменениями, он же (человек), соответственно изменившимся растительным организмам, изменяет и условия культуры, изменяет агротехнику. Отсюда, чем выше, чем интенсивнее культура данного растения, тем всё больше и больше увеличивается роль искусственного отбора в создании новых форм и всё больше и больше уменьшается роль и значение естественного отбора.
Дарвин на основе обобщения громадного опыта человеческой практики, увязав это с наблюдениями за растениями и животными в естественной природе, создал своё эволюционное учение. Он показал, что растительный и животный мир изменяются. Дарвин открыл причины наблюдаемой нами пригнанности организмов к условиям среды обитания, а у культурных организмов, кроме того, ещё и пригнанность форм для удовлетворения потребностей человека. Этим самым учение Дарвина развязывает людям руки, инициативу для действия, для создания новых форм растений и животных.
Гениальное учение Дарвина в буржуазных странах не получило и не могло получить настоящего расцвета и дальнейшего развития. Лучшие учёные дарвинисты в капиталистических странах, как, например, Бербанк в Америке, так же как и наши революционеры биологии К. А. Тимирязев и И. В. Мичурин в царской России, были борцами-одиночками.
Основные нападки на дарвинизм с самого начала его появления велись именно по линии отрицания созидательной роли отбора как естественного, так и искусственного.
Для сокращения времени не буду подробно останавливаться па первых этапах борьбы за дарвинизм. Дарвинизм в капиталистическом обществе прокладывал себе дорогу в жесточайших боях.
Критики из раздела биологической науки — генетики, не имея возможности опровергнуть дарвинизм по существу, всегда, как правило, старались фальсифицировать его, часто под видом исправления неточных методов работы Дарвина или ссылкой на неточность методов научных работ во времена Дарвина.
Дефризианцы противопоставили эволюционному учению Дарвина свою теорию мутаций (как будто бы Дарвину не были известны скачкообразные изменения!!). Известно также отношение к дарвинизму со стороны Бэтсона, Лотси. Известно также отношение Иогансена, давшего учение о чистых линиях самоопылителей. Учение о чистых линиях самоопылителей Иогансена в корне отрицает центральный пункт эволюционного учения Дарвина, а именно творческую роль искусственного и естественного отбора.
Можно было бы привести ряд выдержек из книги Иогансена «Элементы точного учения об изменчивости и наследственности», где он отвергает созидающую роль естественного и искусственного отбора. Ограничусь только одной цитатой: «Для науки о наследственности, как биологической дисциплины преимущественно аналитического характера, было бы лучше всего не переплетать с текущей исследовательской работой воззрений Дарвина и других классиков эволюционной теории»[24].
Этим самым Иогансен говорит, что учение Дарвина к теории наследственности и изменчивости, то есть к тому предмету, которым должна заниматься генетика, никакого отношения не имеет. Мы же, работники советской агронауки, хорошо знаем, что любая наша исследовательская работа в любом направлении изучения растительного организма должна быть насквозь пронизана дарвинизмом. Мы хорошо знаем отношение к дарвинизму лучших учёных биологов-селекционеров, давших миру огромное количество прекрасных сортов. Не буду сейчас говорить о И. В. Мичурине, — о нём я уже много раз говорил. Вспомним величайшего селекционера Америки, покойного Лютера Бербанка. В книге «Жатва жизни» Бербанк говорит:
«Моя приверженность в течение всей моей жизни к учению Чарлза Дарвина не была результатом слепой веры в его авторитет; некоторые из его теорий я даже взял, вследствие моего небольшого опыта, сперва под сомнение.
Но со временем у меня всё больше было случаев практически проверить его теорию в саду и в поле, и, чем старше я становился, тем крепче я убеждался, что он действительный учитель.»[25].
В той же книге (стр. 168) Бербанк рассказывает, как он советовал одному молодому человеку, интересовавшемуся закономерностями в наследственности растительных организмов, подбирать литературные источники для изучения этих закономерностей.
«Я советую вам начать изучать Менделя чтением Дарвина, затем покончить с Менделем и почитать Дарвина более основательно».
Этой цитатой я только хотел подчеркнуть, как Лютер Бербанк в своей глубоко творческой работе ценил учение Дарвина о развитии растительных форм. Далее Бербанк пишет: «Я давал такой совет, потому что я видел, что утверждения многих известных ученых не подтверждались и с ними практически я не мог ничего предпринять, тогда как у Дарвина, как я убедился, всегда все совпадало с фактами и никогда он не бродил в темноте, упрямо следуя за какой-нибудь любимой теорией или из-за предвзятого мнения, и не шел по неверному пути» (стр. 169). Заподозрить Бербанка, что он не специалист, что ему не нужно было знать закономерностей развития растительного организма, я думаю, никто но может.
Бербанк много раз указывает на творческую роль естественного и умелого искусственного отбора. Иогансен же творческую роль отбора отрицает. Мне могут сказать: «Позвольте, Иогансен пришёл к этим выводам на основании точного эксперимента». В том-то и дело, что, на наш взгляд, эксперимент Иогансена неубедителен. Его эксперимент, описание которого переносится из одного учебника в другой, состоял в следующем: была взята фасоль определённого сорта и по крупности зёрен была разделена для посева. В урожае обнаружилось, что этот сорт по своей наследственной природе неоднороден. Он состоял из разных биотипов. Отдельные биотипы, вернее, их потомство, Иогансен назвал чистыми линиями.
В этой части своего эксперимента Иогансен ни в какое противоречие с эволюционным учением Дарвина не вступил. Он только лишний раз подтвердил, что, отбирая для посева крупные зёрна, в урожае будут также более крупные зёрна, нежели от высеянных в этих же условиях более мелких семян.
Вся оригинальность вывода Иогансена основывается на дальнейших опытах с той же фасолью. Он пришёл к выводу, что в посеве растений-самоопылителей, в прошлом происходящих из урожая одного растения и в дальнейшем не подвергавшихся перекрёстному опылению, отбор растений на семена роли не играет. Независимо от того, будет ли выбираться на племя лучшее или худшее растение, всё равно при посеве в одних и тех же условиях будет получаться качественно и количественно одинаковый урожай.
К этому хотя и оригинальному выводу, но противоречащему не только учению Дарвина об отборе, но и обычной сельскохозяйственной практике, Иогансен пришёл на основании шестилетнего опыта.
Почему же результаты шестилетнего опыта с отбором для посева крайних вариантов (наиболее крупных и наиболее мелких семян) у Иогансена встали в противоречие со всей сельскохозяйственной практикой, всегда успешно применяющей отбор на семена (на племя) лучших экземпляров?
Дарвин приводил многочисленные примеры, как путём отбора люди всё больше и больше улучшают породы сельскохозяйственных животных и растений. Может быть, Дарвин был неправ, может быть, люди в своей практике просто заблуждались, да и теперь заблуждаются, отбирая на семена лучшие растения? Но, прежде чем подумать так о мировой сельскохозяйственной практике и заподозрить ошибку в обобщении, сделанном непревзойдённым мыслителем — биологом Дарвином, необходимо уяснить себе, на основе какого же материала Иогансен пришёл к выводу о бесполезности отбора у растений-самоопылителей, происходящих в прошлом из одного растения и не подвергавшихся в дальнейшем скрещиванию.
Изложим кратко суть опытов Иогансена. Он взял урожай отдельного растения фасоли, отобрал несколько зёрен наиболее крупных, несколько зёрен наиболее мелких и отдельно их высеял. Оказалось, что размер зёрен нового урожая из посева крупными и мелкими фасолинами в среднем был одинаков. Из урожая всех растений вместе взятых, выросших из крупных семян, он опять отобрал несколько крупных фасолин для посева, а из второго варианта растений, выросших из мелких семян, он отобрал несколько более мелких зёрен для посева.
Весь шестилетний опыт в этом и заключался. Оказалось, что и на шестой год опыта отбор не дал ни положительных, ни отрицательных результатов. Средний размер зёрен и первого и второго варианта был одинаков.
За шесть лет посева, в опыте с линией фасоли № 1, в урожае обоих вариантов (посев крупных и мелких семян) Иогансеном было получено всего 1 525 зёрен. Следовательно, с каждого варианта за все годы опытов было получено в урожае немного меньше 800 зёрен, а ежегодно в среднем получалось примерно по 150 зёрен. Другими словами, для посева ежегодно отбиралось в среднем всего-навсего от 2 до 5 крупных зёрен и такое же количество мелких семян. Отбор производился исходя не из растений, давших в среднем наиболее крупные или наиболее мелкие семена, в сравнении с другими растениями, развивавшимися в этих же условиях, а выбиралось 2–5 зёрен из смешанного урожая от всех растений данного варианта.
Известно, что не только семена на одном и том же растении, но даже зёрна одного и того же боба неодинаковы по своим размерам. Известно также, что разница в размерах зёрен не всегда говорит о разнице природы зародышей этих зёрен, из которых разовьются будущие растения.
Поэтому Иогансен, по-своему проверяя установленную Дарвином творческую роль отбора, на наш взгляд, мог придти и к другому, ещё более печальному для дарвинизма, выводу, а именно: отбирая мелкие семена для посева, в урожае могли получиться в среднем более крупные семена, нежели в урожае от посева более крупных семян.
Ведь в этом случае, отбирая всего два крупных семени из смешанного урожая, могли же они (или одно из них) случайно быть представителями природы в среднем наиболее мелкосемянных растений, а отбирая два мелких семени, случайно можно было наскочить на природу наиболее крупносемянных растений. Такой случай вовсе не исключён, если для посева отбирают всего два-пять зёрнышек, да ещё из смешанного урожая разных растений, а не исходят при отборе из растений (а не зёрен) крайних вариантов.
Если бы Иогансен отобранные линии фасоли размножил до более значительных размеров, ну хотя бы до центнера, и применял отбор крайних вариантов, учитывая условия развития отбираемых растений, или хотя бы применил отбор в значительно большем масштабе, то он во всяком случае не пришёл бы к выводу, говорящему о бесполезности отбора у растений-самоопылителей, происшедших в прошлом из одного семени.
При большем количестве растении больше шансов, что отдельные индивидуумы попадут в резко отличные условия развития, а это нередко может отражаться на изменении их наследственной природы. Применяя умелый (а не всякий) искусственный отбор в таких посевах, человек в своей практике не только удерживает длительное время хорошие сорта и породы животных, но из года в год улучшает их.
Слепое, некритическое использование учения Иогансена о чистых линиях не может давать положительные результаты. И все лучшие селекционеры мира, даже те из них, которые в теории как будто бы поддерживали учение Иогансена о чистых линиях, в своей практической работе поступали не по-иогансеновски.
Никогда нельзя забывать об отборе. Всегда у всех растений необходимо отбирать на племя, на семена только лучшее. И дарвиновские идеи об отборе но только не нужно забывать, но их нужно всё больше и больше внедрять в сознание миллионов наших колхозников. Лучшие колхозники-стахановцы, как нам хорошо известно из периодической печати, не только применяют отбор на семена лучших растений, но даже занимаются таким кропотливым трудом, как отбор семян по зёрнышку.
Своих оппонентов я знаю довольно хорошо. Они мне скажут: «Теперь генетики думают и поступают не по Иогансену, поэтому напрасно Лысенко ломится в открытую дверь. Генетики не отрицают творческой роли отбора». Именно это и вынуждает меня привести выдержку из книги современного генетика, всеми нами уважаемого Томаса Моргана «Экспериментальные основы эволюции»[26]. На странице 76 читаем: «Современники Дарвина принимали, что путём отбора крайних типов какой-нибудь популяции ближайшее поколение изменится в направлении отбора. Однако, это верно, только когда присутствуют различные генетические факторы, и даже при этом процесс скоро кончается, как только указанные факторы будут выделены. Ничего действительно нового не достигается, за исключением большего числа особей соответствующих типов, причём захождения за пределы исходной популяции не происходит».
В той же книжке Моргана на странице 106 мы читаем: «Отсюда следует, что естественный отбор не играет созидающей роли эволюции». Таким образом, взгляд генетиков на отбор, как искусственный, так и естественный, резко расходится с учением Дарвина. Созидательной роли в эволюционном процессе за отбором генетики по существу не признают. С их точки зрения, никакие отклонения в индивидуальном развитии организмов не могут играть роли для филогенетических изменений, то есть изменений наследственной природы.
Естественный и искусственный отбор генетики признают только как сито, отсевающее наследственную природу одних организмов от других. Они не хотят понять, что дарвиновский естественный отбор ежесекундно включает в себя наследственность, изменчивость и выживаемость организмов.
Уже одно отрицание творческой роли естественного и умелого искусственного отбора в эволюционном процессе говорит о том, что основные теоретические концепции генетической науки развиваются не в плане эволюционного учения Дарвина. Вокруг этого основного вопроса и идет у нас дискуссия.
Я и мои единомышленники стоим за эволюционное учение Дарвина, за дарвинизм во всех разделах агробиологической науки. Отсюда мы в корне не согласны со взглядами школы Н. И. Вавилова и взглядами многих генетиков на эволюцию, на создание новых форм растений.
В этом вопросе между этими двумя направлениями в науке есть принципиальное различие, примирить которое путём договорённости по отдельным мелким частным вопросам невозможно.
Я не являюсь любителем дискуссии ради дискуссии в теоретических вопросах. Я с темпераментом дискуссирую только в тех случаях, когда вижу, что мне необходимо для выполнения поставленных тех или иных практических заданий преодолеть препятствия, стоящие на дороге моей научной деятельности. По ходу своей работы так я поступал по отдельным разделам физиологии, по отдельным разделам агротехники, поскольку это касалось яровизации как агроприёма. Так я поступил и в разделе селекции.
Мне кажется, что по всем этим разделам дискуссия уже закончилась или почти закончилась.
Какие же работы заставляют меня, вместе с доктором Презентом и рядом других учёных, поставить вопрос о пересмотре исходных генетических позиций? Какие работы привели нас к этой дискуссии? Это — две проблемы. Первая проблема — повышение качества посевного материала растений-самоопылителей путем внутрисортового скрещивания, и вторая проблема — переделка природы растений в нужном нам направлении путём соответствующего их воспитания. Разрешение этих двух проблем и заставило меня вступить в дискуссию по вопросу о наследственности и изменчивости.
Перехожу к первому вопросу. Я начну прямо с того, что многие генетики не признают возможности вырождения сортов-самоопылителей. Это понятно, так как генетики не признают изменчивости генотипа в длительном ряду поколений; это для них является основой также и для отрицания созидательной роли отбора.
Мы же придерживаемся других взглядов. Сорта самоопылителей — чистые линии — при длительной культуре изменяются и отсюда — нередко ухудшаются, вырождаются. Кто хоть немного знает культуру томатов, тот, во-первых, знает, что они принадлежат к самоопылителям, во-вторых, знает, что если без отбора на семена лучших растений культивировать хороший сорт томатов, то уже через 3–5 лет он выродится. На культуре томатов это очень легко подметить потому, что эти растения легко поддаются изменениям, кроме того, к этой культуре люди предъявляют большие требования, и сразу будет подмечено изменение формы плода или изменение времени созревания и т. д.
Дарвин много и обстоятельно занимался вопросом изучения биологической вредности самоопыления и пользы перекрёстного опыления. В результате он пришёл к выводу, что сорта самоопылителей в практике, благодаря самоопылению, не выдерживают длительной культуры. Они ухудшаются, вырождаются, уступают место новым сортам. Не буду зачитывать выдержек из работ Дарвина. Довольно много их привёл д-р И. И. Презент в своей обстоятельной статье, помещённой в журнале «Яровизация» № 3 (1935 г.).
Перейду к объяснению тех причин, благодаря которым может идти изменение, а отсюда нередко и ухудшение сортов-самоопылителей как гибридного, так и негибридного происхождения. При этом мы не будем здесь затрагивать всем хорошо известный вопрос об ухудшении сортов-самоопылителей механическими примесями. Кому не ясно, что на посевах пшеницы не должно быть колосьев ржи, на посевах пшеницы красноколосой не должно быть пшеницы белоколосой. Не об этом идёт речь и не будем об этом говорить. Само собой понятно, что всеми мерами мы должны добиться наибольшей чистосортности не только в элитных посевах, но и на товарных колхозных полях.
Обычно оплодотворённые половые клетки в большей степени обладают всеми возможностями повторения путей развития своих ближайших предков. Наиболее близкими предками являются родители. Поэтому потомство в наибольшей степени, как правило, обладает возможностью повторения пути развития родителей. Обладать же возможностями развития при данных условиях внешней среды — это значит быть приспособленным к тому, чтобы жить и развиваться в данных условиях. Таким образом, мы исходим из дарвиновского положения, что половые клетки в той или иной степени отражают, аккумулируют пройденный путь развития предыдущих поколений, особенно ближайших предков.
У растений-самоопылителей и мужская и женская половые клетки развиваются на одном и том же растении, в одном и том же цветке. Поэтому каждая половая клетка — и мужская и женская — у растений-самоопылителей обычно отражает более тождественный путь пройденного развития, нежели у растений — перекрёстников, где при оплодотворении соединяются мужские и женские половые клетки с разных растений, а следовательно, при оплодотворении в зиготе представлен, отражён путь развития не одного предыдущего растения, а двух.
Я уже указывал, что растение обладает возможностями в той или иной степени отражать, повторять пройденный путь развития своих ближайших предков, а не только непосредственных предков — родителей. Каждому также хорошо известно, что чем более далеки предки, тем всё меньше и меньше их путь развития отражён в данном поколении. Развитие последующих поколений как бы стирает путь развития предыдущих поколений; или, вернее, не стирает, а всё время превращает его в относительно новый.
Исходя из этого, не трудно представить, что растения, например озимой пшеницы Крымки и какой-либо другой озимой пшеницы, полученные из семян после внутрисортового скрещивания, обладают возможностями повторять путь развития как отцовского, так и материнского организма. Поэтому приспособительные возможности к условиям внешней среды у таких растений более многообразны, нежели эти же возможности у отцовского и материнского растения в отдельности.
Чем дольше будут самоопыляться растения, полученные от перекрёстного опыления, тем всё больше и больше будут затухать, сглаживаться различия бывших отцовских и бывших материнских возможностей развития.
Таким образом, у каждой новой генерации, полученной путём самоопыления, всё больше и больше суживается круг приспособительных возможностей развития, полевые же условия никогда не бывают постоянными. Поэтому непластичный организм, суженный в своих приспособительных возможностях, будет хуже развиваться в сравнении с организмами, у которых варьированию полевых условий соответствует вариетабильность возможностей развития. Отсюда получается, что длительного самоопыления сорта обычно не выносят. Культурные сорта самоопылителей при длительной их культуре вырождаются, снижают урожай, и люди заменяют их новыми, более молодыми сортами, дающими больший, лучший урожай.
Напрашивается вопрос, как же в природе тысячелетиями живут виды и расы растений-самоопылителей?
На этот вопрос ответ дал Дарвин. После детальных исследований Дарвин показал (и до сих пор ни одному антидарвинисту не удалось этого опровергнуть), что нет ни одного сорта, ни одной расы культурных и диких растений-самоопылителей, которые не подвергались бы время от времени перекрёстному опылению в том или ином проценте своих представителей.
У диких растений все семена от урожая биологически предназначены для посева, выживает же обычно примерно такое количество растений, сколько их было и в предыдущем поколении. Поэтому если перекрёстное опыление у известной расы дикого самоопылителя происходит только в одном проценте, то и этого вполне достаточно для довольно частого обновления всей данной расы, для освежения её «крови».
У культурных растений получается иная картина. Благополучие культурных растений во многом зависит от человека. У культурного растения, хотя бы у пшеницы, для посева идут не все семена, полученные с урожая, а обычно 5-10 %. Главное же заключается в том, что агротехникой люди создают такие условия, что почти все посеянные растения выживают.
Роль естественного отбора у культурных растений с улучшением агротехники всё больше и больше уменьшается, отсюда — один-два процента естественного перекрёста у культурных растений-самоопылителей не могут обновить, освежить сорт. Растения из семян от естественного внутрисортового перекрёста получают преимущества только для индивидуального развития, в обновлении же сорта эти растения играют значительно меньшую роль, нежели у диких растений. Поэтому для обновления сортов-самоопылителей мы предлагаем искусственно время от времени производить внутрисортовой перекрёст.
Дарвин на основе собранного им громадного материала, а также на основе лично тщательно проведённых опытов пришёл к категорическому выводу: самоопыление биологически вредно, перекрест биологически полезен. Он же доказал, почему в природе создавались и создаются самоопылители. Для выживания, для продолжения потомства растениям бывает нередко полезнее оплодотворяться своей пыльцой, нежели оставаться вовсе неопылёнными, если нет чужой пыльцы, если она не принесена ни ветром, ни насекомыми. Отсюда Дарвин показал биологическую полезность способности самоопыляться.
Необходимо напомнить, что дарвиновское настойчивое утверждение о биологической вредности длительного самоопыления и полезности хотя бы периодического перекрёстного опыления многократно подтверждал и лучший последователь Дарвина — К. А. Тимирязев.
Мне кажется, что каждому человеку, кроме разве людей, догматически исповедующих буржуазную генетику, утверждающих о неизменности генотипа и отдельных генов в десятках тысяч поколений, должно быть ясно, что три миллиона растений наиболее чистосортной пшеницы, размещённых на гектаре посева, не могут быть абсолютно одинаковы по своей природе. Если это сорт Украинка, то все — и вместе взятые и в отдельности — являются Украинкой. А ведь они всё же чем-то отличаются друг от друга в большей или меньшей степени. Но всё это разнообразие укладывается в рамки понятия — сорт Украинка.
Боязнь, высказываемая нашими оппонентами, что скрещивание внутри чистосортного посева создаст разнообразие, уменьшит чистосортность, абсолютно не обоснована. Если посев до внутрисортового скрещивания был чистосортным, то после внутрисортового перекрёста сорт, как правило, будет ещё более выравненным. Скрещивание в большинстве случаев обычно сглаживает, а не создаёт разнообразие. Приведу обычный пример: в любой семье дети между собой более похожи, нежели их отец и мать похожи друг на друга (смех, аплодисменты). Я всегда подчёркивал, что для опыта по внутрисортовому скрещиванию необходимо брать наиболее чистосортные посевы пшеницы, ячменя или других культур-самоопылителей. Проводить же внутрисортовое скрещивание на таких посевах, где среди красноколосой пшеницы имеется и белоколосая пшеница, другими словами — на загрязнённых посевах, никто из нас не советовал.
Я предлагал и предлагаю производить внутрисортовое скрещивание только на чистосортных посевах, а если сорт грязный, то прежде всего его необходимо механическим путём очистить.
Н. И. Вавилов в своём докладе заявил, что проводить внутрисортовое скрещивание, не нужно, бесполезно. В подтверждение этого положения, что сорт самоопылителей не стареет, не вырождается (если исключить загрязнение примесями и случайные скрещивания с другими сортами), Вавилов указал, что есть много примеров, говорящих о столетней и большей долговечности сортов пшеницы, ячменя и других полевых культур-самоопылителей. Мне сразу же вспомнилось прошлогоднее (1935 г.) заявление Н. И. Вавилова в Одессе на выездной сессии зерновой секции Академии, где я в своём докладе подверг критике применявшуюся методику инцухта перекрёстноопыляющихся растений. В ответ на моё утверждение, что этим методом за десятки лет работы никто в мире ещё не создал для производства сортов, Вавилов заявил, что сорта, выведенные методом инцухта, занимают в производстве значительные площади. Конкретно районов и сортов он не назвал по той причине, что тогда под руками этих примеров у него не было.
Прошёл год с лишним, и Н. И. Вавилов вчера в докладе по этому вопросу говорил уже другое. Сортов, выведенных методом инцухта, пока в производстве на больших площадях не оказалось ни по кукурузе, ни по ржи, ни по подсолнечнику. Возможно, что после более детальной проверки Вавилов и по вопросу о невырождаемости сортов-самоопылителей на протяжении столетий придёт к другому выводу.

Рис. 51. Озимая пшеница Крымка.
Слева — посев обычными семенами (контроль), справа — посев семенами от внутрисортового скрещивания (первое поколение). При промораживании в холодильнике растения из семян от внутрисортового скрещивания показали высокую морозостойкость.

Рис. 52. Озимая пшеница Гостианум 0237.
Слева — посев обычными семенами (контроль), справа — посев семенами от внутрисортового скрещивания (первое поколение). При промораживании в холодильнике растения из семян от внутрисортового скрещивания показали высокую морозостойкость.
Пусть даже сортов-самоопылителей, живущих столетиями, будет во много раз больше, нежели то количество, которое Н. И. Вавилов назвал в своём докладе, но эти факты сегодня уже не нужны. Они, может быть, играли бы роль в 1935 г., когда я поднял вопрос о вырождаемости сортов благодаря длительному самоопылению, предлагая противопоставить этому явлению внутрисортовой перекрест. К счастью, этих фактов у генетиков в 1935 г. не было (аплодисменты). При всём огромном сопротивлении и противодействии генетиков, вплоть до высмеивания предлагаемых нами опытов по внутрисортовому скрещиванию, генетики в 1935 г. не могли привести фактического материала, подтверждающего столетнюю неизменяемость природы — генотипа сортов[27].
Если бы этот фактический материал был, то, конечно, в 1936 г. нам было бы несравненно труднее организовать не только массовые проверочные опыты примерно в двух тысячах колхозов различных районов СССР, но и в условиях своего опытного учреждения пришлось бы проводить эти опыты вне утверждённой программы. Ведь с точки зрения генетики этот вопрос просто антинаучен. Несмотря на это, я и теперь берусь утверждать, что долго сорта не живут, не подвергаясь изменениям. И дарвиновское утверждение о биологическом вреде длительного самоопыления остаётся таким же верным, каким оно было и во времена Дарвина. Но сейчас решение этого вопроса перешло уже в новую, более высокую фазу. Сейчас вопрос о пользе и вреде само- и перекрёстно-опыления решается не побочными фактами наблюдений длительности существования сортов-самоопылителей, а путём решающего эксперимента. И произведённое нами, а также сотнями хат-лабораторий внутрисортовое скрещивание является настоящим решающим экспериментом.
Генетикам в настоящее время, мне кажется, надо готовиться не для подыскивания фактов неизменяемости сортов, а подумать о том, как с точки зрения их теории «корпускулярности наследственного вещества» объяснить вырождение сортов от длительного самоопыления, с одной стороны, и, с другой — объяснить благотворное влияние внутрисортового перекрёста.
Ведь в самом деле, товарищи генетики! А вдруг сотни колхозных хат-лабораторий весной 1937 г. на разных сортах, в разных районах уже покажут, что озимая пшеница от внутрисортового перекреста становится более зимостойкой? Вдруг довольно большие опыты Одесского института селекции и генетики по искусственному замораживанию тоже это подтвердят? А что, если, в добавление к этому, и сортоиспытание покажет значительную прибавку урожая озимых пшениц от применения внутрисортового скрещивания? Ведь полевые опыты с яровыми пшеницами у нас в институте это уже показали. Вопрос о внутрисортовом скрещивании у нас уже вышел из рамок теоретического обдумывания; на это было достаточно времени до разворота опытов по внутрисортовому скрещиванию. Теперь же дело находится в таком положении, при котором решать будет практика, эксперимент, а не ссылка на существование сортов столетиями в неизменном виде.
Исходя же из теоретических глубин дарвиновского эволюционного учения, таких сортов не может быть до тех пор, пока люди не овладеют по-настоящему эволюционным процессом и не заставят, если это нужно, природу растений изменяться лишь в нужном для нас направлении.
Генетики говорят, что внутрисортовое скрещивание — дело бесполезное, ибо у чистой линии наследственная природа миллионов растений одинаковая. На всякий случай теперь они уже допускают, что изредка не исключена возможность улучшения природы семян от внутрисортового скрещивания, объясняя это гетерозисом. А если их спросить, а что такое гетерозис на русском языке, они ответят: ну, например, когда растение бывает мощнее или более раннее в сравнении с рядом растущими родителями, так это и называется гетерозисом. В общем большую мощность растений от внутрисортового перекреста генетики объясняют гетерозисом, то есть большей мощностью. (Смех, аплодисменты.)
Пример с объяснением явления мощности «гетерозисом» в генетике далеко не единственный. Так, изменение наследственной основы без скрещивания в генетике называется термином мутация. Когда же начинаешь доискиваться причины изменчивости генотипа, то генетики в один голос заявляют: «Здесь дело ясно, — изменение происходит потому, что организм мутирует». В переводе же на русский язык это означает, что организм наследственно изменяется, потому что происходит наследственное изменение.
Если бы бесконечные генетические термины были переведены на русский язык, так многим из генетиков было бы несравненно легче понять неверные положения их науки, ушедшей в сторону от эволюционного учения Дарвина.
Каковы же результаты проведённого нами эксперимента по внутрисортовому скрещиванию?
В печати я уже несколько раз публиковал результаты проведённого нами летом 1936 г. полевого опыта по внутрисортовому скрещиванию яровой пшеницы. Результаты сортоиспытаний пяти сортов яровой пшеницы, у которых было произведено внутрисортовое скрещивание, для меня и для ряда работающих со мной товарищей вполне убедительны. Они полностью подтвердили наши предположения. Не только первое и второе, но и третье поколение после внутрисортового скрещивания дало повышение урожайности даже у новых сортов гибридного происхождения. Последние дали повышение урожая от полутора до двух центнеров, а старые сорта — от трёх до четырёх центнеров на гектар.
Таковы результаты от внутрисортового скрещивания яровых пшениц.
Одним из препятствий для проведения в колхозах опытов по внутрисортовым скрещиваниям была существующая техника скрещиваний. Ведь высококвалифицированный работник на опытной станции при принятом методе скрещивания мог получить в день не больше 15 г семян (500–600 зёрен): как же предлагать такое дело массам колхозников, да ещё с таким расчётом, чтобы семенами, полученными от скрещивания, засеять хотя бы 0,5 га семенного участка? Но для нас главное было не в этом, главное было в том, — будет или не будет ощутительным хозяйственный эффект от этого мероприятия. Мы были уверены, что если положительный эффект будет, то за разработкой техники скрещивания остановки не будет. В 1936 г. без всякой моральной поддержки со стороны Сельскохозяйственной Академии, при хорошей деловой помощи газеты «Социалистическое земледелие» мы взялись за популяризацию постановки опытов по внутрисортовому скрещиванию в колхозных хатах-лабораториях. Местные работники — областные и районные — сочувственно отнеслись к этому делу. Поэтому нам и удалось в 1936 г. провести внутрисортовые скрещивания примерно в двух тысячах колхозов. В каждом колхозе этой работой занималось по 5–8 человек. Значит, минимум 10 тыс. колхозников прекрасно овладели методом скрещивания.
Почему колхозники так быстро научились скрещивать? Потому, что был изменён, упрощён способ скрещивания. И напрасно проф. Вакар в 12-м номере журнала «Социалистическая реконструкция сельского хозяйства» в своей статье пишет, что способ кастрации, предложенный Долгушиным, негоден. «Я его сам проверил, — пишет Вакар, — у меня получилось только 1–2 процента завязывания семян». Но какое нам дело, проф. Вакар, до того, что у вас этот способ дал плохие результаты? Ведь в двух тысячах колхозов получилось завязывание семян у 80–90 процентов кастрированных цветков, а это более важно. Причём просмотр в колхозах контрольных изолированных колосьев показал, что работа по кастрации в громадном большинстве была проведена хорошо.
Сотни гектаров семенных участков колхозов осенью 1936 г. были уже засеяны для размножения семенами озимых пшениц, полученными от внутрисортового скрещивания. В трёх пунктах проведены сортоиспытания: в Институте селекции, на Одесской областной станции (Выгода) и на Московской областной станции.
В 1936 г. за 4–5 дней работы 5–7 колхозников получили примерно по килограмму семян от скрещивания. Думаю, что на будущий год можно будет получить по 5-10 кг (аплодисменты). Некоторые товарищи говорили: «Ну, ладно, вам удалось получить в колхозе килограмм обновлённых семян. Но что такое килограмм семян для колхоза?». Эти товарищи забывают или просто не знают неограниченных возможностей быстрого размножения семян в теперешних условиях наших колхозов при наличии хат-лабораторий, при наличии небывалой ещё борьбы за качество семян. Нам вместе с 19 колхозами Одесского района в 1936 г. удалось в обычных полевых условиях засушливого лета из 130 кг семян яровой пшеницы к осени получить около 190 ц семян. Из килограмма семян через 2 года можно иметь 50–60 т семян. Никаких трудностей здесь нет, и колхозы это легко выполнят. В сотнях колхозов, в разных местах Советского Союза проведены посевы озимой пшеницы семенами, полученными от внутрисортового скрещивания. В 1937 г. любой человек сможет убедиться в том, что будут уже получены сотни тонн обновлённых семян из посева семенами от внутрисортового скрещивания, проведённого в 1936 г.
Все предварительные результаты, полученные нами от внутрисортового скрещивания озимых пшениц, также говорят о том, что метод внутрисортовых скрещиваний будет одним из способов улучшения качеств природы семян. Поэтому я прошу академиков, научных сотрудников и Академию в целом как можно скорее убедиться самим в полезности этого мероприятия.
Ведь если подтвердится наше предположение о том, что благодаря большей мощности озимых растений, развившихся из семян от внутрисортового скрещивания (а они на деле значительно более мощные), у них будет и большая зимостойкость, то нам необходимо будет добиваться летом 1937 г. проведения этого мероприятия хотя бы в 50–70 тыс. колхозов.
Каждому из генетиков и другим научным работникам теперь уже есть возможность убедиться в эффективности этого мероприятия на сотнях гектаров в колхозах и в трёх точках сортоиспытания. Время есть. Предстоят ещё зима и весна. Если проводимые опыты с озимой пшеницей дадут хороший эффект, то нужно будет провести большую организационную работу. Взять хотя бы такой вопрос, как приобретение ножниц[28]. Нам потребуется для этого дела до 500 тыс. ножниц. То же самое и с подготовкой кадров. Для работы по внутрисортовому скрещиванию необходимо будет подготовить до 500 тыс. колхозников. Дело Академии взять на себя всю эту большую организационную работу.
Перехожу ко второму вопросу моего доклада — переделка природы растений путём воспитания.
Никто не возьмёт на себя смелости сказать, что внешние условия не играют роли в эволюционном процессе растительной формы. В то же время генетики категорически отрицают возможность направленного изменения наследственной основы растений путём соответствующего воспитания их в ряде поколений.
Любую попытку овладеть этим процессом генетики сразу же без разбора зачисляют в разряд ламаркизма. Они забывают, что, исходя из позиции ламаркизма, в работе не может получиться положительный результат. Если же нам путём соответствующего воспитания растений уже удаётся направленно переделывать их природу наследственности, то это говорит уже за то, что мы не ламаркисты и исходим не из ламаркистских позиций.
Ведь, нельзя же учёного только потому, что он признаёт неотъемлемую роль внешних условий в эволюционном процессе растительного и животного мира, зачислять в ламаркисты.
Трудно найти большего врага ламаркизма, чем д-р Презент, и в то же время тов. Презент, как вы знаете, не только поддерживает идею переделки наследственной природы растений путём соответствующего воспитания, но сам является одним из немногих авторов постановки довольно широких опытов в этом направлении.
В общих чертах всем ясно, что внешние условия играют колоссальную роль в бесконечном процессе формирования растительных организмов. Но, насколько мне известно, никому ещё не удалось экспериментально показать и доказать, какие условия, когда, в какие моменты развития растений необходимы для того, чтобы в заданном направлении изменять природу растений последующих поколений.
Уровень знаний нашей советской науки о развитии растений, мне кажется, уже достаточен для того, чтобы взяться за действенное овладение процессом направленного формообразования.
Наилучше, наиболее полно и детально мы знаем биологию развития того этапа в цикле развития растения, который мы именуем стадией яровизации. Поэтому, естественно, и опыты по направленной переделке природы растений, путём соответствующего их воспитания, мы начали с переделки природы этих изученных нами процессов, то есть по изменению наследственной природы растения в отношении стадии яровизации.
Теперь уже хорошо известно, что отличие природы озимых растений от яровых заключается в том, что для одного из периодов индивидуального развития, называемого периодом (стадией) яровизации, озимых растений требуются более низкие температурные условия, нежели для яровых растений. Досконально выяснено, что сорта озимых и яровых растений (пшеницы, ячменя, ржи, овса и др.) представлены переходным рядом по свойству озимости. Одни сорта требуют для прохождения стадии яровизации более низких температур и более длительного периода времени, другие сорта требуют менее низких температур, третьи сорта для прохождения стадии яровизации требуют ещё менее низких температур и могут довольно быстро проходить свою стадию яровизации ежегодно при весеннем посеве. Последние сорта и называются яровыми.
Таким образом, выяснено, что различные сорта, например пшеницы, являются в разной степени озимыми или яровыми.
Различная степень яровости или озимости создаётся и закрепляется естественным отбором в процессе эволюционного формирования этих растений. Вопрос закрепления свойств озимости или яровости для агронауки был уже давно ясен. Озимые при весеннем посеве не могут плодоносить, поэтому они выпадают, не оставляют потомства, — остаются только яровые формы. Наоборот, яровые растения пшеницы и многих других культур при осеннем посеве зимой вымерзают, — остаются только озимые. Неясен был вопрос: каким путём идёт само создание озимости или яровости?
Мы уже довольно хорошо изучили условия, требуемые озимыми растениями для прохождения ими стадии яровизации. На этой основе путём предпосевной яровизации озимые растения любого сорта пшеницы, ржи, вики и других культур в любом районе при весеннем посеве можно заставить плодоносить.

Рис. 53. Озимая пшеница Украинка.
Сверху — посев обычными семенами (контроль), снизу — посев семенами от внутрисортового скрещивания (первое поколение). При промораживании в холодильнике растения из семян от внутрисортового скрещивания оказались более морозостойкими.
На основе этих же знаний мы в 1935 г. приступили к постановке опытов но переделке путём воспитания природы самих требований растений к условиям внешней среды для прохождения стадии яровизации.
Известно, что путём предпосевной яровизации, разработанной нами как агротехнический приём, природа растений, их требования к условиям внешней среды не переделываются. Путём предпосевной яровизации мы создаём, лишь те внешние условия, которые требуются этими растениями для нормального индивидуального их развития. В данном же случае мы поставили вопрос о переделке уже самой природы требований этих растений.
Теоретические предпосылки для постановки этих опытов были следующие. Требования растений к условиям внешней среды для развития, например для прохождения стадии яровизации, у каждого сорта выражены своей амплитудой колебаний. Например, для яровизации озимой пшеницы Кооператорка требуется температура примерно от 0 до 15–20° тепла, причём при 15–20° растения Кооператорки хотя и проходят стадию яровизации, но чрезвычайно медленно. Так, если яровизация сорта Кооператорка при температуре 0–2° требует 40 дней, то при температуре 15–20° она требует 100–150 дней. Для практического, хозяйственного выращивания озимых растений яровизировать при температуре 15–20° невозможно.
Понятно, что прохождение процессов яровизации у растений одногои того же сорта при температуре 0–2° и при температуре 15–20° будет относительно разным не только по быстроте течения процессов, но оно будет относительно разным и качественно. Клетки конуса нарастания растений пшеницы (где и происходят процессы яровизации), развиваясь при температуре 0–2°, и клетки конуса нарастания другого растения пшеницы того же сорта, развиваясь (яровизируясь) при температуре 15–20°, будут сходными в том отношении, что и первые и вторые обладают качеством яровизации. Только это качество (качество яровизации) клеток может дальше видоизменяться, развиваться в направлении движения к образованию колоса и органов плодоношения. Но эти яровизированные клетки — у одного растения при 0–2°, а у другого при температуре 15–20° — будут качественно отличаться друг от друга. Ведь то или иное температурное состояние является необходимейшей составной частью условий прохождения процесса яровизации. Поэтому мы и предположили, что при разных температурных условиях будут получаться яровизированные клетки относительно разные, причём это относительное биологическое различие будет заключаться в том, что следующее поколение растений, полученное из. семян растений, яровизированных при высокой температуре (15–20°), будет легче, быстрее проходить свою стадию яровизации при этой же высокой температуре.
Ход наших рассуждений был такой. Яровизированные клетки получаются в конусе нарастания. Из них развивается вся дальнейшая непрерывная цепь клеток соломины, колоса, мужских и женских половых клеток. Таким образом, в известный момент развития растений яровизированные клетки являются непосредственными исходными клетками для всего дальнейшего построения организма, который в будущем даст зрелые семена. Поэтому, если яровизированные клетки у двух растений одного и того же сорта, благодаря разным температурным условиям в период прохождения яровизации, получились разными, то это различие в той или иной форме будет пронесено всеми последующими клетками и биологически отражено в половых клетках, следовательно, и в новых семенах.
Растения в своих требованиях к условиям внешней среды для прохождения всего цикла развития, как говорит д-р Презент, повторяют, в известной мере отображают, пройденный путь предков. Чем более близки предки, тем в большей степени их пройденный путь развития остаётся биологически запечатлён, аккумулирован в данном потомстве. Поэтому, несмотря на то, что взятая нами для опыта озимая пшеница, для переделки её природы в яровую, веками, из поколения в поколение проходила процесс яровизации при сравнительно низких температурах (намного ниже, нежели яровые пшеницы), мы предположили, что не так уже много потребуется новых поколений, яровизируемых при высоких температурах, чтобы эта пшеница стала наследственно яровой.
Необходимо подчеркнуть, что когда мы говорим — путём выдерживания растений в тот или иной момент их развития, например при прохождении озимыми стадии яровизации, необходимо этим растениям предоставлять высокую температуру для переделки их в яровые формы, то это не значит, что необходимо этим растениям давать как можно более высокую температуру. Если температура будет выше той, при которой данные организмы могут хотя бы медленно проходить стадию яровизации, то эти растения не будут яровизироваться, — они просто будут ожидать более пониженных температурных условий или погибнут. При переделке нормы требований внешних условий для прохождения соответствующих процессов (например, в нашем опыте процесса яровизации) необходимо выдерживать растения примерно на той грани этих требований (обусловленных природой взятого растения), в какую сторону хотят в потомстве этих растений сдвинуть норму требований.
Исходя из вышеизложенных теоретических предпосылок, базируясь на нашем знании развития растительных организмов, мы в Институте генетики и селекции (Одесса) в настоящее время и развернули довольно широкие опыты по переделке, путём соответствующего воспитания, природы растений.

Рис. 54. Кооператорка посева 30 сентября 1936 г.
Слева — первое поколение (контроль), справа — третье поколение.
В настоящее время этими опытами охвачены: озимая пшеница — по переделке её в одних случаях в яровую, в других (и это главное) в более озимую; озимая рожь — по переделке её в яровую; хлопчатник — по переделке его в сторону меньшей теплолюбивости, и некоторые другие растения. Наиболее продолжительными опытами в настоящее время являются у нас опыты по переделке озимой пшеницы сорта Кооператорка в яровую.
Эти опыты начаты в марте 1935 г. За пройденный период мы вырастили три поколения и в сентябре 1936 г. уже высеяли четвёртое поколение этих растений.
Благоприятные результаты опытов с Кооператоркой заставили нас включить в работу и ряд других растений, но из этих растений мы до настоящего времени успели только вырастить первое поколение и высеять второе. Поэтому результатов опытов с этими растениями, за исключением опытов с рожью, ещё нет. Приведём кратко результаты опытов по переделке озимой пшеницы Кооператорка в яровую. Для опытов были взяты два растения озимой пшеницы Кооператорка и два растения Лютесценс 329 Саратовской станции, высеянных 3 марта 1935 г. в теплице в одном вазоне.
Цель посева — не дать этим озимым растениям пройти стадию яровизации и этим заставить их как можно дольше жить, не давая выколашивания. Но вазон с этими растениями, высеянными 3 марта, был оставлен не в тёплой теплице, а в прохладной, где температура с 3 марта и до конца апреля нередко была не выше 10-15°. Только начиная с мая температура была более высокой, а главное, не спускалась ниже 15°. Оба растения сорта Лютесценс 329, как более озимые, нежели Кооператорка, жили до глубокой осени и, не дав выколашивания, погибли. Растения сорта Кооператорка в начале августа имели вид сильно раскустившихся со многими живыми и отмершими листьями. Одиночные побеги у этих растений в начале августа развили соломины.
Примерно в середине августа одно растение Кооператорки из-за вредителей, подгрызших его корни, погибло. Осталось только одно растение, с которого 9 сентября было собрано несколько первых зёрен. Плодоношение у этого растения было чрезвычайно растянутое, вплоть до января 1936 г., когда это растение со многими ещё зелёными колосьями погибло.
9 сентября 1935 г. был произведён посев первых собранных семян с указанного растения Кооператорки. Одновременно в тех же условиях были высеяны для сравнения и контроля обычные семена Кооператорки, взятые из склада. Посев был произведён в теплице, где в ноябре-декабре температура обычно не бывает ниже 15–20°. Опытные растения уже примерно через месяц после посева можно было на глаз отличить от контрольных. На вид они были более яровыми (более приподнятые листья, более тугая листовая обвёртка), нежели контроль. Зимой, ввиду малого количества дневного света, все растения страдали. Выколашивание наступило в конце января. Опытные растения колосились значительно лучше по сравнению с контрольными, причём у многих колосьев настолько были укорочены ости, что их на глаз легко можно было отличить от контрольных. У опытных растений значительно больше было стерильных колосьев, нежели у контрольных. В общем в этом посеве уже ясно было видно, что опытные растения отличаются своим поведением, своей природой от контрольных.
28 марта 1936 г. был произведён новый посев семян, собранных с опытных растений (этот посев мы называем посевом третьего поколения). Одновременно был произведён посев и семян, собранных с бывших, контрольных растений (этот посев мы называем вторым поколением), а также произведён посев обычных семян Кооператорки, взятых со склада. Все эти растения были поставлены в более тёплые условия, нежели первый посев, произведённый 3 марта 1935 г.
В третьем посеве результаты опыта проявились значительно резче, нежели в предыдущих посевах. Прежде всего, контрольные растения не только не дали выколашивания, но большинство из них к осени погибло. Растения второго поколения (из бывшего контрольного посева 9 сентября 1935 г.) чувствовали себя хотя и лучше в сравнении с контрольными растениями, незначительно хуже, нежели растения третьего поколения. Колошение растений третьего поколения в сравнении с колошением указанного второго поколения наступило значительно (дней на 30–50) раньше, а главное, более дружно. Растения второго поколения имели много листьев и только по 1–2 стебля, растения же третьего поколения дали полностью выколашивание из всех побегов кущения.
В этом же посеве у нас было второе поколение из остатков семян первого опытного растения, проходившего яровизацию не зимой, а летом 1935 г., то есть в более тёплых условиях. Поведение этих растений было ближе к поведению третьего поколения из семян растений, проходивших яровизацию зимой в теплице. Наибольшая же разница в поведении растений была между третьим поколением и контролем.
Все растения третьего поколения, начиная с августа, дали полное колошение, а контрольные растения только к концу сентября дали два выколосившихся стебля (из всех растений).
Одновременно со значительными изменениями в стадии яровизации в сторону яровости, у второго поколения и, особенно, у третьего поколения произошли изменения во многих признаках колоса, чешуи, длины остей и др. В третьем поколении появились узколистные формы.
В этих опытах легко было наблюдать, что чем сильнее было подвергнуто растение переделке в стадии яровизации, тем сильнее расстраивалась согласованность дальнейшего развития организма.
Мы приходим к выводу, что необходимо переделывать растения более постепенно, давать менее жёсткие условия, тогда организм меньше будет расстроен, времени же на переделку потребуется не больше, так как быстрее можно будет выращивать поколения.

Рис. 55. Кооператорка посева 19 сентября 1936 г.
В первых 6 вазонах слева — растения первого поколения (контроль), в остальных вазонах — растения третьего поколения.
В сентябре 1936 г. было высеяно четвёртое поколение, а также был проведён посев из запасных семян третьего и второго поколений и контрольными (обычными) семенами, взятыми со склада. В этом посеве ещё более резки различия между разными поколениями растений сорта Кооператорка. Старшие поколения на 50-60-й день от посева уже дали трубки (развитые соломины). Контрольные же растения ведут себя, как типичные озимые. Ещё более лёгкой переделке в сторону яровости поддаётся озимая рожь. Нами была взята озимая рожь Таращанская и высеяна весной сеялкой в поле без предпосевной яровизации. Растения этого посева долго кустились, в середине лета часть из них выколосилась и дала урожай семян. Часть этих семян 29августа 1936 г. была высеяна одновременно с посевом обычных семян (для контроля). Уже через месяц после посева легко было наблюдать различие в поведении этих растений. В начале ноября опытные растения дали значительно более дружное развитие соломы, нежели контрольные, — из контрольных выколосились буквально единичные растения.
Приводимые некоторыми товарищами возражения, что рожь как перекрёстник неконстантна и что здесь поэтому произошла не переделка природы растений, а просто отбор более яровых форм, мы отводим. В самом деле, если рожь имеет в своей «крови» свойство яровости, а яровость — безупречный доминант, то как же она многие годы зимовала в наших районах? Яровые растения ржи в наших районах не могут зимовать. Кроме того, по растениям второго поколения (нашего опыта) видно, что они ещё не полностью яровые.
Переделка ржи идёт более быстро и более легко в сравнении с пшеницей, на наш взгляд, только потому, что рожь всегда является гибридом, как растение перекрёстноопыляющееся. Гибриды же всегда более легко подвергаются изменениям, у них круг приспособительных возможностей более широкий. Поэтому в дальнейшем для переделки природы растений мы берём уже и у самоопылителей, например у хлопчатника, не константные формы, а гибриды (первое поколение).
На первый взгляд может показаться, что описанные мною опыты никакого практического значения не имеют. На самом же деле это далеко не так. Ведь если мы можем превращать озимые растения в яровые путём воспитания их в период прохождения стадии яровизации при относительно высоких температурах, так это значит, что мы можем изменить природу растений и в противоположном направлении. Воспитывая растения в момент прохождения стадии яровизации при более низкой температуре, ниже, чем они яровизировались в природных условиях, мы будем их делать всё более и более озимыми. Иначе говоря, мы будем делать растения всё более и более зимостойкими, ибо длительность стадии яровизации является хотя и не единственным, но одним из основных факторов зимостойкости.
Подтверждения того, что озимые пшеницы путём соответствующего их воспитания можно из поколения в поколение превращать в более озимые, можно найти и в некоторых фактах из жизни. Все мы хорошо знаем, что наиболее зимостойкие сорта в нашем Советском Союзе — это сорта озимой пшеницы Саратовской станции. Известно, что одной из многих заслуг Саратовской селекционной станции является выведение самых зимостойких в мире сортов озимой пшеницы. Наименее зимостойкими сортами в европейской части Советского Союза являются сорта, выведенные Одесской селекционной станцией (Кооператорка, Земка). Если взять сорта Харьковской станции, то они значительно более зимостойки, чем одесские, и приближаются в этом отношении к саратовским. Одесские же сорта более зимостойки, нежели сорта озимой пшеницы южной селекции, например Кировабадской станции (Азербайджан).
Отдавая должное заслугам селекционеров всех перечисленных станций, нельзя, конечно, обойти того факта, что чем суровее район, в котором расположена селекционная станция, тем её сорта, как правило, являются более морозостойкими.
Если мы вспомним, что саратовские сорта более озимые, то есть имеют более длинную стадию яровизации, в сравнении с харьковскими, и харьковские имеют более длинную стадию яровизации, в сравнении с одесскими, то нетрудно придти к выводу, что в создании природы растений с той или иной длительностью стадии яровизации играют роль какие-то внешние условия в конкретных районах.
Задача заключается в том, чтобы правильно расшифровать эти условия. Мы должны уметь их создавать в любом пункте селекции для того, чтобы выводить сорта с необходимой нам зимостойкостью. Этими условиями является температура поля при прохождении стадии яровизации растениями озими в том или ином районе.
Температура при яровизации озимой пшеницы на полях районов Саратова и Харькова из поколения в поколение, как правило, более низкая, нежели в районе Одессы. На наш взгляд, это и является одним из главнейших природных районных условий, создающих ту или иную степень озимости, а отсюда — определяющих в немалой степени и зимостойкость озимых сортов.
При температуре ниже 1–2°мороза (это доказано довольно большим экспериментальным материалом) яровизация озимых пшениц не идёт. Следовательно, во всех районах яровизация проходит в тот период, когда ещё не установились постоянные морозы, то есть при температуре выше нуля. Для нас сейчас ясно, что чем ближе к нулю температура в полевых условиях в период прохождения пшеницей, из поколения в поколение, стадии яровизации, тем более озимые сорта создаются в данном районе.
В то же время из приведённых ниже в таблице цифр мы видим, что ни в одном из перечисленных нами пунктов посевы пшеницы не проходят яровизацию всё время при температуре 0° или хотя бы близкой к этой температуре.
Среднемесячные температуры за пять лет (1905–1909 гг.)
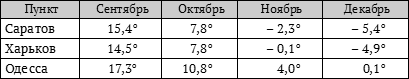
Из этой таблицы следует, что яровизация озимой пшеницы, высеянной в сентябре, проходит в полевых условиях Одессы при более высокой температуре, чем в районах Саратова и Харькова. Но даже и в условиях Саратова и Харькова яровизация озими идёт в сентябре и октябре при температурах много выше 0°. Проведение же яровизации озимых пшениц при температуре 0° должно сдвинуть в потомстве природу растений в сторону большей озимости.
На этой основе мы сейчас уже развёртываем опытную работу по повышению зимостойкости наших пшениц. Мы проводим яровизацию посевного материала в искусственной обстановке при температуре 0° и высеваем этот материал в поле.
Нами и рядом других исследователей (Ф. М. Куперман, М. Т. Тимофеева, Е. П. Мельник) было установлено, что после прохождения стадии яровизации растения несравненно хуже закаливаются против морозов. Поэтому, высеяв осенью яровизированный при 0° посевной материал, мы обрекаем на вымерзание все наименее зимостойкие формы. Останутся только наиболее стойкие формы — те, которые имеют или более длинную стадию яровизации, вследствие чего в холодильнике не успели её закончить, или, если и закончили стадию яровизации, то по другим свойственным им качествам могут переносить зимовку.
В нашем посеве проводится, таким образом, жёсткий отбор более зимостойких форм. В то же время идёт и переделка природы растений, увеличение у их потомства озимости, так как родители проходят стадию яровизации при температуре значительно более низкой, чем в природных условиях.
Эту работу мы предполагаем повторять из года в год. Пшеница из года в год будет делаться всё более и более озимой, то есть более зимостойкой. Для этого часть урожая, полученного от таких посевов, будет идти в размножение на семена, а другая часть будет подвергаться воздействию с целью дальнейшего изменения стадии яровизации.
Этим же самым путём мы приступили и к переделке природы «теплолюбивых» растений. В самом деле, если можно «холодолюбивые» растения, например озимую пшеницу, которая не мирится с теплом в период яровизации, заставить ослаблять свои требования к пониженным температурам в этот период, то и такое растение, как хлопчатник, можно путём соответствующего воспитания заставить постепенно ослабить требования к повышенным температурам в первые дни своей жизни.
То же самое можно сказать и о переделке растений, требующих в известные моменты своего развития короткого или длинного дня и т. д.
Главное здесь заключается в том, чтобы, поняв по-мичурински развитие растений, поняв роль и место внешних условий в эволюции растительных форм, сознательно делать то, что в природе делалось и делается случайно.
В природе путём изменчивости и естественного отбора могли создаваться и создаются прекраснейшие формы животных и растений. Человек, овладев этим путём, во-первых, сможет творить такие прекрасные формы в неизмеримо более короткие сроки, а во-вторых, сможет создать и такие формы, каких не было и какие не могли появиться в природе и за миллионы лет.
Генетики по-настоящему не вдумываются в действенно осваиваемые нами глубины дарвиновского эволюционного учения, развитого и в отдельных своих частях конкретизированного лучшими биологами мира, в первых рядах которых стоят К. А. Тимирязев и И. В. Мичурин.
Всё вышеизложенное мною говорит о том, что на базе развития дарвиновского эволюционного учения мы экспериментально овладеваем и в некоторой степени уже овладели делом направленного изменения наследственной основы растительной формы.
Изложенное мною в докладе, конечно, не укладывается в очерченные генетической наукой рамки. И генетики пытаются спрятаться от нашей критики путём приписывания нам ламаркизма, отрицания материальности наследственной основы и других «смертных грехов». Но это говорит лишь за то, что критики плохо разбираются в ламаркизме, называя ламаркизмом, как и полагается метафизикам-автономистам, всякое признание действенной роли внешних условий. Не лучше они разбираются и в материализме. Они в своей механистической ограниченности полагают, что, если мы отрицаем единственность и исключительность роли морфологии хромосом, этим самым мы якобы отрицаем материализм,
Высказанные мною положения действительно противоречат генетической концепции. В этой части я с предъявляемым нам обвинением вполне согласен, но высказанное мною не только не противоречит фактическому материалу, полученному и получаемому генетиками, а, наоборот, объясняет его с позиции развития. Поэтому многое из экспериментального материала, добытого генетиками и бывшего в их руках мёртвым капиталом, будучи освещено с позиции развития, становится причиной действенного знания.
Основное, из чего исходит генетическая наука в своих построениях и с чем мы не можем согласиться, заключается в том, что в организме — в хромосомах клеток — есть отличное от тела организма «вещество наследственности». Это особое вещество наследственности (генотип) отделено от тела организма и не входит в понятие самого тела уже потому, что оно не подвержено видоизменениям, превращениям.
Вещество сомы — тела организма — развивается: отсюда оно не только может изменяться, но оно обязательно видоизменяется, превращается. «Вещество наследственности», которое является частью хромосом, на взгляд генетиков, как правило, за редкими исключениями не подвержено изменениям, — следовательно, оно не подвержено и развитию, иначе что же это за развитие без видоизменений, превращений? «Вещество наследственности» (гены) размножается самовоспроизведением, и интереснее всего то, что, размножаясь, увеличиваясь, начиная от зиготы и до взрослого организма, в миллионы раз, оно не изменяется, не превращается. Таким образом, основное отличие выдуманного генетиками «вещества наследственности» от вещества тела организма — это неизменяемость в длительном ряде поколений первого вещества и изменяемость второго, из которого в организме строятся различные органы и признаки. Согласно генетической концепции, «вещество наследственности» бессмертно; из поколения в поколение тянется нить этого неизменного вещества; вещество же, из которого состоит тело организма, всё время путём ассимиляции и диссимиляции изменяется и с концом индивидуальной жизни умирает. Изложенное мною в докладе, конечно, в корне противоречит и «закону» гомологических или параллельных рядов изменчивости Н. И. Вавилова. Этот «закон» в своей основе зиждется на генетической теории комбинаторики неизменных в длительном ряду поколений корпускул «вещества наследственности». Я не чувствую в себе достаточной силы, знаний и уменья, чтобы по-настоящему разбить этот «закон», не отвечающий действительности эволюционного процесса. Но в своих работах я всё время наталкиваюсь на неприемлемость этого «закона»; сама работа говорит, что нельзя мириться с этим «законом», если ты берёшься за действенное, направленное овладение эволюцией растительных форм.
Согласно «закону» параллельной изменчивости новые формы получаются не путём развития старых форм, а путём пересортировки, перекомбинации из начала существующих корпускул наследственности. Получается точный параллелизм изменчивости видов, родов и даже семейств. Причём указание на параллелизм изменчивости Н. И. Вавилов выставляет как основу действенности этого «закона» в руках экспериментатора. На взгляд Н. И. Вавилова получается, что стоит только знать разнообразие форм, допустим, пшеницы, ячменя, как на этой основе можно предвидеть всё разнообразие форм ржи, овса и других культур. Разнообразие последних культур должно быть точно таким же, как и разнообразие пшеницы. Если же в природе какой-либо формы не окажется в готовом виде, так, согласно «закону» гомологических рядов, эту форму можно создать. Если пшеница могла дать, допустим, безлигульную форму, значит она есть или может быть и у ржи. В природе встречаются круглые плоды яблони — следовательно, должны быть или могут быть деревья с круглыми плодами груши, вишни, винограда и т. д. В общем стоит только подметить что-либо у одного вида, как, согласно «закону» гомологических рядов, такие же признаки должны или могут быть и у других видов.
На первый взгляд, особенно, если не знать или не желать знать дарвинизм, действенность «закона» гомологических рядов подкупающая. Но это может случиться только с теми людьми, которые не знают эволюционного учения Дарвина, не знают, как создавались и создаются в природе и в человеческой практике новые формы растений и животных.
Несколько слов следует сказать о кажущейся практической действенности «закона» гомологических рядов. Если в природе есть круглоплодная яблоня, следовательно, могут быть круглые плоды и у груши, вишни, винограда и т. д. Ну, а если в природе нет дикой бессемянной яблони, вишни, сливы, так по «закону» гомологических рядов нельзя иметь в культуре и бессемянной груши? А ведь таковую селекционеры создали. Махровых цветов у видов капусты нет, а у левкоя такие цветы садоводы создали, невзирая на то, что у диких крестоцветных махровости не наблюдается.
Стоит только сопоставить изменчивость диких форм с культурными, как сразу иллюзорность обязательности параллелизма изменчивости исчезнет.
Согласно дарвиновскому эволюционному учению, расы, виды и роды растений всё время развиваются, дают начало новым расами видам. Эволюционное учение прекрасно объясняет общность и постепенность происхождения всего настоящего многообразия живых форм и указывает нам пути улучшения старых и создания новых, нужных нам сортов растений.
Закон же гомологических рядов говорит о том, что виды и роды в своей эволюции не могут расходиться. Дивергенция невозможна благодаря точному параллелизму изменчивости. Если же дивергенции в настоящее время нет, то, следовательно, согласно «закону» гомологических рядов, её не было и в прошлом. Все теперешние виды, выходит, были и в прошлом, они только были менее разнообразны своими формами, зато каждая форма была более богата своими потенциями, набором генов. Отсюда, из «закона» гомологических рядов вытекает геногеография, центры и фонды генов и т. д.
Вот чем объясняются наши расхождения с генетиками, наши несогласия со школой Н. И. Вавилова, исходящей из «закона» гомологических рядов, противоречащего основам дарвиновского эволюционного учения.
Наши оппоненты говорят, что Лысенко отрицает генетику, то есть науку о наследственности и изменчивости. Это неправда. Мы боремся за науку о наследственности и изменчивости, а не отбрасываем её.
Мы боремся против многих неверных, вымышленных генетических положений. Мы боремся за развитие генетики на основах и в плане дарвиновского эволюционного учения. Генетику, как один из важнейших разделов агробиологической науки, нам необходимо как можно скорее и полнее освоить, переработать на наш советский лад, а не просто принимать многие антидарвиновские установки основных генетических положений.
Блестящие работы цитологии, многое уже давшие в смысле морфологического описания клетки и, особенно, ядра, никто из нас не отрицает, мы целиком поддерживаем развитие этой работы. Так же глупо было бы отрицать полезнейшую работу систематиков, описание морфологии пшеницы или других культур, в том числе и диких форм. Всё это нужные разделы науки, дающие знания. Но мы отрицаем то, что генетики, вместе с цитологами, увидят под микроскопом ген. Пользуясь микроскопом, можно и нужно будет видеть всё больше и больше деталей в клетке, ядре, в отдельных хромосомах, но это будут кусочки клетки, ядра или хромосомы, а не то, что генетика разумеет под геном.
Наследственная основа не является каким-то, особым от тела, саморазмножающимся веществом. Наследственной основой является клетка, которая развивается, превращается в организм. В этой клетке равнозначимы разные органеллы. Но нет ни одного кусочка, не подверженного развитию эволюции.
На этом я и закончу. (Бурные, продолжительные аплодисменты.).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК