Сенсорные способности, таксисы
Сенсорные способности, таксисы
Специфические условия, в которых живут насекомые, далеко идущее приспособление двигательной активности при обилии и разнообразии качественно весьма различных агентов среды, управляющих их поведением, — все это обусловило появление у данных животных многих, и притом своеобразно устроенных, органов чувств.
Большую роль в жизни насекомых играет зрение. В разном количестве у разных видов имеются простые одиночные глаза. Но главный орган зрения — своеобразный сложный глаз, получивший название фасеточного, так как его поверхность состоит из отдельных, четко ограниченных участков — фасеток. Каждой фасетке соответствует самостоятельный глазок — омматидий, изолированный от своих соседей пигментной прокладкой и функционирующий независимо от них. Общий внешний покров глаза образует над омматидиями утолщения, обозначаемые как роговица или роговой хрусталик и играющие роль диоптрического аппарата глаза. Каждый омматидий содержит 7–8 зрительных клеток, аксоны которых оканчиваются в оптических ганглиях мозга.
Благодаря такому устройству сложных глаз в мозг насекомого передаются «мозаичные» изображения воспринимаемых предметных компонентов среды. Конечно, это не означает, что насекомое действительно видит окружающий мир в виде мозаики. В мозгу поступающие из отдельных омматидиев зрительные раздражения синтезируются в целостную картину, как это имеет место и у позвоночных. Все же нам очень трудно представить себе, как на самом деле видит насекомое.
Еще в прошлом веке делались попытки установить путем фотографирования через фасеточные глаза четкость и дальность зрения насекомых и раков. Позже, в частности в 60-х годах нашего века, такие опыты повторялись, но едва ли возможно таким образом решить проблему, ибо, как известно, видит не глаз, а мозг. Насекомые, очевидно, весьма близоруки: ясное видение, например, бабочки-крапивницы возможно лишь на расстоянии приблизительно 2–3 сантиметров. От объектов, расположенных дальше (уже начиная с 5–7 сантиметров), насекомые получают лишь расплывчатые изображения, но, очевидно, этого достаточно для их общей ориентировки в пространстве.
На расстоянии пчела способна увидеть лишь объекты в сто раз большей величины, чем это необходимо для зрения человека. Отдельный цветок она увидит лишь тогда, когда она приблизится к нему вплотную. Все эти особенности зрения насекомых вполне соответствуют тому микроландшафту, в котором живут эти животные.
Острота зрения (разрешающая способность, т. е. способность видеть две сближенные точки) зависит у насекомых при прочих равных условиях, в частности, от количества омматидиев, которое, однако, сильно колеблется даже в пределах отрядов. Так, в отряде жуков это количество колеблется от трех-четырех десятков до нескольких десятков тысяч. У пчелы каждый глаз состоит приблизительно из четырех тысяч фасеток. Чем мельче фасетки, тем слабее в них освещение, но тем детальнее зрение. Особенно большое значение это имеет для предметного зрения. Кроме того, при более выпуклом глазе оси омматидиев сильнее расходятся, и в результате расширяется поле зрения. Одновременно суживается точка, видимая одной фасеткой, чем также увеличивается острота зрения.
Наилучшим образом насекомые видят подвижные объекты или неподвижные предметы во время собственного движения. Пролетающая над лугом пчела, например, видит покачивающиеся на ветру цветы или же изображение неподвижного цветка скользит по ее фасеточному глазу. В любом случае это подвижное (расплывчатое) изображение заставляет ее спуститься вниз и обследовать растение с минимального, т. е. оптимального для зрения пчелы, расстояния.
Что касается цветового зрения, то видимый ими спектр шире, чем у человека, так как его граница проходит в ультрафиолетовом секторе. Спорный вопрос о способности насекомых к восприятию красного цвета в настоящее время, очевидно, решен в положительном смысле (за исключением медоносной пчелы и некоторых других насекомых). Советский энтомолог Г. А. Мазохин-Поршняков считает, что вообще цветовое зрение у взрослых насекомых представляет универсальное явление. По его данным, цвета различаются даже видами, активная жизнь которых протекает ночью.
До сих пор мы касались только реакции насекомых на свет, который играет в их жизни очень большую роль. Не меньшее, если не большее значение имеют для них запахи. В естественном поведении насекомых стимулы различной модальности всегда выступают в комплексе. Так, уже упомянутые осы рода Philanthus охотятся за пчелами, руководствуясь зрением с расстояния 30 сантиметров, но опознают их по запаху лишь с расстояния 15–5 сантиметров. Во время зрительного поиска оса совершенно не реагирует на запах пчелы.
Аналогично обстоит дело и у других членистоногих. Водяные пауки рода Pirata ориентируются прежде всего по вибрации поверхности воды, в непосредственной же близости от жертвы — зрением (рис. 36). Другие пауки узнают о попадании в их сеть насекомого, воспринимая вибрации паутины. Но обследование жертвы производится сразу тремя способами: паук прикасается к ней передними конечностями, снабженными хеморецепторами, а также так называемыми щупиками (органы осязания), и, кроме того, паук на вкус пробует жертву укусом.
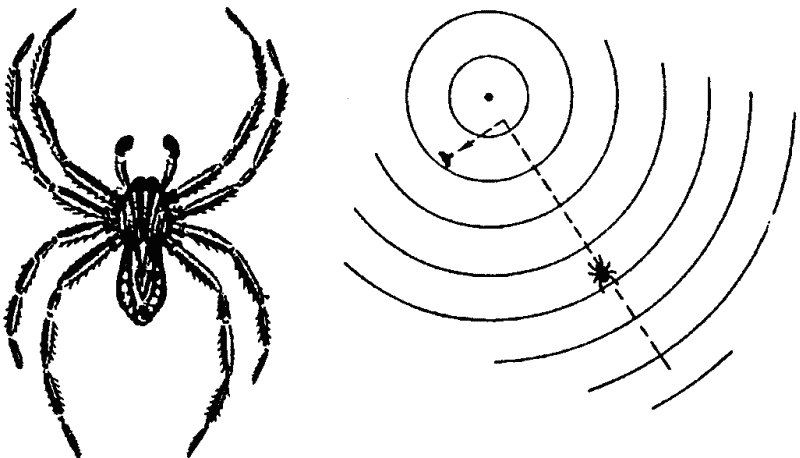
Рис. 36. Водяной наук Pirata бежит по поверхности воды к источнику вибрации (центральная точка). Только непосредственно около неподвижной мухи вид последней приобретает преобладающее влияние и паук сворачивает к ней (опыт Берестынской-Вильчек)
Для ознакомления с таксисами у насекомых рассмотрим в качестве примера фототаксисы. Эти таксисы (положительные или отрицательные) встречаются у насекомых прежде всего в виде тропо-, тело- и менотаксисов, хотя не утратили своего значения и примитивные таксисы. Так, например, положительные и отрицательные клинотаксисы с четко выраженными маятникообразными поисковыми движениями играют ведущую роль в оптической ориентации у многих личинок мух. Тропо- и телотаксисы связаны у них друг с другом многими переходными формами. У взрослых насекомых чаще всего встречаются, однако, четко выраженные телотаксисы. Как правило, тропотаксисное движение очень скоро переходит у них в телотаксисное. У комаров и вшей преобладает тропотаксисная ориентация движения.
Менотаксисы, относящиеся к более сложным формам ориентации, встречаются у высших насекомых (муравьев, пчел, бабочек, жуков). Немецкий исследователь поведения насекомых М. Линдауер выделяет здесь следующие четыре случая: 1) угол фиксации источника света случаен (муравьи, жук-навозник); 2) угол фиксации является врожденным (некоторые бабочки); 3) угол фиксации сообщается особями одного вида друг другу («танцы» пчел, информирующие об угле ориентации по положению солнца); 4) угол фиксации выучивается в индивидуальном порядке. Последний случай можно уже отнести к высшей категории таксисов — мнемотаксисам (выученная ориентация), которые широко распространены среди позвоночных, но также встречаются у высших насекомых (пчел, ос, муравьев) (рис. 37).

Рис. 37. Мнемотаксис у роющей осы Philanthus. После отлета осы кольцо из шишек вокруг норки было перемещено в сторому. После возвращения оса ищет норку внутри кольца и находит ее лишь после повторных облетов (опыт Тинбергена)
Необходимо, однако, подчеркнуть, что и в тех случаях, когда ориентация осуществляется на основе других фототаксисов, поведение насекомых одновременно определяется рядом дополнительных факторов. Например, доказано, что у многих насекомых характер их фототаксисного поведения сильно видоизменяется в зависимости от температуры воздуха, времени дня, физиологического состояния и т. д. Хорошо известно, что муха не реагирует на свет, пока спокойно летает по комнате. Но стоит начать ее преследовать, как у нее сейчас же «просыпается» положительный фототелотаксис: она быстро направляется к источнику света, к окну.
Аналогичным образом насекомые реагируют на разнообразные механические, температурные, химические и оптические раздражения, на силу тяжести, и соответственно у них встречаются во многих различных формах тигмо-, рео-, анемо-, гидро-, гео-, термо-, хемо-, фото- и другие таксисы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
§ 2. Тропизмы и таксисы простейших
§ 2. Тропизмы и таксисы простейших Относительно небольшие размеры простейших дают возможность непосредственно использовать рецепторный аппарат мембраны для быстрого изменения поведения. Размер большинства простейших обычно не превышает нескольких миллиметров (рис.
9.2. Сенсорные системы
9.2. Сенсорные системы Сенсорные системы возникают на очень ранних стадиях эволюции животных для непосредственного восприятия сигналов внешнего мира. В ходе этой эволюции постепенно совершенствуется воспринимающая структура, формируются аналитические центры, из
12. Навигационные способности птиц
12. Навигационные способности птиц Открытие способности птиц ориентироваться по солнцу изумило ученых, но то, что во время ночных пролетов птицы ориентируются по звездам, буквально потрясло их. Это было доказано через несколько лет после открытия Крамера молодыми
Интеллектуальные способности
Интеллектуальные способности При исследовании поведения человека мы прежде всего исходим из того, что принято называть интеллектуальностью. Измерение интеллектуальности, как правило, выполняется посредством самых различных тестов. И хотя подобного рода тесты не могут
Неиспользованные способности
Неиспользованные способности Несмотря на серьезные недостатки, изучение коэффициента интеллектуальности все же позволяет сделать предварительное заключение. Нас интересуют два вопроса. Во-первых, есть ли связь между коэффициентами интеллектуальности и генетическими
1.2. Сенсорные пороги
1.2. Сенсорные пороги АБСОЛЮТНАЯ ЧУВСТВИТЕЛЬНОСТЬ СЕНСОРНОЙ СИСТЕМЫ Абсолютная чувствительность сенсорной системы основана на её свойстве обнаруживать слабые, короткие или маленькие по размеру раздражители. Абсолютную чувствительность измеряют порогом той или иной
От чувствительности к способности ощущать?
От чувствительности к способности ощущать? Давайте же, наконец, начнем наше путешествие. Мать-природа (или, как мы ее называем сегодня, процесс эволюции путем естественного отбора), вовсе не обладая даром предвидения, постепенно произвела на свет способных предвидеть
Инстинктивные движения и таксисы
Инстинктивные движения и таксисы Общая характеристика инстинктивных движений уже давалась выше. Также говорилось о том, что они являются «хранителями» самого ценного, жизненно необходимого, что накоплено видом в результате естественного отбора, и что именно этим
Органы чувств и сенсорные способности
Органы чувств и сенсорные способности Большой интерес для познания психической деятельности низших многоклеточных беспозвоночных представляет устройство и функционирование их органов чувств, представленных также весьма различными образованиями в соответствии с
Таксисы
Таксисы Как и на более низких уровнях эволюционного развития, пространственная ориентация совершается на высшем уровне элементарной сенсорной психики, преимущественно на основе примитивных таксисов. Но с усложнением жизнедеятельности организмов возрастают и
Сенсорные особенности
Сенсорные особенности В полном соответствии с высокоразвитыми эффекторными системами высших позвоночных находится и высокий уровень их сенсорных способностей. Наибольшее значение имеют органы слуха и равновесия (внутреннее ухо, начиная с земноводных — дополнительно
СЕНСОРНЫЕ СПОСОБНОСТИ ДИКИХ ЛЮДЕЙ
СЕНСОРНЫЕ СПОСОБНОСТИ ДИКИХ ЛЮДЕЙ Кроме телесной и физиологической перестройки организма у диких людей под давлением изменившихся условий существования начинают развиваться новые психосенсорные адаптации. Как показали наблюдения, снежный человек ощущает
3. Имеющиеся и утерянные способности
3. Имеющиеся и утерянные способности Непосредственный обмен информациейТо, что личность и комендант при жизни тела свободны и что массу информации комендант получает посредством личности, имеет очень важные последствия для тела — если личность убедит коменданта, что
9. Музыкальные способности
9. Музыкальные способности Часто высказывалось предположение о том, что у африканцев хорошие музыкальные способности и особенно выражено чувство ритма. Кажется, это было впервые сказано в XIV в. арабским автором Ибн Бутланом, который писал, что если бы африканцу
6. Музыкальные способности
6. Музыкальные способности Простые музыкальные способности, такие как определение изменения высоты тона и запоминание мелодий, коррелируют с интеллектом и могут считаться компонентом интеллекта (Carroll, 1993). Ввиду этого интересно выяснить, являются ли музыкальные