Пластичность поведения простейших
Пластичность поведения простейших
Как мы видим, и в моторной и в сенсорной сфере поведение достигает у ряда видов простейших известной сложности. Достаточно указать на фобическую реакцию (реакцию испуга) туфельки в вышеописанном примере клинотаксиса: наткнувшись на твердое препятствие (или попав в иную неблагоприятную зону), туфелька резко останавливается и принимает «оборонительное положение», т. е. «съеживается», готовясь пустить в ход ядовитые стрекательные капсулы. Одновременно меняются движения ресничек, происходит тактильное и химическое обследование объекта и т. д. У эвглены фобическая реакция выражается в том, что она, остановившись, начинает производить передним концом тела круговые движения, после чего уплывает в другом направлении (рис. 29).
Ясно, что такая интеграция моторно-сенсорной активности возможна лишь с помощью специальных функциональных структур, аналогичных нервной системе многоклеточных животных. Однако о морфологии этих аналогов еще очень мало известно, и только относительно инфузории удалось с определенной достоверностью доказать существование специальной сетевидной системы проводящих путей, располагающейся в эктоплазме. Очевидно, проведение импульсов осуществляется у простейших и системой градиентов в самой цитоплазме.

Рис. 29. Фобическая реакция эвглены при внезапном изменении освещения (по Джен-нингсу)
Способность к индивидуальному изменению описанных генетически фиксированных форм поведения путем научения выражена у простейших слабо. Все же способность простейших к научению, хотя бы в элементарных формах, сейчас можно считать доказанной. Если, например, держать парамеции некоторое время в треугольном или квадратном (в сечении) сосуде, то они сохраняют привычный путь передвижения вдоль стенок сосуда и после их перемещения в сосуд округлой формы (рис. 30). Аналогичным образом инфузория, плававшая длительное время (около двух часов) в сосуде, имевшем форму треугольника, придерживается этой траектории затем и в квадратном сосуде большей площади (опыты немецкого ученого Ф. Брамштедта). В опытах советской исследовательницы Н. А.Т ушмаловой инфузории были подвергнуты постоянно действующему ритмичному раздражителю — вибрации. В результате животные постепенно переставали реагировать на него обычным образом (сокращением). Исследовательница усматривает в этом пример элементарных следовых реакций, представляющих собой простую форму кратковременной памяти, формирующейся на этом уровне филогенеза за счет чисто молекулярных взаимодействий.
Подобные опыты ставились и другими экспериментаторами, применявшими различные формы воздействия. Высказывались сомнения, действительно ли в таких случаях можно говорить о каких-то формах научения, ибо сильные воздействия могли оказать хотя бы временное вредное влияние на подопытных животных. К тому же вообще в опытах с простейшими очень трудно учесть все возможные побочные факторы (особенно химические изменения среды).
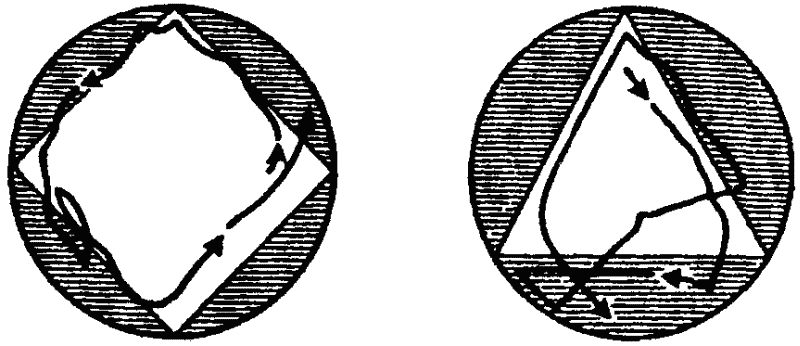
Рис. 30. Привыкание у парамеции (по Брамштедту). Пояснения см. в тексте
Все же, очевидно, мы имеем здесь дело с элементарной формой научения — привыканием. Привыкание (к измененным внешним условиям), как мы уже знаем, играет немаловажную роль и в поведении высших животных, но имеет качественно иной характер уже хотя бы потому, что строится у них не только на ощущениях, но и на восприятиях. При элементарной же сенсорной психике животное может привыкать лишь к воздействию отдельных раздражителей (или их совокупностей), являющихся воплощением отдельных свойств или качеств компонентов окружающей среды. Именно это и имело место в приведенных примерах, когда наблюдалась модификация видотипичного поведения: врожденные реакции на определенные раздражители постепенно элиминируются, если при многократном повторении не наступит биологически значимый эффект. Так, в опытах Тушмаловой фобическая реакция у инфузорий со временем сводилась на нет при повторной встрече с «безобидным» раздражителем.
Привыкание необходимо отличать от утомления, выражающегося в явлениях «истощения» животного. В опытах Тушмаловой это выражалось в неспособности инфузории к дальнейшему реагированию, если она в течение 2–3 часов подвергалась очень сильным раздражениям. Утомление связано с перерасходом энергетических ресурсов, привыкание же представляет собой активную приспособительную реакцию, значение которой состоит в экономии этих ресурсов, в предотвращении траты энергии на бесполезные для животного движения.
Как форма научения привыкание характеризует низший уровень элементарной сенсорной психики, хотя и не теряет своего значения на всех ступенях развития психики, как это было показано при ознакомлении с онтогенезом поведения животных (см. ч. II). Однако у высших представителей простейших, возможно, уже существуют и зачатки ассоциативного научения, вообще характерного для более высоких уровней психического развития.
Об ассоциативном научении можно говорить в тех случаях, когда устанавливается временная связь между биологически значимым и «нейтральным», точнее, биологически маловалентным раздражителем. Именно такую связь Брамштедту удалось выработать у туфельки, которая, как уже отмечалось, не реагирует заметным образом на изменение освещения, но весьма чувствительна к температурным изменениям. Если в эксперименте затенить одну половину капли воды, в которой плывут парамеции, и при этом подогреть освещенную часть капли, то вскоре инфузории соберутся в холодной темной части, но останутся там (в течение 15 минут) и после того, как температура в обеих частях капли будет уравновешена. Правда, эти опыты Брамштедта подвергались серьезной критике, так как при подогреве меняется и химизм воды (меняется растворимость содержащихся в ней газов), что не может не влиять на поведение инфузории (опыты У. Грабовского).
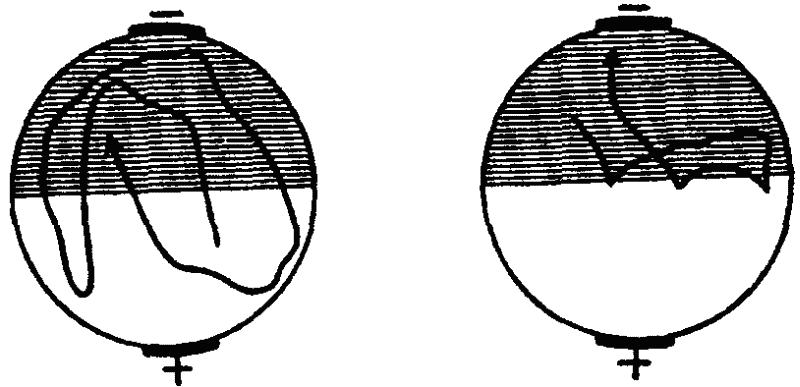
Рис. 31. Образование у парамеции временной связи между электрическим раздражением и светом (по Зесту). Пояснения см. в тексте
Вместе с тем аналогичный результат может быть получен и путем наказания животных при заплыве в освещенную зону ударами электрического тока (рис. 31). Эффект избегания сохранялся в этом случае в течение 20 минут (опыты Г. Зеста). Однако обратный результат, т. е. избегание темноты и заплыв в освещенную зону, Зесту получить не удалось. Это обстоятельство, однако, является результатом методической ошибки: инфузориям свойствен гальванотаксис, выражающийся в их устремлении к катоду. В упомянутых же опытах Зеста катод всегда находился на темной половине и гальванотаксис оказывался сильнее электрического раздражения, почему инфузории и направлялись только в темноту.
В более тщательно поставленных опытах польскому ученому С. Вавржинчику удалось преодолеть эти методические недостатки и с успехом научить инфузории избегать темный участок стеклянной трубки, где их раздражали электрическим током. Постепенно туфельки все чаще оставались в освещенной зоне и поворачивали на границе с затемненной зоной даже при перемещении этой границы вдоль трубки еще до получения электрического удара. Наконец, они оставались в освещенной зоне в четыре раза дольше, чем в темной, даже тогда, когда раздражения током совсем прекратились. Эта реакция сохранялась даже в течение 50 минут, что является достаточно большим сроком для простейших. Четкие положительные результаты этот исследователь получил и в других экспериментах.
Правда, Я. Дембовский, повторивший эти опыты с некоторыми изменениями, пришел к выводу, что, хотя и не исключена возможность выработки у простейших обусловленных реакций, эти опыты с туфельками все-таки не решают до конца сути вопроса, ибо существуют несравненно более сильные факторы, особенно химические, которые маскируют реакции на свет. Ввиду отсутствия вполне убедительных экспериментальных данных Дембовский оставил вопрос о наличии у простейших обусловленных реакций открытым.
Вместе с тем и у других (кроме туфельки) инфузорий, в частности у стентора и сувойки, удалось после 140–160 сочетаний выработать сократительные реакции в ответ на световое раздражение.
На инфузориях проводились и другие опыты с применением разных методических приемов. В частности, использовались капилляры, иногда с загнутым концом, в которые сажались парамеции, после чего измерялось время, необходимое им для высвобождения. В новейших исследованиях, проведенных таким способом, получались вполне положительные результаты: при каждом повторении опыта время выхода парамеции из трубочки заметно уменьшалось, что и признается доказательством их способности к ассоциативному научению.
Однако американские ученые Ф. Б. Эпплуайт и Ф. Т. Гарднер, недавно повторившие эти опыты, считают такой вывод необоснованным. Эти ученые засасывали последовательно в один и тот же капилляр разные, еще ни разу не побывавшие в опыте туфельки и установили, что время выхода этих животных из капилляра также постепенно уменьшается. Но этого не случалось, если капилляр до употребления тщательно промывался и кипятился в деионизированной воде. Использование обработанных таким образом капилляров для повторного проведения опыта с одной и той же туфелькой не давало практически никакого эффекта научения. Отсюда экспериментаторы вывели заключение, что уменьшение времени выхода из капилляра является у парамеции не результатом научения, а реакцией на загрязнение внутренней поверхности трубки продуктами обмена веществ, которое увеличивается от опыта к опыту и от которого животные пытаются все скорее уйти.
С другой стороны, в аналогичных исследованиях других ученых учитывались подобные возможные изменения среды, и было установлено, что время выхода инфузорий из трубки сокращается лишь в первых опытах, потом же оно остается постоянным (среда в трубке должна, однако, продолжать изменяться).
Таким образом, вопрос о наличии у инфузорий (а тем более у простейших вообще) ассоциативного научения еще нельзя считать решенным. Все же, очевидно, такое научение существует у них в зачаточном виде.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
О водорослях и простейших
О водорослях и простейших В мире микроорганизмов мы можем наблюдать ряд признаков, характерных и для высших организмов. Мы уже знаем, что некоторые грибы по размерам относятся к микроорганизмам, а другие видны невооруженным глазом (например, шляпочные лесные грибы). Тем
§ 2. Тропизмы и таксисы простейших
§ 2. Тропизмы и таксисы простейших Относительно небольшие размеры простейших дают возможность непосредственно использовать рецепторный аппарат мембраны для быстрого изменения поведения. Размер большинства простейших обычно не превышает нескольких миллиметров (рис.
Развитие поведения
Развитие поведения Каково же значение этих исследований для человека? Прежде всего генетически фиксированные формы поведения у человека играют подчиненную роль. Строим ли мы дом, общаемся ли со своими друзьями — мы действуем теми способами, которым, сознательно или нет,
Избыточность и эволюционная пластичность
Избыточность и эволюционная пластичность Как мы уже отмечали в главе 4, отбор на стабильность, т. е. фактически на постоянство и неизменность, парадоксальным образом повышает эволюционную пластичность (приспособляемость) организмов, т. е. склонность к изменениям.
Основы поведения
Основы поведения Когда мы, наконец, покинули серое царство теории и обратились к практической работе с собакой, пожалуй, оправдывает себя легко понятное объяснение (вместо вышеприведенного слабо измененного философского вырисовывания понятия бесстрашия) той формы
6. ПЛАСТИЧНОСТЬ РЕАЛИЗАЦИИ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ И ПРОБЛЕМА «ИМПРЕССИНГА»
6. ПЛАСТИЧНОСТЬ РЕАЛИЗАЦИИ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ И ПРОБЛЕМА «ИМПРЕССИНГА» Если перейти от содержания наследственной информации к ее реализации (даже если речь идет о самых элементарных, биохимических или морфологических особенностях), то в каждой данной ситуации, в
11.1. Наследование поведения
11.1. Наследование поведения Согласно гипотезе формативной причинности, наследование поведения зависит от генетической наследственности, а также от морфогенетических полей, которые контролируют развитие нервной системы и всего животного в целом, а также от моторных
Движения простейших
Движения простейших Движения простейших отличаются большим разнообразием, причем в этом типе простейших животных встречаются способы локомоции, которые совершенно отсутствуют у многоклеточных животных. Это своеобразный способ передвижения амеб при помощи
Пластичность поведения
Пластичность поведения Поведение кольчатых червей, как и других низших беспозвоночных, характеризуется малой пластичностью, консервативностью. Господствуют врожденные стереотипы («врожденные программы поведения»). Индивидуальный опыт, научение, играет в жизни этих
Ригидность и пластичность в поведении высших позвоночных
Ригидность и пластичность в поведении высших позвоночных Как уже указывалось, вопреки еще распространенному мнению инстинктивное поведение не теряет своей значимости в процессе эволюции, так как оно принципиально не может замещаться научением. Подчеркнем еще раз, что
О срочности поведения
О срочности поведения В нашей книге мы будем много говорить о срочности поведения. Под «срочностью», здесь будет пониматься не необходимость куда-то спешить, а отрезок времени между поступком и его ожидаемым эффектом. Другими словами, срочность поведенческих целей, или,
Понятие поведения
Понятие поведения О психических явлениях объективный наблюдатель судит не непосредственно, а измеряя их проявления в активности живого организма, в первую очередь в поведении. Поэтому остановимся подробнее на этом понятии.Термин «поведение» для обозначения
Склонность к имитации и пластичность поведения представителей поведенческого типа Б
Склонность к имитации и пластичность поведения представителей поведенческого типа Б Способность к социальному обучению различна у представителей разных психологических типов. Еще одним преимуществом особей психологического типа Б является высокая склонность
Пластичность женщин и ригидность мужчин
Пластичность женщин и ригидность мужчин Главные различия в психике и поведении представителей разных полов в том, что женщины пластичны, а мужчины ригидны.Ригидность – это свойство, противоположное пластичности. Мы не используем русскую кальку «твердость» из-за
Глава 1 ФАНТОМНЫЕ КОНЕЧНОСТИ И ПЛАСТИЧНОСТЬ МОЗГА
Глава 1 ФАНТОМНЫЕ КОНЕЧНОСТИ И ПЛАСТИЧНОСТЬ МОЗГА Обожаю дурацкие эксперименты. Я сам их всегда ставлю. Чарльз Дарвин Ещё будучи студентом-медиком, я осматривал пациентку по имени Михи во время моей практики в неврологическом отделении. Стандартное клиническое
Пластичность головного мозга: воздействие окружающей среды
Пластичность головного мозга: воздействие окружающей среды У новорожденного ребенка мозг примерно вчетверо меньше, чем у взрослого человека. Размеры нейронов мозга увеличиваются, а характер нервных связей и сетей усложняется по мере роста ребенка, его общения с людьми