Лофотрохозои: в шкафу только скелеты
Кольчатые черви, долгое время считавшиеся несомненными предками членистоногих, поскольку они членистые, передвигаются с помощью двуветвистых конечностей (параподий) и имеют обособленный головной отдел, в кембрийских слоях тоже встречаются — в виде отпечатков покровов, конечно. Но в тысячи раз реже, чем го-ловохоботные. Представлены они исключительно остатками подвижных многощетинковых червей. Причем ни один из кембрийских кольчецов по совокупности признаков не вписывается в какую-либо современную группу: у них отсутствуют антенны, опорные щетинки, спинные и брюшные усики, а щупики устроены очень просто. Сколекодонты — ископаемые челюсти многощетинковых червей — появляются только в ордовикских отложениях, а достоверные сидячие трубчатые кольчецы — в мезозойскую эру.
Ископаемая летопись древних кольчецов служит хорошей иллюстрацией идеи зоолога Вильфрида Вестхайде из Университета Оснабрюкка о происхождении щетинок как покровных защитных элементов у подвижных форм, ползавших по поверхности осадка: именно такими — «ощетинившимися» — мы видим первых кольчецов, например канадию (Canadia); затем, по мере развития червевидно-го способа передвижения, в покрове появляются поперечные, лишенные щетинок прорехи, и все тело становится как бы сегментированным. В работах по молекулярной филогенетике кольчатых червей коллектива Торстена Струка из того же университета говорится об отчетливом разделении этой группы на две основные ветви — бродячие и сидячие. Общего предка всех кольчецов Струк и его соавторы тоже реконструируют как подвижную сегментированную форму с развитыми, закономерно расположенными вдоль всего тела параподиями. Примечательно, что среди ордовикских кольчецов встречались махайри-дии {Machaeridia) — сегментированные формы с обызве-ствленными элитрами — чешуевидными спинными усиками параподий, под которыми, как выяснили палеонтолог Джейкоб Винтер и его коллеги из Йельского университета, скрывались щетинки. Элитры махайридий структурно близки к раковинам моллюсков полиплакофор, или хитонов, и разнообразных древних «многостворок».
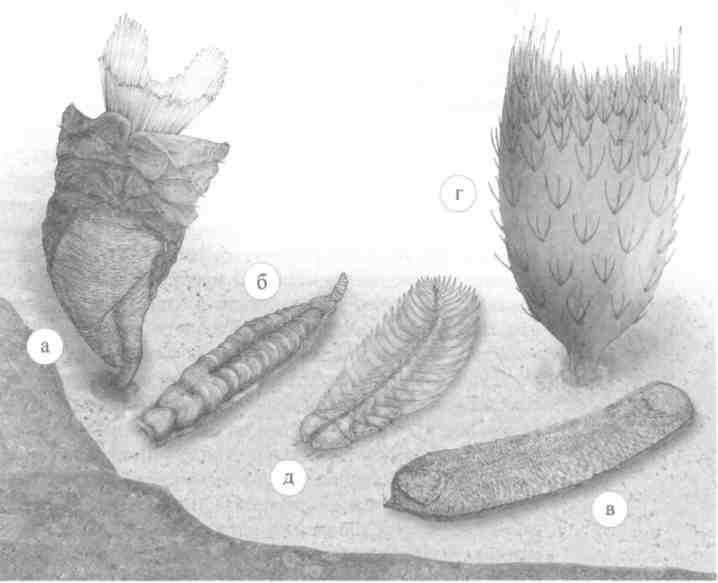
Кембрийские (а — г) и ордовикский (д) Lophotrochozoa, 540–480 миллионов лет: (а, б) томмотииды: а) Paterimitra, б) Camenella, {в, г) целосклеритофоры: в) Halkieria, г) Allonnia (ханцеллорииды), д) кольчатый червь Plumulites (махаиридии). Художник Всеволод Абрамов
Кембрийские многостворчатые формы — халькиерии-ды (Halkieriidae) — представляют наибольший интерес с точки зрения происхождения моллюсков. Сложный покровный скелет этих существ, или склеритом, развитый на спинной и боковой сторонах, состоит из многочисленных известковых и/или органических склеритов и спикул различной формы и одной-двух раковин на переднем и заднем концах тела, напоминающих раковины брахиопод или моноплакофор. Палеонтологи Саймон Конвей Моррис из Кембриджского и Джон Пил из Упсальского университетов, сравнивая крупные торцевые раковины с таковыми брахиопод, а прочие склериты — с элитрами и щетинками кольчатых червей, рассматривают халькиериид как исходную группу для Lophotrochozoa в целом. Напротив, зоологи Амели Шелтема из Океанографического института в Вудс-Холе и Дмитрий Люмбергович Иванов из Зоологического музея МГУ, опираясь на данные по строению склеритов — наличие сложной системы каналов, связывающих внутреннюю полость склерита с его поверхностью, а также ротового аппарата, напоминающего радулу, — считают их несомненными моллюсками, предшественниками хитонов и беспанцирных моллюсков — аплакофор. Интересно, что даже в самых жестких элементах склери-тома, если полностью растворить их минеральную составляющую, просматриваются капилляры — такие же, как в щетинках одновозрастных кольчатых червей, например канадий.
Еще больше усложняют (но, может быть, и проясняют) ситуацию ханцеллорииды (Chancelloriida), склериты которых обычно собраны в розетки с одним полым центральным шипом и тремя периферическими. Микроструктура этих склеритов и общий план строения неотличимы от таковой халькиериид, но образовывали они совершенно иную конструкцию — полусферический или узкоконический склеритом с открытой вверх полостью. Из-за этого ханцеллориид долгое время считали губками: ведь нижним концом такой организм закреплялся на грунте.
Если самые древние моллюски, за исключением ханцеллориид, и кольчатые черви были представлены исключительно подвижными формами, иная картина наблюдается среди древнейших щупальцевых, или лофофорат. Кембрийские щупальцевые различаются числом створок: от двух, как у брахиопод до тридцати и более, как у пате-римитры и эксцентротеки (Paterimitra, Eccentrotheca), принадлежащих к вымершим томмотиидам (Tommotiida).
Томмотииды были открыты Владимиром Владимировичем Миссаржевским из Геолотического института РАН на реке Алдане и получили название по самому крупному тамошнему поселку — Томмот. У эксцентротеки — одной из древнейших томмотиид — фосфатные пористые раковинки были двух типов: неправильной колпачковидной и уплощенной форм. Эти склериты, как установили палеонтолог Кристиан Сковстед и его коллеги из Упсальского университета, располагаясь по спирали, образовывали узкоконический, расширяющийся вверх, наружный скелет 4-миллиметровой высоты, открытый сверху и снизу. Склеритом патеримитры в общих чертах был устроен, как у эксцентротеки, но со склеритами трех типов, два из которых напоминали раковины брахиопод. Сходство с последними усугубляется тождеством микроструктуры раковин и наличием пор с отпечатками щетинок (благодаря фосфатизации иногда сохраняются и сами щетинки). Эти особенности позволяют рассматривать томмотиид как исходную для брахиопод группу. Особенно интересны как переходные формы двустворчатая томмотиида микрина {Micrina) и таннуолина (Tannuolina) с небольшим числом склеритов: у микрины Уве Бальтасар из Упсальского университета обнаружил двустворчатую эмбриональную раковину с карманами, где формировались щетинки, как у брахиопод.
Интересно, что две группы томмотиид, представленные единственным родом каждая — каменелла (Camenella) и суннагиния (Sunnaginid), обладают парными зеркально-симметричными склеритами, которые, вероятно, располагались на поверхности двусторонне-симметричных животных. Значит, среди первых лофофорат были не только сидячие фильтраторы, но и ползавшие по грунту в поисках пищи существа. Вот только дали они начало каким-либо нынешним щупальцевым или исчезли без следа подобно ханцеллориидам, единственным сидячим древним моллю-скоподобным формам?
Если бы не кембрийские ископаемые, представить, как выглядели общие предки брахиопод, мшанок, моллюсков и кольчатых червей, было бы исключительно сложно. Молекулярная биология на этот вопрос ответить не в состоянии, а современные лофотрохозои практически не сохранили общих для этой группы признаков. К важнейшим из них, как считают эмбриолог Стефани Глайн и ее коллеги из Калифорнийского университета (Беркли), следует отнести спиральный тип дробления зародыша и плавающую в водной толще личинку — трохофору с предротовым венчиком ресничек, ресничным султаном и теменной пластинкой, которая при превращении личинки во взрослый организм погружается под покровы и образует головной мозг. Спиральный тип дробления установлен у кольчатых червей, сипункулид, эхиурид, мизостомид, камптозой, фо-ронид, немертин и моллюсков, а похожие личинки есть у кольчатых червей, сипункулид, эхиурид, мизостомид, камптозой, форонид, мшанок, циклиофор и моллюсков; правда, у немертин и брахиопод трохофора очень упрощенная, а у головоногих моллюсков и поясковых червей она считается утраченной. Общей чертой Lophotrochozoa, распознаваемой в ископаемой летописи, следует считать скелетные микроструктуры (моллюски, мшанки, брахио-поды, кольчатые черви, возможные ископаемые форони-ды) и хитиновые ?- или ?-хитин) щетинки, выделяемые в мешковидных впячиваниях кутикулы крупными покровными клетками с микроворсинками, следы которых остаются в виде капилляров.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК