Общение и групповое поведение
Общение и групповое поведение
У высших беспозвоночных уже в полном объеме представлена категория поведения, которая, как было показано, только в зачаточной форме обнаруживается у некоторых представителей низших многоклеточных беспозвоночных. Речь идет об общении, которое у высших беспозвоночных проявляется иногда и в очень сложных формах подлинного группового поведения (в отличие от колониальной жизни, свойственной многим низшим беспозвоночным). Напомним, что общение, в точном значении этого термина, появляется только тогда, когда контакты между животными начинают осуществляться посредством специальных сигнальных действий.
У головоногих моллюсков и членистоногих, особенно у насекомых, мы находим четкие системы коммуникации с передачей информации по разным сенсорным каналам. Это относится прежде всего к пчелам и муравьям, что, конечно, связано со сложной, высокодифференцированной структурой их сообществ и разделением функций между членами последних (распределение пищедобывательнои, защитной и воспроизводительной функций между отдельными особями).
У муравьев разные формы химической чувствительности играют, очевидно, большую роль, чем другие виды рецепции, и общение между особями осуществляется прежде всего по этому каналу: личинки муравьев выделяют специфические вещества, побуждающие взрослых рабочих особей кормить их, по запаху обитатели одного муравейника узнают «своих» и «чужих». Даже о том, жив или мертв муравей, его сородичи узнают по специфическим выделениям.
Согласование поведения обитателей муравейника происходит путем подачи и приема химических сигналов. Достаточно указать на химические средства муравьев (а также пчел и ос), выделение которых служит сигналом тревоги. Эти вещества выделяются муравьями в случае опасности и быстро распространяются по воздуху. Их восприятие вызывает у других муравьев («солдат») защитную реакцию (готовность к нападению на врага), хотя источник опасности им еще не известен. Поскольку, однако, радиус действия такого сигнала, подаваемого одним муравьем (например, в случае вторжения в муравейник какого-либо насекомого), невелик, то он способен указать и место, откуда угрожает опасность. Чем больше эта опасность, тем больше муравьев выделяют химические сигналы тревоги и тем больше, следовательно, особей призывается на помощь.
По другим выделениям, оставляемым на субстрате в виде следа, муравьи узнают, куда, в какую сторону ушел их сородич. К тому же интенсивность такого пахучего следа указывает на количество пищи, обнаруженной в том месте, куда ведет след, тем более что, направляясь к муравейнику, муравьи наносили по пути пахучие метки лишь в тех случаях, когда они находили корм. При этом тем больше оставляется меток, чем больше обнаружено корма. Поскольку эти метки довольно быстро испаряются, вновь идущие по этому следу муравьи будут воспринимать лишь свежие метки, а таких будет все меньше по мере приближения кормового объекта. Таким образом, муравьи, в конце концов узнают о том, что пищевые ресурсы в данном месте исчерпаны, и перестанут к нему направляться.
У некоторых пчел также существует химический способ передачи информации о нахождении пищи. Это относится, прежде всего, к пчелам — из подсемейства Meliponinae, у которых нет «языка танца». Эти широко распространенные в тропиках пчелы не имеют жала, но яростно кусаются. У мелких форм (самая маленькая имеет длину всего 1,5 миллиметра) нашедшая пищу сборщица только приводит других пчел в возбужденное состояние, беспокойно бегая по сотам, толкая их и время от времени подбегая к летку. Вылетающие же пчелы ориентируются только по запаху цветов. У других видов возбуждение пчел первоначально достигается подобным же образом, но дополнительно сборщица, принесшая взяток, приводит соты толчками в вибрирующее состояние. После этого она возвращается к обнаруженным цветам, а на обратном пути оставляет на траве, ветках или камнях через каждые два-три метра пахучие метки, по которым затем приводит группами других пчел к источнику пищи. Это достигается тем, что пчела-разведчица летает взад и вперед между двумя соседними метками, пока другие пчелы не присоединяются к ней. Следовательно, здесь мы имеем дело с комплексной передачей информации по оптическому, тактильно-вибрационному и, возможно, акустическому каналам, но при этом ведущую роль играют оставляемые сборщицей запахи.
У нашей медоносной пчелы также существует химический способ передачи информации, но только как вспомогательный, когда она имеет дело с кормом, лишенным собственного заметного запаха. В этих случаях место нахождения кормового объекта метится секретом из так называемой железы Насонова.
Однако основным и наиболее совершенным способом передачи информации о кормовых объектах являются «танцы». Найдя источник пищи (нектара) и вернувшись в улей, пчела раздает другим пчелам-сборщицам пробы нектара и приступает к «танцу», который состоит из пробежек по сотам. Рисунок танца зависит от местонахождения обнаруженного источника пищи: если он находится рядом с ульем (на расстоянии 2–5 метров от него), то производится «танец-толчок»: пчела беспорядочно бежит по сотам, время от времени виляя брюшком; если же корм обнаружен на расстоянии до 100 метров от улья, то выполняется один из описанных К. Фришем танцев — «круговой», который состоит из пробежек по кругу попеременно по часовой стрелке и против нее (рис. 38). Если же нектар обнаружен на большем расстоянии, то выполняется «виляющий» танец, также описанный и изученный Фришем. Это пробежки по прямой, сопровождающиеся виляющими движениями брюшка с возвращением к исходной точке то слева, то справа (рис. 38, а). Интенсивность виляющих движений указывает на расстояние находки: чем ближе находится кормовой объект, тем интенсивнее выполняется танец (рис. 38, б).
Кроме того, как тоже показал Фриш, во второй форме танца угол между линией пробежки и вертикалью (на вертикально расположенных сотах) соответствует углу между линией полета пчелы от улья к кормовому объекту и положением солнца. Таким образом, горизонтальное направление истинного полета переносится на вертикальную плоскость, ориентация по положению солнца подменяется ориентацией по земному притяжению. При этом, если следует лететь навстречу солнцу, пчела танцует снизу вверх, если же от солнца, то сверху вниз (рис. 38, в). Таким образом, медоносная пчела способна к транспозиции с фотоменотаксисов на геоменотаксисы: фиксированный угол к видимому положению солнца переносится в темном улье к силе тяжести.
Эта способность медоносной пчелы к транспозиции, коренящаяся в широко распространенной среди насекомых способности к переносу менотаксисных движений на вертикальную плоскость (рис. 39), обеспечивает, как мы видим, исключительную полноту и точность передачи информации. Такая транспозиция является частным случаем трансформации, лежащей в основе всякого общения между животными: свойства действительных предметов и ситуации трансформируются на языке животных с помощью видоспецифического «кода» в оптические, акустические и прочие условные сигналы, информирующие других особей об этих свойствах. Несмотря на всю сложность и точность, система коммуникации медоносной пчелы в принципе не отличается от любой другой системы общения у животных. Переводятся ли истинные пространственные параметры в сигналы, воспринимаемые с помощью зрения, осязания и чувства силы тяжести, как это имеет место у медоносной пчелы, или же в сигналы, воспринимаемые хеморецепцией (муравьи), — информация в любом случае передается в трансформированном, условном виде. Здесь как нельзя нагляднее выступают характер и функциональное значение психики у животных.
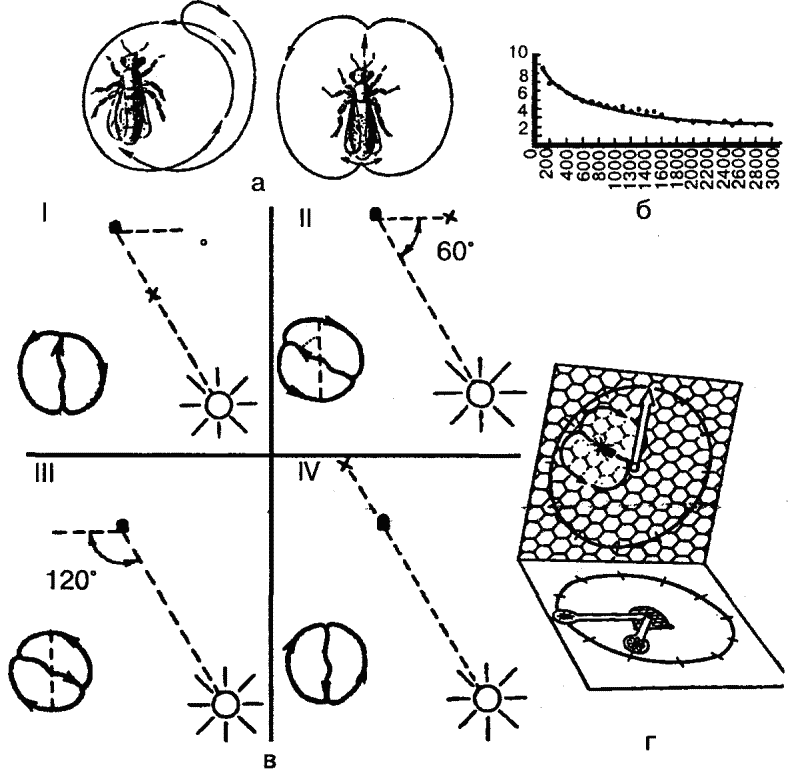
Рис. 38. «Танцы пчел». Обозначения: а — «круговой танец» (слева) и «виляющий» (справа); б — интенсивность выполнения «виляющих танцев» как показатель расстояния до местонахождения источника пищи (на оси ординат отмечено число «танцев» за 15 секунд, на оси абсцисс — расстояние до пищи в метрах); в — «виляющий танец» как показатель направления полета к источнику пищи: I — навстречу солнцу, II — под углом 60? налево от положения солнца, III — направо от положения солнца, IV — от солнца. Точка — местонахождение улья, крестик — местонахождение пищи. Изображение «танцев» следует представить себе на вертикальной плоскости (по Фришу); г — схема, иллюстрирующая транспозицию угла полета по отношению к положению солнца (горизонтальная плоскость) на вертикальную плоскость сот, на которых пчела выполняет «виляющий танец» (по Кэстнеру)
Вершиной развития инстинктивных компонентов общения является ритуализация поведения, которая особенно четко выступает в воспроизводительном поведении насекомых, в частности в «ухаживании» самцов за самками. Вот типичный брачный «ритуал» самца бабочки-бархатницы, описанный Тинбергеном: полетев вслед за самкой, самец садится рядом с ней, потом становится перед ней, начинает «дрожать» крыльями (переднезадние движения сомкнутых крыльев), затем повторно раскрывает и закрывает их, что сопровождается постукиванием усиками по крыльям самки. После этого самец обхватывает самку крыльями и зажимает между ними ее усики, которое при этом прикасаются к пахучему участку на крыльях самца. Только после этого, если самка не отвергает «ухаживания» самца, начинается спаривание.
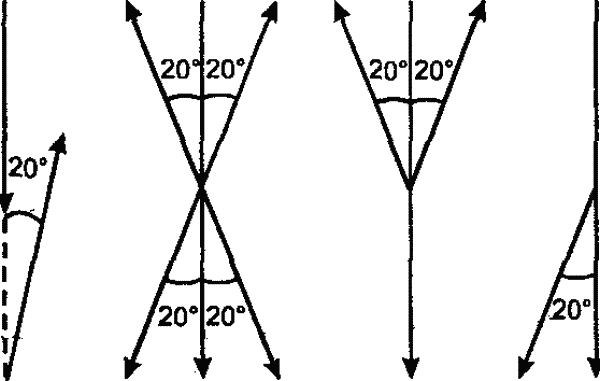
Рис. 39. Транспозиция оптической ориентации в другую сенсорную сферу у разных насекомых. Обозначения: а — менотаксисное движение по горизонтальной плоскости под углом 20° к источнику света. Вертикальная стрелка — направление светового луча; б—г — продолжение движения по вертикальной плоскости у муравья (б), божьей коровки (в), майского жука (г). Вертикальная стрелка — направление силы тяжести
В этом примере обращает на себя внимание строгая фиксированность последовательности сигнальных поз и телодвижений, что и характерно для ритуализованного поведения. Как мы видим, при ритуализации двигательные акты всегда носят условный характер: выполняемые животным движения несут лишь сигнальную, но не непосредственно «механическую» функцию. Именно на этой основе зародился язык поз, телодвижений и звуков у животных во всем своем удивительном многообразии. При этом в условности сигналов находит непосредственное воплощение психический компонент общения.
Наиболее сложные системы общения присущи, конечно, насекомым, живущим большими семьями. Это относится опять-таки в первую очередь к медоносной пчеле и муравьям. Дифференцированные и многообразные взаимоотношения между членами пчелиной семьи, опирающиеся на сложные формы коммуникации, являются уникальными в животном мире. По поводу этого сообщества высказывались самые различные мнения начиная от антропоморфичного толкования ее как копии человеческого общества или государства (сохранилось неудачное, вводящее в заблуждение название «общественные насекомые») до представления о некоей сумме независимо действующих «рефлекторных автоматов». Вместе с тем оказалось (исследования К. П. Трубецкой), что пчелиная семья представляет собой объединение особей, поведение которых носит самостоятельный, но не независимый характер. Самостоятельность поведения отдельных пчел проявляется прежде всего в резко выраженных индивидуальных различиях: предпочтение отдельных видов ульевых работ, различия в сроках и характере перехода от ульевой деятельности к летной и др. Большую сложность в количественном и качественном аспектах достигают взаимоотношения между обитателями улья, в результате чего функциональная настроенность одних пчел может оказать влияние на других. Рабочие особи с их повышенным уровнем нервной деятельности часто сообщают возбуждение другим, менее активным рабочим пчелам.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ИНТЕЛЛЕКТУАЛЬНЫЕ СПОСОБНОСТИ И ГРУППОВОЕ ПОВЕДЕНИЕ
ИНТЕЛЛЕКТУАЛЬНЫЕ СПОСОБНОСТИ И ГРУППОВОЕ ПОВЕДЕНИЕ Муравей обладает не только ощущениями, но и рассудком, разумом, памятью. Цицерон. О природе богов Я отметил, сколь счастливо, кажется, живут вместе члены одного сообщества. Высшая гармония царит повсюду.
Общение
Общение Исследования биологической коммуникации (общения) насекомых и птиц продвинулись достаточно далеко, в то время как для большинства видов млекопитающих они находятся в начальной стадии. Этология охотничье-промысловых животных изучает значение для них различных
4. СОЗНАНИЕ, ОБЩЕНИЕ И РЕЧЬ
4. СОЗНАНИЕ, ОБЩЕНИЕ И РЕЧЬ Концепция коммуникативной природы сознания была выдвинута впервые П. В. Симоновым [1981]. Позднее сходные мысли были высказаны другими авторами [Hesslow, 1994; Frith, 1995]. По определению П.В. Симонова, сознание представляет собой знание, которое в
Поведение
Поведение Умственное и социальное развитие щенков продолжает прогрессировать и к возрасту в три — четыре недели они начинают играть в простые игры, например, «салочки», бегая друг за другом. И уже в этом раннем возрасте обычно определяется лидер в стае. В возрасте пяти
Поведение
Поведение В начальные сутки опытов у многих животных поведение было значительно изменено. Собаки почти все время находились в активных позах, много прыгали, вертелись, у них часто регистрировались продолжительные двигательные реакции большой интенсивности. Одна из них
Поведение
Поведение Позные состояния. В переходный период животные 68% времени по данным наблюдения находились в активных позах. Расслабленные положения тела регистрировались больше глубокой ночью. Иногда днем наблюдалось возникновение сидячих пассивных поз, что объяснялось
Поведение
Поведение Позные состояния, движения и голосовые реакции при удлинении опытов были чрезвычайно разнообразны и в большей степени, чем любые другие функции, носили на себе отпечаток индивидуальных особенностей отдельных животных. В первые 20 суток поведение изменялось в
13. Общение, общение и еще раз общение
13. Общение, общение и еще раз общение Внезапно Homo sapiens — человек разумный — стал самым многочисленным из всех крупных животных. Десять тысяч лет назад в мире насчитывалось около 10 млн. людей. Они были изобретательны, находчивы и активно общались между собой, однако как
Инстинктивное поведение и общение
Инстинктивное поведение и общение Все животные периодически вступают во внутривидовые контакты друг с другом. Прежде всего это относится к сфере размножения, где часто наблюдается более или менее тесный контакт между половыми партнерами. Кроме того, представители
Научение и общение. Подражание
Научение и общение. Подражание Существенную роль в формировании поведения высших животных играют явления подражания, которые в основном, хотя и не все, относятся к сфере научения. Мы здесь не будем рассматривать формы подражания, которые относятся к инстинктивному
Общение
Общение У высших позвоночных особой сложности достигают и процессы общения. Как и у других животных, средства коммуникации включают у них элементы различной модальности — ольфакторные, тактильные.Ольфакторная сигнализация, т. е. передача информации другим особям
Групповое поведение обезьян и зарождение общественных отношений
Групповое поведение обезьян и зарождение общественных отношений Общественные отношения зародились в недрах первых форм трудовой деятельности. Труд с самого начала был коллективным, общественным. Это вытекало уже из того, что люди с момента своего появления на земле
Общение – это не только разговоры
Общение – это не только разговоры Но как предсказание того, что человек будет делать дальше, позволяет решать проблему общения? Как бы ни были хороши мои предположения и мои предсказания, каким бы похожим я ни сделался на вас, я никак не смогу напрямую сравнить смысл того,
Поведение типа А – «борьба или бегство», поведение типа Б – «затаивание»
Поведение типа А – «борьба или бегство», поведение типа Б – «затаивание» Два этих термина сначала были предложены для описания поведения животных, у которых значительная часть стрессорных событий связана с угрозой нападения хищников и неблагоприятными изменениями
Феромоны и общение человека с животными
Феромоны и общение человека с животными Владельцам домашних животных хорошо известна проблема запахов. Хотя феромоны действуют в малых концентрациях, которые не вызывают ощущения запаха, в природе животные должны обеспечить его распространение на большие расстояния.
Общение животных и язык человека: качественные различия или эволюционная преемственность?
Общение животных и язык человека: качественные различия или эволюционная преемственность? На симпозиуме в битком набитой аудитории председательствовала Беатриса Гарднер. В своем кратком вступительном слове она признала тему симпозиума дискуссионной. Говоря об