Система и филогения
Последнее сорокалетие — время попыток построить «большую систему». Конечно, система должна быть естественной, т. е. отражать филогению соответствующих групп животных. Ряд таких систем предложен, то ни одна из них не оказалась общепринятой: у каждой есть сторонники и противники, и притом обычно эти оба лагеря примерно одинаковы. А такое «разделение голосов» — нехороший признак.
Главнейшие моменты, положенные в основу новых систем, — преимущественно факты эмбрионального развития. Эти факты привели к созданию соответствующих «теорий» происхождения тех или иных групп животных, что и нашло свое отражение в классификационных схемах.
У всех многоклеточных животных эктодерма образуется из наружного зародышевого листка, энтодерма — из внутреннего. Личинка губок двуслойна: наружный слой соответствует эктодерме и состоит из жгутиковых клеток, внутренний слой, соответствующий энтодерме других многоклеточных, образован более крупными, зернистыми клетками. При метаморфозе происходят оригинальные явления: наружный слой («эктодерма») попадает внутрь, а внутренний («энтодерма») — наружу. Такое «наоборот» позволяет некоторым зоологам противопоставлять губок всем остальным многоклеточным животным и делить «подцарство многоклеточных» на два раздела — настоящие многоклеточные (Eumetazoa, Enterozoa) и пара-многоклеточные (Parazoa), куда входят только губки. Это противопоставление, вполне законное в определительной таблице, вряд ли отражает родственные связи губок и во всяком случае не может служить основанием для выделения их в особый «раздел» столь высокого порядка, т. е. для приравнивания их ко всем остальным многоклеточным. Как никак, а «извращение» зародышевых листков личинки губок — своеобразное «уродство». А уродства, даже и очень выдержанные в пределах группы, нельзя противополагать «нормальному». Другие особенности губок не выходят из пределов особенностей типового характера.
Лучистая и двусторонняя симметрия (преимущественно нервной системы) находит отражение и в некоторых современных классификациях. Говорить об искусственности таких подразделений не приходится.
Важное значение имеет способ образования окончательного ротового отверстия, «дефинитивного рта». У первичноротых (Protostomia) окончательный рот возникает непосредственно в области «первичного рта», т. е. бластопора, или гастропора (места как бы перехода первичной эктодермы в энтодерму, где нередко бывает и первичное ротовое отверстие) гаструлы. У вторичноротых (Deuterostomia) окончательный рот возникает заново, а бластопор обычно становится анальным отверстием. Есть и другие различия между первично- и вторичноротыми. У первичноротых кровеносная система незамкнутая, нервная система лестничного типа, хорошо развита кутикула: дробление детерминированное (в самых ранних стадиях развития зачатки определенных органов уже распределены между отдельными бластомерами — клетками, образовавшимися в результате дробления яйцевой клетки, т. е. бластомеры качественно различны); мезодерма (кроме плеченогих) развивается из телобластов (обособленных бластомеров). У вторичноротых кровеносная система в основном замкнутая, центральная нервная система никогда не бывает лестничной; мезодерма образуется энтероцельным путем (первичный кишечник образует боковые симметричные выпячивания вроде мешков-карманов); дробление чаще недетерминированное (бластомеры долгое время остаются недиференцированными, т. е. одинаковыми качественно). Первичноротые — это членистоногие, моллюски, черви и большинство «червеобразных» (моллюсковидных старых авторов). Вторичноротые — хордовые, иглокожие и некоторые из «червеобразных». К числу различий между первично- и вторичноротыми относятся и весьма любопытные физиологические особенности. Мышцы тех и других содержат соединения фосфора, различные в обеих группах: для первичноротых характерны соединения типа аргинино-фосфата, тогда как у вторичноротых преобладают креатино-фосфаты. Иглокожие и кишечнодышащие обладают обоими типами фосфатов.
Гаструла ракообразного (Chondracanthus) в продольном разрезе:
1 — бластопор (первичный рот); 2 — эктодерма; 3 — энтодерма; 4 — клетки, дающие мезодерму.
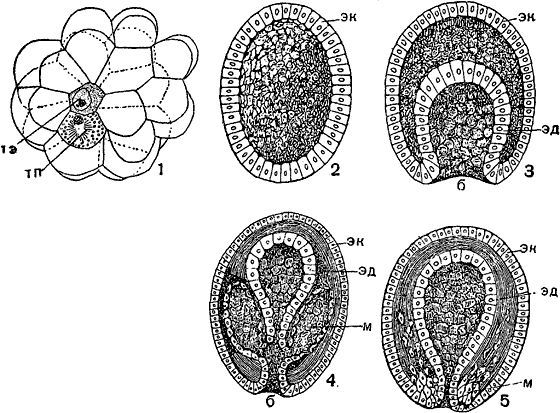
Различные типы образования мезодермы:
1 — зародыш сагитты (Sagitta) на стадии 32 бластомеров; ясно виден зачаток энтодермы (тэ, телобласт энтодермы) и половой зачаток (тп); 2 — бластула; 3 — гаструла; 4 — образование мезодермы путем парного выпячивания стенок первичного кишечника (энтероцельный способ); 5 — образование мезодермы путем врастания клеток, образующих позже мезодермальные полоски (эктероцельный способ); м — мезодерма, эд — энтодерма, эк — эктодерма.
Затем можно говорить о двуслойных (Diploblastica), не имеющих типично развитой мезодермы (губки, кишечнополостные), и трехслойных (Triploblastica), обладающих таковой (все остальные многоклеточные).
Полость тела (целом, coelom) — это полость, лежащая между кишечником и стенками тела; она есть не у всех животных. Так, у кишечнополостных можно увидеть только кишечную полость. Различают первичную и вторичную полости тела. Первичная полость тела — остатки полости бластулы, производные мезодермы развиваются внутри нее. Вторичная полость тела возникает внутри зачатков мезодермы, окружена производными мезодермы, имеет, как правило, эпителиальную выстилку, под которой развиваются половые продукты. Вторичная полость тела может образоваться различными путями. При энтероцельном способе первичный кишечник дает выпячивания, которые позже от него отделяются; стенки выпячивания образуют мезодерму, а их полости — вторичную полость тела. Животных с таким способом образования вторичной полости тела называют «энтероцельными» (Enterocoelia). Им противопоставляют животных «эктероцельных» (Ecterocoelia), у которых мезодерма и вторичная полость тела образуются не путем выпячивания стенок первичного кишечника, а как-либо иначе, обычно телобластическим способом, т. е. из рано обособившихся бластомеров (образуются две полоски мезодермы, которые потом расщепляются или раздвигаются, получившаяся щель и кладет начало полости).
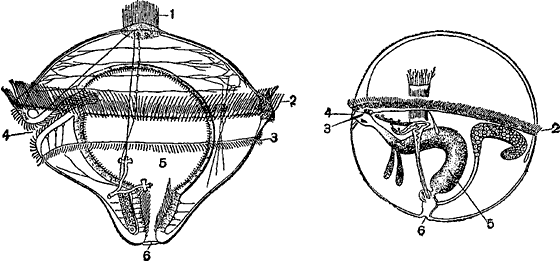
Личинка червя Polygordius, представляющая типичную трохофору (налево) и шарообразная коловратка Trochosphaera (направо):
1 — теменной пучок неподвижных ресничек; 2 — предротовой венчик ресничек; 3 — послеротовой венчик; 4 — рот; 5 — кишка; 6 — анальное отверстие.
Некоторые зоологи придают очень большое значение соотношениям главной оси тела и оси гаструлы, различая первичноосных (Protaxonia) и разноосных (Heteraxonia). У первичноосных (губки, кишечнополостные) главная ось тела (орально-аборальная ось, т. е. идущая от ротового отверстия к противоположному концу тела) взрослого животного соответствует главной оси гаструлы, соединяющей бластопор с противоположным концом тела. Это неудивительно, так как грубо схематизированное кишечнополостное — своего рода «пожизненная гаструла». У разноосных главная ось тела расположена под углом к главной оси гаструлы; таковы все остальные многоклеточные животные, кроме иглокожих и кишечнодышащих.
Большие надежды возлагались в свое время на «трохофорную теорию», связанную с личинкой трохофорой, очень напоминающей взрослую коловратку. Наличие трохофоры или трохофорообразной стадии связывало плоских червей и коловраток с кольчатыми червями и моллюсками, а Семпер (1875) и А. Дорн (1875), видевшие в кольчецах исходную точку позвоночных, втянули в этот «круг» и последних. В настоящее время большинство зоологов не придает трохофоре такого крупного значения, но свою, и очень полезную, роль эта теория сыграла. Между прочим, наличие двустороннесимметричной нервной системы, обычно с многими парами узлов, — признак, очень часто сопутствующий трохофоре. Отсюда объединение кольчецов, моллюсков, моллюсковидных (червеобразных), членистоногих, плоских червей и немертин в группу Zygoneura.
Те или иные из указанных особенностей строения или развития и положены разными зоологами в основу составленных ими классификаций. Эти классификации довольно многочисленны и иногда резко разнятся между собой. Особенности их в большинстве заключаются в группировках типов, т. е. в числе и объеме «надтиповых» объединений, отчасти и в объеме самих типов. Классы, как правило, остаются почти неизмененными, хотя иногда и меняют свое место. Один тип хордовых почти всюду приводится одинаково, и только низшие хордовые у некоторых авторов включаются в другие типы.
Наличные системы можно разделить на две группы: одни зоологи придают огромное значение первично- и вторичноротости и способу закладки вторичной полости тела, другие относятся к этим признакам довольно равнодушно, полагая, что здесь могут сказываться и явления конвергенции, чт? лишает такой признак значения «филогенетического абсолюта». Этот второй взгляд проведен в ряде крупных иностранных руководств, а у нас — в учебнике Холодковского. Отказать в вполне основательной осторожности таким зоологам трудно: действительно, при наличии всего двух, самое большее трех возможных способов образования окончательного рта или вторичной полости тела явлениям конвергенции открывается здесь самое широкое поле. И затруднительность размещения в системе некоторых «уклоняющихся» групп, может быть, является результатом как раз конвергентного образования некоторых признаков, наличие которых резко противоречит всем остальным особенностям строения данной группы.
Арнольд Ланг (A. Lang, 1855–1014), ученик и последователь Э. Геккеля, профессор Иенского, а позже Цюрихского университетов, создал ряд филогенетических теорий. Наиболее интересны его обобщения, касающиеся различных групп червей: теория о происхождении турбеллярий от ктенофорообразных предков, «турбеллярная теория» происхождения кольчатых червей, особая теория происхождения кровеносной системы. В 1888–1894 гг. он издал руководство «Учебник сравнительной анатомии беспозвоночных животных»[93], в котором и предложил свою систему: 7 типов, без более «высоких» объединений.
1. Простейшие (Protozoa) с 6 классами: Monera, Sarcodina, Flagellata, Gregarinida, Infusoria, Suctoria.
2. Зоофиты, или кишечнополостные (Zoophyta, или Coelenterata) с 8 классами: гастреады (Gastreada), губки (Porifera) и стрекающие (Cnidaria).
3. Плоские черви (Plathelminthes), 3 класса: турбеллярии (Turbellaria), сосальщики (Trematoda), лентецы (Cestoda).
4. Черви (Vermes), 6 классов: немертины (Nemertini), круглые черви (Nemathelmia), кольчецы (Annulata), переднеанальные (Prosopygii, т. е. червеобразные; сюда отнесены также приапулиды, сипункулиды и перистожаберные — Priapulidae, Sipunculidae, Pterobranchia), коловратки (Rotatoria), щетинкочелюстные (Chaetognatha).
5. Членистоногие (Arthropoda) разделены на 2 подтипа:
а) Жабернодышащие (Brancbiata), всего 1 класс ракообразных (Crustacea), к которому как добавления отнесены трилобиты (Trilobita), ракоскорпионы (Gigantostraea), гемиаспиды (Hemiaspida), мечехвосты (Xiphosura) и многоколенчатые (rantopoda).
б) Трахейнодышащие (Tracheata), 3 класса: первичнотрахейные (Protracheata), усиконосные (Antennata, т. е. многоножки и насекомые) и хелицероносные (Chelicerata). Тихоходки (Tardigrada) помещены как добавление к типу.
6. Моллюски (Mollusca): 5 обычных классов.
7. Иглокожие (Echinodermata): 5 обычных классов.
В 1913 г., в редактированном им «Руководстве по сравнительной анатомии беспозвоночных животных» Ланг внес в эту систему небольшие изменения: разделил тип простейших на 2 подтипа, членистоногих — на 3 подтипа, а немертин перенес в тип плоских червей.
Француз Эдмон Перрье (Е. Perrier, ум. 1925) полагал, что всякий организм — результат образования последовательных «ассоциаций» разных ступеней. Все животные, по Перрье, были раньше колониями, которые, путем взаимного приспособления составляющих их организмов, медленно и постепенно сделались организмами высшего порядка. По существу это комбинация «лестницы» с некоторыми натурфилософскими «кругами». Всех ступеней развития у Перрье четыре: 1) пластиды — первоначально одноклеточные организмы, как материал для постройки органического мира, 2) мериды — ассоциации пластид, 3) ассоциации мерид — зоиды, 4) ассоциации зоид — демы. Конечно, эти любопытные соображения и легли в основу системы Перрье, о которой даже очень снисходительные зоологи писали, что она «является несколько отсталой по своему времени».
Э. Перрье («Trait? de zoologie», 1893) принимает три ступени организации и четыре типа строения.
A. Ступень Простейшие (Protozoa), с двумя типами строения:
I. корненожки (Rhizopoda) с единственным типом того же наименования (1) и II. «кругоживотные» (Perizoa) с тремя типами: 2) Megacystida, 3) Sporozoa (споровики), 4) Infusoria (инфузории вместе с жгутиковыми).
Б. Ступень Мезозои (Mesozoa).
B. Ступень Многоклеточные (Metazoa). Разделены на два типа строения: животнорастения (Phytozoa) и настоящие животные (Artiozoa).
III. Животнорастения (Phytozoa) разделены на три серии:
а) Spongia (губки) с двумя типами: 5) Calcarea (известковые) и 6) Silicosa (кремневые губки).
б) Polypae (полипы) с тремя типами: 7) Hydromedusae (полипы и медузы), 8) Anthozoa (коралловые полипы) и 9) Ctenophora (ктенофоры).
в) Echinodermata (иглокожие) с двумя типами: 10) Anangia (звезды и офиуры), 11) Angiophora (остальные иглокожие).
IV. Настоящие животные (Artiozoa):
а) Серия Chitinophora (хитиновые), 2 типа: 12) Arthropoda (членистоногие), 13) Nemathelminthes (круглые черви вместе с щетинкочелюстными).
б) Серия Nephridia (нефридиевые), 6 типов: 14) Lophostoma (коловратки, мшанки, плеченогие), 15) Vermes (черви плоские, немертины, кольчатые; также брюхоресничные и кишечнодышащие), 16) Mollusca (моллюски), 17) Tunicata (оболочники), 18) Vertebrata (позвоночные).
Четыре «типа развития» — четыре последовательные ступени «ассоциаций». Типов Перрье принимает 18, но его «типы» то соответствуют типам других систем (12, 16, 17), то слишком объемисты (15), то не превышают объема класса (1, 5, 6). Последовательность также очень оригинальная: в 10-м и 11-м типах — иглокожие, в 12-м — членистоногие, в 13-м — круглые черви, а в 14-м, в числе прочих, — плоские черви. Настоящее место этой «системе» — в ряду систем Боннэ, Ламарка и других «лестниц».
Отто Бючли (О. Butschli, 1848–1920) 42 года пробыл профессором зоологии в Гейдельберге. Он автор теории ячеистой структуры протоплазмы (прекрасный химик, он пытался обосновать эту теорию и физико-химически), знаменитый знаток простейших, один из крупнейших цитологов (установил полностью процесс кариокинеза в животной клетке, нашел ядро у некоторых бактерий). Любовь к простейшим отразилась и на системе, предложенной им в 1910 г. в его «Лекциях по сравнительной анатомии»[94]. Бючли разделил простейших на 4 типа, хотя в его системе всего только 11 типов.
I. Подцарство Protozoa (простейшие).
1) Sarcodina (саркодовые), 2) Mastigophora (жгутиковые), 3) Sporozoa (споровики), 4) Infusoria (инфузории).
II. Подцарство Metazoa (многоклеточные).
А. Раздел (Kreis) Spongia (губки); 5) Porifera (губки), 3 класса.
Б. Раздел (Kreis) Eumetazoa (настоящие многоклеточные).
а) Подраздел Radiata (лучистые): 6) Coelenterata (кишечнополостные) с добавлением Planuloidea.
б) Подраздел Bilateralia (двустороннесимметричиые): 7) Vermes (черви) разделены на 3 подтипа, сюда же отнесены и все червеобразные, 8) Arthropoda (членистоногие), разделены на 3 подтипа, 9) Mollusca (моллюски), 10) Echinodermata (иглокожие), 11) Chordata (хордовые).
В этой системе черви соединены в один тип, причем туда же отнесены все червеобразные (Vermioidea), коловратки, кишечнодышащие. Деление червей на три подтипа, основанное на «членистости», — нечленистые, или амерные, малочленистые (олигомерные), многочленистые (полимерные) — очень искусственно, хотя и сохранено некоторыми зоологами нашего времени, очевидно, исключительно из соображений «практичности».

О. Бючли (1848–1920).
Еще объемистее оказался «тип червей» в системе, предложенной Рихардом Гертвигом (R. Hertwig). Гертвигов два брата-биолога — Оскар (1849–1922) и Рихард (род. 1850). Оба они ученики Геккеля, Гегенбаура и М. Шульце. Первое время работали вместе и тогда, между прочим, создали «теорию целома», имевшую большое значение в развитии эмбриологии и в постройках филогенетических «древ» в конце XIX в. Тогда же братья немало занимались исследованием морфологии медуз; актиний и щетинкочелюстных. Позже они разъехались: Оскар — профессором в Берлин, Рихард — в Мюнхен. Исследуя оплодотворение у морских ежей, Оскар заинтересовался ролью ядра и развитием яиц и зародышей различных животных, что привело его к созданию теории «биогенеза» (эпигенетическая теория развития организма, но совсем не теория «происхождения жизни», как можно подумать из-за названия «биогенез»). Он отрицает биогенетический закон, отрицает теорию дарвиновского отбора, так как свойства организма, по мнению О. Гертвига, суть «необходимые следствия определенных причин». Виды развились путем постоянного и закономерного прогрессирования, а вовсе не как «игрушка случая». О. Гертвиг был противником Дарвина и потому, что дарвинизм, по его мнению, этически и политически вредное учение: он революционен и приводит к социализму. Такая трактовка учения Дарвина становится особенно занятной, если сопоставить ее с трактовкой Геккеля, учителя Гертвига, видевшего в отборе нечто «аристократическое» и совсем не собиравшегося устраивать революции. Наоборот, Геккель, социал-дарвинист, видел в дарвинизме своего рода противоядие от покушений социалистов на классовое общество.

Р. Гертвиг (род. 1850).
Рихард Гертвиг много работал по цитологии, по теории оплодотворения, занимался систематикой корненожек, радиолярий, инфузорий, решал проблему «смерти и бессмертия». Он написал большой «Учебник: зоологии» (1891)[95], вышедший в 1924 г. уже 14-м изданием (есть два русских перевода). Р. Гертвиг принимает только 7 типов.
1. Простейшие (4 класса). — 2. Кишечнополостные (с включением подтипа губок). — 3. Черви (3 подтипа, с включением как добавления всех червеобразных, кишечнодышащих, оболочников). — 4. Моллюски. — 5. Иглокожие. — 6. Членистоногие. — 7. Позвоночные.
Гертвиговские «черви» включают не только червей, начиная от турбеллярий, но и кишечнодышащих и даже оболочников. Причины такого объема типа и разрыва между кишечнодышащими — оболочниками и иглокожими — позвоночными кроются во взглядах Р. Гертвига на происхождение позвоночных: он ищет перехода к ним внутри типа червей, который и принимает поэтому в таком широком объеме, не замечая ряда несообразностей, вытекающих из этого.
Н. А. Холодковский (1858–1921, ср. стр. 284), автор крупнейшего до последнего времени русского учебника зоологии[96], дает систему, заметно отличающуюся от гертвиговской. Холодковский принимает 9 типов, выделяя губок и червеобразных в особые типы: 1) простейшие, 2) губки, 3) кишечнополостные, 4) иглокожие, 5) черви (пять подтипов), 6) червеобразные, 7) моллюски, 8) членистоногие, 9) хордовые (оболочники, бесчерепные, позвоночные).
Здесь кишечнодышащие сближены с иглокожими, но эти чрезвычайно удалены от хордовых. Что касается червей, то объединенные в один подтип коловратки и кольчатые черви (да еще и гефиреи) показывают, что «трохофорная теория» пользовалась достаточной симпатией Н. А. Холодковского, — только этим можно объяснить попытку такого объединения.
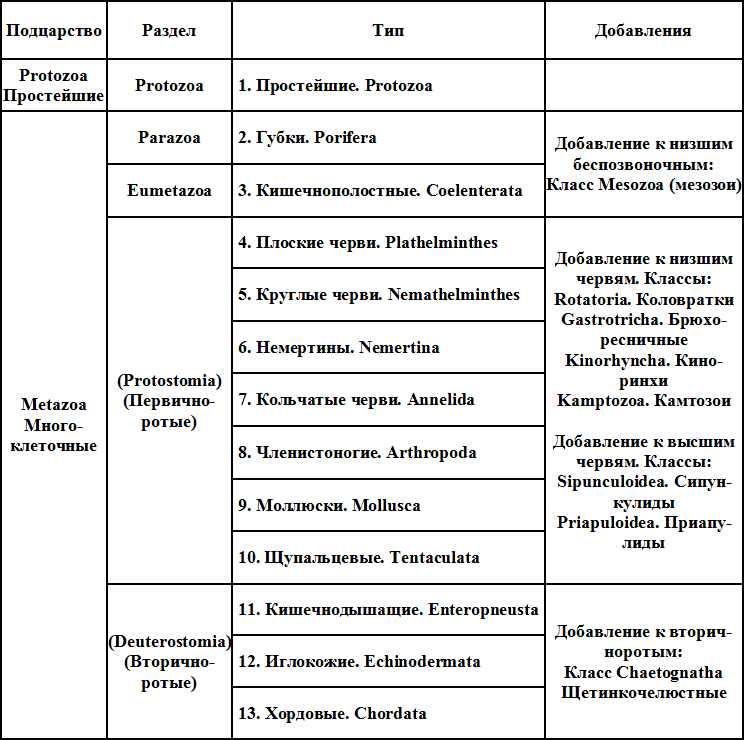
В. Кюкенталь (W. K?kenthal), основатель и первоначальный редактор крупнейшего из существующих руководств по зоологии («Handbuch der Zoologie»), вначале (с 1898 г.) принимал систему Бючли с небольшими поправками, а именно — давал червей в основном по А. Лангу. Но позже он принял для червей деление, предложенное Бючли (три подтипа). Всех типов у Кюкенталя 9. Простейшие принимаются за один тип (2 подтипа — инфузории и все остальные), кишечнополостные также разбиты на 2 подтипа (ктенофоры и стрекающие), черви — 3 подтипа по образцу Бючли. Членистоногие разделены на 6 подтипов; ракообразные, хелицероносные (мечехвосты и паукообразные), многоколенчатые, прогонеаты (сколопендреллы, пауроподы, двупарноногие), губоногие (один класс) и насекомые. Остальные типы интереса не представляют. Наиболее любопытно деление членистоногих на 6 подтипов, вместо обычных трех. Кюкенталь приводит и ряд «надтиповых» группировок. Подцарство Protozoa охватывает один тип простейших. Второе подцарство, Metazoa, разбито на два раздела: Parazoa (губки) и Eumetazoa (остальные многоклеточные), которые в свою очередь разделены на подразделы лучистых (кишечнополостные и ? мезозои) и двустороннесимметричных (все остальные типы, начиная с червей).
Примером эклектической системы может служить система, проведенная в «Курсе зоологии» коллектива авторов под редакцией проф. Б. С. Матвеева (8-е изд. в 1940 г.). Хотя во «Введении» и приведена схема «крупнейших подразделений животных», где указаны такие группировки, как двуслойные, трехслойные, первично- и вторичноротые, а главное — дан последовательный порядок типов (сообразно этим группировкам), в тексте типы перечисляются в ином порядке, отличающемся от такового у Холодковского только тем, что иглокожие перенесены в конец ряда. Конечно, первично- и вторичноротость оказались выдержанными, поскольку иглокожие отнесены к концу, но червеобразные, занимающие «среднее место» между этими крупными группировками, помещены среди первичноротых. Разные типы написаны разными авторами, а потому и система, проведенная в учебнике, не совпадает в целом ни с одной из крупных принятых систем. Так, простейшие — упрощенная система Бючли, членистоногие очень напоминают таковых Холодковского, а для многоножек принято старое обозначение (все в одном классе), правда, с оговоркой, что такая группировка многоногих неестественна. Вот эта система:
1. Простейшие (2 подтипа, 5 классов). — 2. Губки (5 отрядов, что неправильно: непосредственно на отряды тип разбивать нельзя; качественно приведенные отряды неравноценны). — 3. Кишечнополостные (2 подтипа: стрекающие и гребневики). — 4. Плоские черви (4 класса). — 5. Круглые черви (брюхоресничные, нематодовые, волосатиковые, коловратки, киноринхи). — 6. Немертины. — 7. Кольчатые черви (5 классов, как добавление — мизостомиды). — 8. Червеобразные (4 класса). — 9. Моллюски (5 классов). — 10. Членистоногие (3 подтипа, 7 классов). — 11. Иглокожие (2 подтипа, 5 классов). — 12. Хордовые (3 подтипа).
Вторая группа классификаций — попытки соединить в одной схеме порядковое перечисление типов, филогенетическое «древо», а поскольку признаки высших групп даются в виде дихотомии, то и определительную таблицу. Основателями такого метода можно считать Гатчека, Гроббена, отчасти и К. Клауса, А. Гетте, но на деле кое-что в этих системах сильно напоминает Ламарка (дифелитическое происхождение многоклеточных), а корни «надтиповых» группировок нередко можно найти и у натурфилософов середины XIX в. (правда, характеристики групп иные, но это мало меняет сущность дела).
Еще в 1888 г., в 1-м издании своего «Учебника зоологии»[97] Б. Гатчек (В. Hatschek) предложил такое деление для многоклеточных животных:
Первичноосные (Protaxonia) — Губки (Spongiaria), Стрекающие (Cnidaria), Ктенофоры (Ctenophora).
Разноосные (Heteraxonia) (они же двустороннесимметричные, или полостные, Bilateralia, Coelomata) — Трохофорные (Zygoneura: черви, червеобразные, моллюски, членистоногие), Иглокожие (Ambulacralia), Хордовые (Chordonia).
Для трохофорных (Zygoneura) характерна та или иная связь с трохофорой (именно Гатчек создал «трохофорную теорию»), а также двустороннесимметричная нервная система обычно с многими парами узлов. В связи с тем, какова связь с трохофорой, Гатчек разделил «трохофорных» на две группы: 1) Autoscolecida — трохофоре соответствует общая организация животного (плоские и круглые черви, немертины, коловратки) и 2) Aposcolecida — трохофора является только ранней стадией развития (кольчатые черви, червеобразные, моллюски, членистоногие).
В 1911 г. Гатчек опубликовал новую систему[98], в которой резко отграничил ктенофор (Ctenophora) от остальных кишечнополостных и заметно изменил принцип высших группировок, причем дал совершенно иной, чем принято, объем типам. Надтиповые группировки Гатчека основаны на «плане строения», т. е. по существу соответствуют «типам» Кювье, хотя и не совпадают с ними в объеме. Можно найти в этих группировках и нечто общее со «ступенями развития» старых зоологов: и тут и там сказывается желание реализовать в виде системы чисто «идеальные» представления.
Система Гатчека (1911)
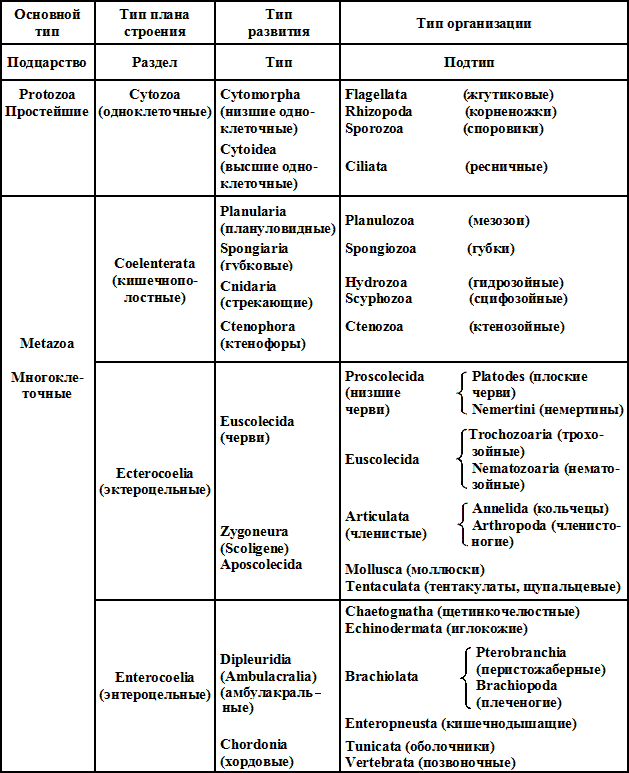
Присмотревшись к системе Гатчека (1911), легко заметить, что общепринятое понятие «тип» у него подменено иным и что «тип» других авторов (в объемном смысле) у него гуляет по трем графам, т. е. по всем крупным группировкам за исключением подцарств. Помещенные в графе «тип развития», она же «тип» (Phylum), группы только в двух случаях из 11 совпадают с обычными объемами типов (Spongiaria, Chordonia). Подтипы четвертой графы — пестрая компания типов, подтипов и даже классов (в обычном их понимании). Странная пестрота и весьма последовательно проведенное не изменение, а искажение объемов типов, т. е. полнейшая перетасовка крупных таксономических категорий, — неизбежное следствие методов построения системы Гатчека. Приняв такие предпосылки, как «тип плана строения», «тип развития» и «тип организации», он естественно должен был расценивать группы сообразно этим «качественным» приметам. А это неминуемо повлекло за собой изменения объемного порядка. Если об естественности гатчековской системы в смысле порядка следования крупных категорий можно спорить, то об «естественности» ее в смысле объемов категорий (таксономические достоинства системы) спорить не стоит, — эта сторона системы никуда не годится.
В вышедшем в 1916 г. «Учебнике зоологии» К. Клауса и Гроббена[99] проведена еще более запутанная в таксономическом отношении система. Правда, здесь авторы не говорят о типах и планах строения и развития, но по существу проводят то же самое, подобно Гатчеку называя «типом» совсем не «тип авторов», а именно тип развития Гатчека. Гатчековский же «тип организации» проведен здесь в виде «ряда», правда, при заметно укрупненных группах. И наконец, в качестве «класса» приводятся отчасти действительные классы, отчасти же «подтипы» и всякого рода промежуточные между подтипом и классом группировки. Снова таксономика дана в некотором «новейшем» понимании, которое сводится к искажению большинства крупных группировок. Как и у Гатчека, принимается дифелитическое происхождение большинства многоклеточных. По существу это системы одного порядка, построенные путем применения одних и тех же методов, и известные расхождения, заметно сказывающиеся в схемах внешне, мало отражаются на их внутреннем сходстве. Трудно удержаться от воспоминаний о К. Карусе, Мак-Лее и прочих «кунстаторах» по части построения систем в виде колец и концентрических кругов, когда смотришь на «графы» Гатчека и Клауса с Гроббеном. Те старались «отразить» в системе некий «единый план», эти… эти делают по существу то же самое, только вместо «плана» нужно поставить «родословное древо».
Близка до известной степени к гроббеновской системе и система, предложенная Д. М. Федотовым (1935)[100], но она сильно выигрывает своей большей простотой. Д. Федотов принимает всего 10 типов, типов обычного объема, за исключением типа червей: все черви объединены в одном типе (кроме червеобразных, — эти выделены). Этот «тип червей» придает схеме несколько странный вид: с одной стороны — первично- и вторичноротые, с другой — архаические «черви». Д. Федотов делит подцарство многоклеточных на 2 раздела: лучистые и двустороннесимметричные, тем самым противополагая губок и кишечнополостных всем остальным многоклеточным. Основание для такого противопоставления он видит в типе симметрии тех и других и в особенностях взаимоотношений главной оси тела взрослого животного и гаструлы. Для двустороннесимметричных принимается дифелитическое происхождение (не стоит останавливаться на щупальцевых, так как они наверное попадут в конце концов в одну из двух групп, и третья ветвь — сейчас их три — будет ликвидирована).
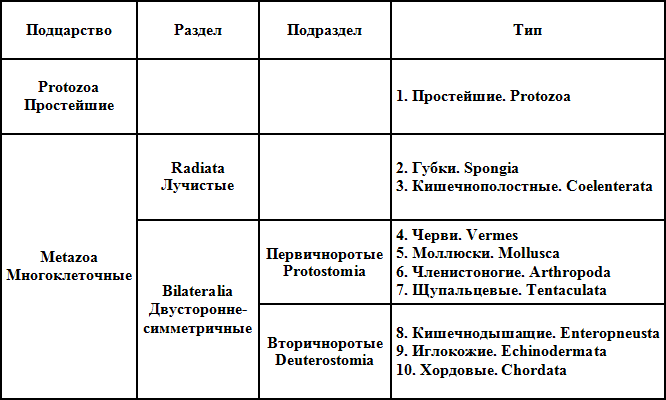
Наконец, в коллективном многотомном «Руководстве по зоологии», издаваемом при участии Зоологического института Московского университета, принято 13 типов. Система, проведенная здесь (вышел пока только I том, 1937), — комбинация систем Клауса-Гроббена и Федотова плюс некоторые изменения, внесенные редакторами «Руководства». Подцарство многоклеточных разделено на два крупных «раздела»: губки противопоставлены всем остальным многоклеточным. Такое противопоставление вряд ли резонно, но оно неизбежный шаг, если не принимать группировки «лучистых», объединяющей губок и кишечнополостных: третья возможная комбинация — губки, кишечнополостные, первичноротые, вторичноротые — была бы явной несообразностью. Любая из возможных комбинаций (губки — остальные многоклеточные; губки — кишечнополостные — остальные многоклеточные, точнее — первично- и вторичноротые) неудачна, так как чисто таксокомически неверна. Причина неудач — желание ввести в классификационную схему специфические моменты «родословного древа», т. е. моменты, прекрасные для схемы только филогенетической, но крайне неудобные для схем чисто классификационных.
В этой системе бросается в глаза значительное количество «добавлений» к типу: таких добавлений — 8 классов. Авторы «Руководства» поступили правильно: они предпочли правду внешней красивости схемы. Все эти классы изучены еще недостаточно, и пока нет бесспорных данных для включения их в тот или иной тип.
Если мы примем эту схему, как последнюю «хронологически», и будем, исходя из нее, оценивать положение вещей, то увидим следующее. Типов установилось 13, возможно, что число это чуть изменится, но, очевидно, колебания не выйдут из пределов 11–16 (уменьшение возможно за счет слияния некоторых типов червей, увеличение — за счет разбивки каких-либо из существующих типов, что, однако, маловероятно). Неясным является во всех отношениях тип «тентакулат» (Tentaculata): не выяснены его связи с первично- и вторичноротыми, не совсем ясен еще и его объем — входящие в его состав группы весьма разнохарактерны, и тип выглядит несколько искусственным. Высшие группировки данной схемы — не таксономические категории, включение их в классификационную схему излишне; детали филогенетических обоснований схемы нужно давать отдельно. Схема «Руководства» — попытка сделать «большую систему» более простой и четкой, чем это приведено в «схеме-шараде» Клауса и Гроббена.
Итак, попытки построить «большую систему», т. е. систему вроде гатчековской или клаус-гроббеновской, пока не увенчались успехом. Можно сказать больше: некоторые зоологи, придя в отчаяние от замысловатости и запутанности таких «систем», отказались от систематических схем вообще. В большом английском руководстве («The Cambridge Natural History»), выходившем под редакцией С. Хармера и А. Шиплея (S. Harmer and A. Shipley, 1896–1909), просто изложены в последовательном порядке 26 основных групп без указания их таксономической оценки.
Причины неудач «большой системы» (вполне удачными наличные системы этого рода назвать нельзя) кроются в самом методе построения. Основное и неизбежное затруднение — построение системы в виде «столбца». Какие бы категории мы ни вводили, но во всякой схеме будет некоторая, так сказать, основная категория («тип» в «Руководстве», «класс» у Клауса-Гроббена, «тип организации» у Гатчека). Эти группы должны быть размещены в естественном порядке, но в виде столбца. Конечно, при таком способе не может быть речи о полной естественности размещения таксономических категорий. Нельзя правильно расположить в двухмерном пространстве то, для чего требуется по меньшей мере пространство трехмерное.
«Столбец» категорий, конечно, не может вполне отразить действительное положение вещей. Отсюда споры о характере системы: одни различают две системы — искусственную и естественную, другие — три, прибавляя еще систему филогенетическую. Наличие таких споров показывает, что самое понятие «система», «классификация» не всегда толкуется правильно. Возможны только две системы — искусственная и естественная, так как естественная система включает в себя понятие филогенеза, и по существу филогенетическая система — синоним естественной. Защитники филогенетической системы обычно ссылаются на то, что такая система должна особенно четко отражать филогенез, должна включать в себя некоторые моменты родословного древа. Очевидно, это требование — стремление как-то исправить недочеты изобразительного порядка, связанные с двухмерностью чертежа схемы. Однако и эти «поправки» не выходят из пределов двухмерного пространства, мало того, попав в него, они неминуемо приводят к установлению надтиповых группировок, т. е. к введению ряда новых категорий, ничего схеме, как таковой, не дающих, но превращающих четкую схему в нечто запутанное. Эта путаница увеличивается произвольным применением номенклатуры таких категорий, их «ранга» и объема, самого характера. Все это естественное следствие при попытках характеризовать результат подготовившим его процессом.
Для обоснованности предлагаемой классификационной схемы (чертежа) можно дать отдельно филогенетическую схему-чертеж, но пытаться соединить то и другое вместе, значит — обрекать схему на неудачу. Мало того, — классификационная схема имеет дело с таксономическими категориями (пусть даже «надуманными» человеком), между тем «первичноротые», «разноосные» и т. п. — увы, это не таксономические категории, не систематические группы, а только моменты, характеризующие историческое развитие той или иной группы. Они нужны при характеристике категорий для выяснения их родственных связей, но совершенно неуместны в классификационной схеме, где категории приводятся в своем чисто «таксономическом» значении.
Введение моментов родословного древа — первая «организационная ошибка», допущенная при построении «большой системы». Вторая ошибка — введение в систему дихотомического принципа. Такой прием, очень удобный для составления определительной таблицы, крайне неудобен в классификационной схеме: противопоставление признаков неминуемо вызывает представления о равноценности сравниваемых групп, слишком часто далеко не таковых на самом деле. Такие «определительные» признаки обычно встречаются только в «надтиповых» группировках, т. е. снова связаны с желанием ввести в схему нечто от родословного «древа»: эта «ветвь» пошла в одну сторону, та — в другую. Но то, что выглядит очень хорошо в чертеже «древа», никуда не годится в графах схемы: длинная и короткая ветви здесь заменены названиями, изображение исторического значения и различия группировок отсутствует, а одинаковая категоризация «ветвей», конечно, ведет к грубо ошибочным оценкам их качества. Губки не равноценны остальным многоклеточным, лучистые не равноценны двустороннесимметричным, однако при способе «дихотомии» они оказываются таковыми. В схеме «древа» эти ветви были верны, в классификационной схеме они вызывают ошибочные сопоставления (ср. стр. 186).

Гриф (Альберт Великий, 1545).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК