Органы чувств и сенсорные способности
Органы чувств и сенсорные способности
Большой интерес для познания психической деятельности низших многоклеточных беспозвоночных представляет устройство и функционирование их органов чувств, представленных также весьма различными образованиями в соответствии с общим уровнем организации животного.
У наиболее низкоорганизованных представителей беспозвоночных органы чувств еще очень слабо дифференцированы как в морфологическом, так и в функциональном отношении. У этих животных трудно выделить органы осязания, химической чувствительности и т. д. Очевидно, первичные органы чувств вообще были плюромодальными, т. е. они обладали лишь общей, присущей всей живой материи чувствительностью, но в повышенной степени. Существование таких плюромодальных чувствительных клеток является весьма вероятной гипотезой, но в настоящее время такие рецепторные клетки, очевидно, уже не существуют. Специализация таких клеток по отдельным видам энергии привела к появлению унимодальных рецепторных образований, которые, как правило, реагируют лишь на один специфический вид энергии. Так появились термо-, хемо-, механо-, фото- и другие рецепторы.
Согласно этой гипотезе, все органы чувств многоклеточных животных развились из органов осязания — наименее дифференцированных рецепторов. В наиболее элементарных случаях осязательная функция присуща всем клеткам поверхности тела. Но уже у кишечнополостных появляются специальные осязательные клетки, которые, скапливаясь в определенных местах, образуют подлинные органы осязания. Это вытянутые цилиндрические или веретеновидные клетки, несущие на конце неподвижный чувствительный волосок или пучок волосков. Однако эти органы часто выполняют и обонятельную функцию. Особенностью низших многоклеточных беспозвоночных является то, что во многих (хотя и не во всех) случаях эти две рецепторные функции слиты и не поддаются морфологическому разграничению.
В этом нетрудно усмотреть остатки первичной плюромодальности.
С другой стороны, органы зрения относятся к наиболее сложным по строению и функционированию. Иногда органы чувств низшего порядка превращаются в органы чувств высшего порядка (например, у пиявок некоторые из органов осязания — так называемые «сенсиллы» — превращаются в глаза). Известный советский зоолог, специалист по сравнительной анатомии беспозвоночных B-А. Догель говорил в таких случаях о «повышении органа в ранге». Однако в процессе филогенеза, как отмечает Догель, нередко имело место и обратное явление; в других случаях какие-то рецепторы беспозвоночных исчезали, чтобы потом вновь появиться в несколько измененной форме. Это непостоянство и легкость перестройки привели к тому, что у близкородственных беспозвоночных однотипные рецепторы, в частности органы оптической чувствительности, подчас бывают совершенно различными по строению и функциям.
Если взять, к примеру, кишечнополостных, то гидра четко реагирует на свет, хотя специальных органов зрения у нее нет. Она воспринимает свет всей поверхностью тела. Положительный фототаксис гидры выражается в том, что животное производит в освещенной сфере круговые или маятникообразные колебательные движения и в конце концов занимает положение в сторону источника света или даже направляется (ползет) к нему. Свободноживущие же представители кишечнополостных — медузы — обладают уже специальными многоклеточными органами светочувствительности. В простейшем случае эти органы представлены так называемыми глазными пятнами, которые находятся среди обыкновенных эпителиальных клеток, и даже нечетко отграничены от них. Более дифференцированным рецептором является глазная ямка (рис. 32). Однако чаще у медуз встречаются уже настоящие глаза, причем наиболее сложно устроенные из них представляют собой погруженные под слоем эпителиальных клеток глазные пузыри приблизительно шарообразной формы. Эпителий над глазным пузырем утончен и представляет собой прозрачную роговицу. Дно и стенки пузыря состоят из двух типов клеток: ретинальных и пигментных, причем ретинальные клетки снабжены чувствительными палочками. В полости глазного пузыря находится стекловидное тело — студенистая масса, защищающая ретину от механических повреждений. Иногда встречаются даже хрусталик и радужка, и тогда налицо все основные компоненты глаза высших животных (нет, однако, глазодвигательных мышц и систем фокусировки).
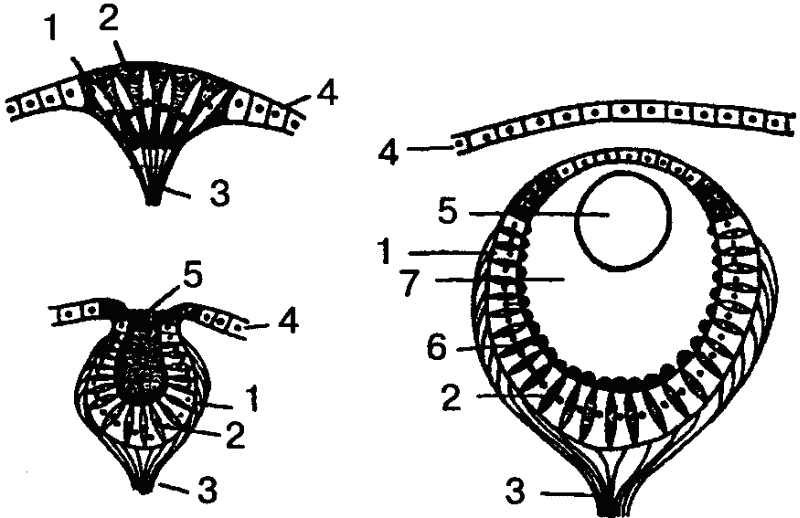
Рис. 32. Развитие глаза беспозвоночных (на примере гидромедуз). Слева вверху — глазное пятно; слева внизу — глазная ямка; справа — глазной пузырь. 1 — зрительная клетка; 2 — пигментная клетка; 3 — зрительный нерв; 4 — эпителий; 5 — хрусталик; 6 — светочувствительные палочки; 7–стекловидное тело (по Бючли)
Учитывая, что глазное пятно является исходной формой вообще всех органов зрения, можно, следовательно, в ряду медуз проследить путь усложнения структуры от самого примитивного органа светочувствительности до сложного, высокодифференцированного глаза.
Очень разнообразны по своему строению и глаза червей, как и других низших многоклеточных беспозвоночных; в принципе к ним относится то же, что говорилось в отношении глаз медуз. В соответствии с многообразием движений кольчатых червей, разнообразием способов добывания пищи и других моментов жизнедеятельности находится и уровень развития сенсорной сферы этих животных. Это, правда, не означает, что у кольчецов имеются рецепторы для всех видов энергии, воздействующих на них, или даже что для всех форм чувствительности имеются специальные органы чувств. Так, например, у кольчатых червей встречаются сложно устроенные глаза, снабженные даже хрусталиками. Имеются весьма сложные глаза и у некоторых планарий и улиток. По их обладатели, насколько известно, неспособны к зрительному восприятию предметов. За исключением, может быть, некоторых улиток, у всех этих животных фотоскопические глаза, позволяющие отличать свет от тьмы и направление, откуда световые лучи падают на животное, а также перемещение светотеней в непосредственной близости от животного. Светочувствительность может при этом быть очень высокой, например, гребешок, двустворчатый моллюск с несколькими десятками глаз, закрывает створки раковины уже при уменьшении интенсивности освещения на 0,3 %. (Для сравнения можно указать, что человек воспринимает уменьшение освещения лишь не менее чем на 1 %).
Большой интерес представляют активно плавающие многощетинковые кольчатые черви из семейства Alciopidae, ведущие хищный образ жизни. У этих полихет глаз не только отличается исключительно сложным строением и величиной, но и снабжен аккомодационным устройством в виде специальных сократительных волокон, способных передвигать хрусталик и тем самым менять фокусное расстояние. Это единственный известный случай среди низших беспозвоночных: аккомодация глаза встречается только у головоногих моллюсков и позвоночных. Возможно, у этих червей в какой-то степени уже существует предметное зрение, что было бы исключением, подтверждающим общее правило. Это относится и к свободно плавающим хищным моллюскам Heteropoda, которые тоже обладают весьма сложно устроенными глазами с приспособлением, заменяющим аккомодацию.
Что же касается дождевого червя, то здесь обнаруживается чрезвычайно интересный факт: у него нет не только сложно устроенных, но и вообще никаких специальных органов светочувствительности. Вместе с тем ему свойствен четкий отрицательный фототаксис. Функцию светоощущения выполняют рассеянные в коже светочувствительные клетки. Это пример кожной светочувствительности низших многоклеточных беспозвоночных. Кожная светочувствительность наблюдается и у многих моллюсков, причем у двустворчатых это нередко единственная форма фоторецепции. Эти моллюски реагируют как на освещение, так и на затемнение чаще всего одинаковым образом — втягиванием выступающих из раковин частей тела или запиранием раковины. Многие улитки реагируют на внезапное затемнение сокращением «ноги», причем эта реакция сохраняется и после экстирпации глаз, что опять-таки указывает на наличие кожной светочувствительности.
Реакция дождевого червя на условия освещения состоит в том, что он уползает в зону большего затемнения. Если же внезапно уменьшить интенсивность освещения, то червь реагирует на это движениями бегства; в естественных условиях он вползает в почву. Такая же реакция следует за внезапным освещением. Если же осветить лишь определенный участок тела дождевого червя, то время этой реакции сокращается пропорционально размеру освещаемой площади поверхности тела — в наибольшей мере червь реагирует при освещении всей его поверхности. Следовательно, реакция определяется градиентом раздражения освещенных и неосвещенных частей тела. Аналогично реагируют на свет и другие кольчатые черви.
Кольчатые черви реагируют также на прикосновения, химические и термические раздражения, силу тяжести, электрические раздражения, течение воды, а наземные формы (дождевые черви) — на влажность среды. Однако принципиальных отличий от реакции на свет нет: все эти реакции находятся на одном уровне и характеризуются тем, что являются ответами на отдельные раздражители, на отдельные признаки, качества предметов, но не на сами предметы как таковые. Так, например, много- и малощетинковые черви проявляют четко выраженные таксисные реакции на тактильные раздражения. Преобладают отрицательные реакции, но в ряде случаев наблюдаются и положительные тигмотаксисы: прикосновения нервных сегментов животного к субстрату влекут за собой прижимание к нему всем телом, что имеет, конечно, большое значение при роющем образе жизни или жизни в трубках. Интересно, что у дождевого червя соответствующие рецепторные образования представлены лишь отдельными чувствительными клетками, разбросанными по всему телу, но более густо располагающимися на его переднем конце. У многощетинковых же червей органами осязания часто являются щупальца или щетинки.
Хорошо развита и химическая чувствительность, причем в большинстве случаев наблюдается отрицательный хемотаксис. При большой интенсивности химического воздействия черви всегда реагируют отрицательно. С другой стороны, дождевой червь, например, способен по химическим признакам выбирать разные виды листьев, что свидетельствует о специализации в сенсорной сфере. У ряда кольчецов обнаружены расположенные около ротового отверстия органы химической чувствительности в виде ямок. Особенно это относится к пларающим видам, у которых имеется пара таких ямок, выстланных мерцательным эпителием.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Органы чувств у рыб
Органы чувств у рыб Нельзя допустить, что рыбы не наделены зрением, что они не слышат, не имеют обоняния и осязания, не ощущают вкуса. Рыбам присущи все перечисленные пять чувств, у них имеются и соответствующие органы этих чувств. Кроме того, считается, что у рыб есть и
§ 13. Рецепторы и органы чувств
§ 13. Рецепторы и органы чувств Органы чувств по источникам воздействий можно разделить на эндогенные и экзогенные. Первые специализируются на рецепции внутренней среды и органов животного, а вторые информируют о внешней среде. Оба источника информации крайне важны для
§ 43. Нервная система и органы чувств птиц
§ 43. Нервная система и органы чувств птиц Нервная система птиц состоит из центрального и периферического отделов. Головной мозг птиц крупнее, чем у любых современных представителей рептилий. Он заполняет полость черепа и имеет округлую форму при небольшой длине (см. рис.
§ 48. Органы чувств млекопитающих
§ 48. Органы чувств млекопитающих Органы чувств млекопитающих имеют ряд особенностей, которых нет у позвоночных других групп. У млекопитающих хорошо развит орган обоняния. В отличие от птиц он состоит из основного органа обоняния и вомероназальной системы. Основной
9.2. Сенсорные системы
9.2. Сенсорные системы Сенсорные системы возникают на очень ранних стадиях эволюции животных для непосредственного восприятия сигналов внешнего мира. В ходе этой эволюции постепенно совершенствуется воспринимающая структура, формируются аналитические центры, из
1.2. Сенсорные пороги
1.2. Сенсорные пороги АБСОЛЮТНАЯ ЧУВСТВИТЕЛЬНОСТЬ СЕНСОРНОЙ СИСТЕМЫ Абсолютная чувствительность сенсорной системы основана на её свойстве обнаруживать слабые, короткие или маленькие по размеру раздражители. Абсолютную чувствительность измеряют порогом той или иной
Жар чувств
Жар чувств Из 839 американцев и японцев, участвовавших в нашем «любовном» исследовании, 89 % мужчин и 79 % женщин согласились с утверждением: «Когда я уверен (а) в том, что ___ любит меня, я словно парю в воздухе» (см. п. 32 приложения). Пожалуй, ни один признак романтической страсти
Исследование органов чувств
Исследование органов чувств У собак наиболее сильно развито чувство обоняния. Для исследования обоняния берут хлеб или мясо и помещают в посуду с плотно закрытой крышкой и определяют, с какого расстояния полуголодное животное чует запах, если открыть крышку. Ослабление
ОРГАНЫ ЧУВСТВ
ОРГАНЫ ЧУВСТВ Говоря об органах чувств, для собак наиболее важным следует признать хорошее зрение, особенно для рабочих пород. Глаз представляет собой сложный орган, в нем может развиться множество дефектов, и он легко доступен для подробного исследования. Возможно, по
Сенсорные способности, таксисы
Сенсорные способности, таксисы Специфические условия, в которых живут насекомые, далеко идущее приспособление двигательной активности при обилии и разнообразии качественно весьма различных агентов среды, управляющих их поведением, — все это обусловило появление у
Сенсорные особенности
Сенсорные особенности В полном соответствии с высокоразвитыми эффекторными системами высших позвоночных находится и высокий уровень их сенсорных способностей. Наибольшее значение имеют органы слуха и равновесия (внутреннее ухо, начиная с земноводных — дополнительно
Мы не рабы своих чувств
Мы не рабы своих чувств Может показаться, что склонность к галлюцинациям – слишком дорогая цена за способность нашего мозга строить модели окружающего мира. Неужели нельзя было настроить систему так, чтобы сигналы, поступающие от органов чувств, всегда играли главную
СЕНСОРНЫЕ СПОСОБНОСТИ ДИКИХ ЛЮДЕЙ
СЕНСОРНЫЕ СПОСОБНОСТИ ДИКИХ ЛЮДЕЙ Кроме телесной и физиологической перестройки организма у диких людей под давлением изменившихся условий существования начинают развиваться новые психосенсорные адаптации. Как показали наблюдения, снежный человек ощущает
10. Органы чувств
10. Органы чувств Организм живет в постоянно изменяющихся условиях внешней среды с бесчисленным количеством всевозможных раздражителей. Одни из них не имеют никакого отношения к организму и не являются для него сигналами для соответствующего поведения. Другие же