5.3.2. Гипотезы, связывающие появление эукариот с актом симбиоза архе и протеобактерии
5.3.2. Гипотезы, связывающие появление эукариот с актом симбиоза архе и протеобактерии
В последние годы появились основания к пересмотру некоторых элементов эволюционной истории эукариот. Главным образом, изменения касаются самого происхождения эукариотической клетки и ранних этапов ее эволюции.
Преобладающим стало мнение, что клеточной линии, отделившейся от прокариот и самостоятельно трансформировавшейся в ранних эукариот, вообще не существовало. Анализ расшифрованных геномов эукариот и прокариотических организмов, как архе, так и бактерий, показал, что первичные эукариоты возникли не в результате обусловленного мутациями вертикального процесса, а как следствие горизонтального процесса – симбиоза бактерии и архе (Gupta, 1998; Martin and Muller, 1998; Ribeiro and Golding, 1998; Vellai and Vida, 1999; Margulis et al., 2000; Horiike et al., 2002, 2004; Aguilar et al., 2004; Rivera and Lake, 2004; Simonson et al., 2005; Zhaxybayeva et al., 2006). Рассматриваются существенно различные схемы такого симбиоза.
Согласно предложенной рядом авторов гипотезе (Gupta, 1998; Moreira and Lopez-Garcia, 1998; Horiike et al., 2002, 2004), эукариотическая клетка образовалась единственный раз за всю историю Земли при событии симбиоза прокариотической клетки, принадлежащей ветви архе (эоцит) и грамотрицательной бактерии (протеобактерия) (Рис. 5Б). Эти авторы полагают, что клетка архе была поглощена бактерией, образовав ядро композита, а сама бактерия сформировала окружение ядра (цитоплазму). Не исключено, что в симбиоз вступили протопласты или сферопласты, т. е. клеточные формы, лишенные оболочки (Sinkovics J.G., 2001; Wachtershauser G., 2003). Позднее хромосома бактерии переместилась в ядро, сформировав там вместе с хромосомой архе комплексный (химерный) геном эукариотической клетки. В этом геноме “информационные” гены, контролирующие репликацию, транскрипцию и трансляцию унаследованы от архе, а значительная часть “операционных” генов, кодирующих биосинтетические процессы и метаболизм эукариотической клетки, привнесены в химерный геном бактерией (Gupta, 1998; Simonson et al., 2005). В соответствии с этой моделью плазматическая мембрана (наружная оболочка) эукариотической клетки формируется из липидов, синтез которых контролируют гены бактериального происхождения.
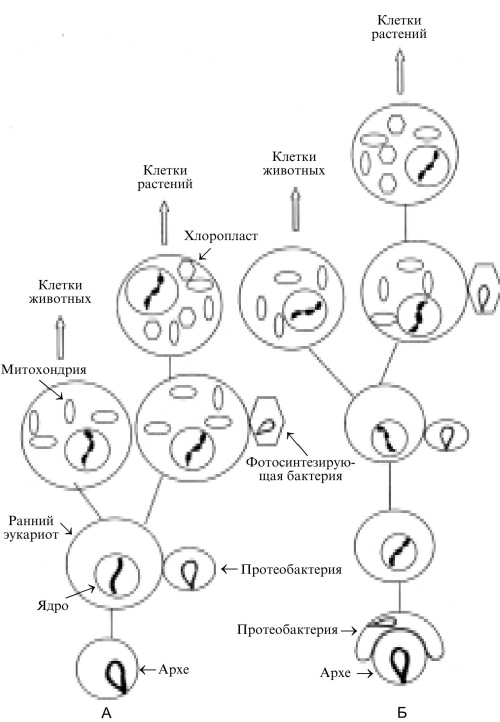
Рис. 5. Формирование современных эукариотических клеток.
А. На базе возникших эволюционным путем ранних эукариотов. Б. На базе эукариота, возникшего благодаря акту симбиоза протеобактерии и клетки архе (подробности в тексте). Рисунок создан при участии О. М. Горбенко.
Однако авторы, поддерживающие эту концепцию, расходятся в предположении о принадлежности бактерии, участвовавшей в симбиозе с архе, той или иной группе протеобактерий. Это свидетельствует о необходимости более детальных исследований. Тот факт, что какие-то гены эукариот происходят от архе, а другие – от бактерий, позволяет объяснить, почему авторы, использовавшие ранее филогенетический (мутационный) подход, вели происхождение эукариот как от архе (Woese et al., 1990; Doolittle, 1999), так и от бактерий (Hashimoto et al., 1998): все дело в том, ген какого происхождения (архе или бактериального) был выбран для исследования. Cформировавшиеся указанным образом эукариоты позже приобрели митохондрии и хлоропласты благодаря последовательным актам симбиоза с ?-протеобактерией и с фотосинтезирующими грамотрицательными бактериями, о чем уже упоминалось выше.
В рамках описанной гипотезы уместно поставить вопрос, почему слияние двух генетически различных прокариотических клеток стало стимулом для формирования клеточного ядра, т. е. образования эукариота. Интересную гипотезу предлагают Мартин и Кунин (Martin and Koonin, 2006). Эти авторы обращают внимание на то обстоятельство, что при формировании в симбионте объединенных геномов происходила фрагментация некоторых генов вследствие образования чужеродных вставок. Однако трансляция непосредственного транскрипта разобщенного гена давала “неправильный” продукт. Для получения прежнего белка следовало освободить мРНК от вставок (интронов). Необходимый для этого процесс вырезания интронов и сшивки фрагментов мРНК (экзонов), именуемый сплайсингом, должен предшествовать трансляции (образованию белков на рибосомах). Предотвратить трансляцию первичных транскриптов можно было, пространственно разделив оба процесса. Именно эту цель преследовало формирование внутренней (ядерной) оболочки. Она изолировала хромосомы от остального объема клетки (цитоплазмы), куда переместились рибосомы, происходившие от клетки архе, занимавшей центральную область симбионта. Образованные в ядре транскрипты после достаточно изощренной и, кстати, небыстрой процедуры сплайсинга уже в форме кодирующих нормальный продукт мРНК транспортируются через ядерные поры в цитоплазму, где подвергаются трансляции на располагающихся там рибосомах. На некоторые данные, вызывающие сомнения в правомерности этой гипотезы, указывают сами авторы, отмечая периодически появляющиеся публикации об осуществлении трансляции в самом ядре на присутствующих там “ядерных” рибосомах (Iborra et al., 2001). Однако утверждение о присутствии рибосом в ядре нельзя считать доказанным (см. Dahlberg and Lund, 2004). Можно высказать и другое соображение. Процессы эволюционного создания инструментов сплайсинга, формирования изолированного от цитоплазмы ядра и отделения рибосом от первичных транскриптов могли занять сотни и сотни поколений. Вместе с тем, клетки для поддержания жизнеспособности должны постоянно синтезировать белки, обладающие необходимой активностью. Это вряд ли выполнимо в случае трансляции первичных транскриптов, содержащих интроны, что неизбежно приводило бы к образованию белков с измененной структурой и нарушенными свойствами. Эту проблему, которая кажется трудноразрешимой, авторы не обсуждают. Авторы рассмотренных выше гипотез формирования эукариотической клетки из композитной клетки, образованной слиянием архе и бактерии, оставляют без ответа и ряд других вопросов. Именно это послужило причиной появления новых идей.
Значительное количество эукариотических генов не имеет аналогов ни у бактерий, ни у архе. Хартман и Федоров (Hartman and Fedorov, 2002) выявили в эукариотических организмах (дрожжах, дрозофиле и др.) более 300 белков, не имеющих “родственников” у известных прокариот. Основываясь на этом факте, авторы предположили участие в формировании эукариотической клетки еще и третьего участника, также прокариота, принадлежавшего линии, названной ими хроноцитами. Эта линия сама не сохранилась и оставила след только в сформированных с ее участием эукариотах. Имея в виду функции белков, отсутствовавших у бактерий и у архе, Хартман и Федоров предполагают, что главной особенностью хроноцитов было присутствие цитоскелета и других структурных элементов цитоплазмы (шероховатый ретикулум, тельца Гольджи), которые отличают цитоплазму всех эукариот от почти неструктурированной цитоплазмы известных прокариот. Как полагают эти авторы, ядра сформировались в результате эндосимбиотического взаимодействия бактерии и архе в хроноците. Последний, уже обладавший актиновым и тубулиновым цитоскелетом, предоставил генетическую информацию, составившую базу для формирования сложно организованной цитоплазмы эукариотической клетки.
Однако было ли действительно необходимым привнесение в композит гипотетическим хроноцитом определенных признаков, которые отсутствовали у двух очевидных симбионтов – бактерий и архе? В принципе, такой необходимости не было, так как значительное количество новых признаков (генов) сформировалось в процессе эволюционного развития эукариота, т. е. уже после образования композита. В первую очередь это относится к самому ядру. О необходимости формирования у эукариот новых механизмов (а следовательно, и генов) в связи с организацией сплайсинга говорилось выше. Вообще, клеточное ядро – весьма сложное образование. Помимо двуслойной ядерной оболочки, пронизанной регулярно распределенными белковыми структурами, образующими ядерные поры, клеточное ядро имеет сложное внутреннее строение. Его фибриллярный скелет (ядерный матрикс) образован, главным образом, ядерными белками ламинами. Детали структуры ядерного матрикса на протяжении многих лет являются предметом обсуждения (см. Георгиев и Ченцов, 1960; Berezney and Coffey, 1977; Gerace and Blobel, 1982; Мосевицкий и Новицкая, 1982; Мосевицкий, 1985; van Eekelen et al., 2002). Ядерный матрикс участвует в организации и функционировании заключенных в ядре хромосом. На нем базируются ферментные комплексы, осуществляющие пооперонную репликацию ДНК (Tubo and Berezney 1987), транскрипцию (Razin et al., 1985; Jackson, 2005), сплайсинг (Ciejek et al., 1982; Mariman et al., 1982) и другие функции (Pienta and Coffey, 1984). Следует добавить также специфические для эукариот белки, обеспечивающие компактную форму хроматина (гистоны), системы, контролирующие регулярность деления эукариотической клетки (митоз) и др. Мы видим, что объем генетической информации, которую необходимо было освоить формировавшемуся эукариоту (в форме новых генов), был очень велик. Создание цитоскелета стало также необходимым условием образования современной эукариотической клетки, так как позволяло увеличить клеточный объем, фиксировать положение ядра, перейти к активному транспорту и др. Но и эта задача могла решаться уже после акта симбиоза, инициировавшего весь процесс формирования эукариотической клетки. Следует также иметь в виду, что такой акт мог бы состояться 3 и даже 3.5 млрд лет тому назад, когда потенциальные симбионты (архе и соответствующие формы бактерий) уже существовали на Земле. С другой стороны, отпечатки клеток с ядром и другими признаками, характерными для эукариотов, обнаружены в отложениях, возраст которых не более 1.7 млрд лет. Это означает, что на эволюцию от момента симбиотического образования химерной клетки до появления эукариотической клетки было отпущено свыше одного миллиарда лет – даже больше, чем на всю предшествовавшую эволюцию на Земле. Сколько на самом деле продолжался этот процесс – неизвестно, т. к. неясно и вряд ли может быть установлено, когда состоялся инициировавший его акт симбиоза.
Идея вторичности эукариотов, т. е. образования их тем или иным путем из прокариотических клеток, является доминирующей, но не единственной. Некоторые авторы отмечают, что прокариоты более совершенны, т. к. устроены рациональнее, чем эукариоты. Согласно этой линии рассуждений, передача функции хранителя наследственного материала от РНК к ДНК сопровождалась объединением небольших хромосом в крупные. При этом оказывалось неизбежным появление в генах вставок (интронов), искажающих генетическую информацию. Соответственно, сплайсинг и формирование ядра, разделяющего процессы транскрипции и трансляции, оказались вынужденными следствиями переходного периода. Согласно этой схеме, последний общий предшественник был эукариотом, а прокариоты архе и бактерии являются сестринскими линиями, происшедшими от ранних эукариот, эволюционировавших в направлении упрощения метаболизма и структуры клетки (Brinkmann and Philippe, 1999; Poole et al., 1999). Появились ветви клеток, в которых гены были освобождены от интронов, благодаря чему отпала нужда и в сплайсинге, и в самих ядрах. Сложно организованный цитоскелет также перестал быть необходимым и сошел на нет. Осваивая новые ниши, прокариоты приобрели способность к фотосинтезу и окислительному фосфорилированию. Наконец, симбиоз обладавших усовершенствованной энергетикой прокариот с породившими их когда-то эукариотами позволил последним значительно расширить ареал существования, а главное, привел к появлению животных и растений. Однако эта не лишенная изящества схема не подтверждена полученными к настоящему времени палеонтологическими данными, которые указывают на присутствие прокариот в значительно более ранних отложениях, причем разрыв составляет не менее 1.5 млрд лет. Вместе с тем, следует отметить, что находки, сделанные в ранних отложениях, пока весьма ограничены. Обнаружение в них именно прокариот может быть объяснено их более широким, по сравнению с первичными эукариотами, распространением. Обнаружение в ранних осадочных породах отпечатков клеток, содержавших ядра, позволит обсуждать описанную гипотезу всерьез. На первичность эукариот могли также указать данные мутационного анализа генов, которые показали бы, что именно гены, контролирующие сплайсинг, формирование элементов ядерного матрикса и других чисто ядерных структур, наиболее древние. Однако, насколько известно автору, таких данных нет. Мы будем придерживаться более обоснованной сегодня, хотя и остающейся весьма схематичной, концепции, утверждающей, что первичны прокариоты, а формирование эукариот было инициировано симбиозом архе и бактерии.
Главным аргументом в пользу разделенного во времени появления ранних эукариот, а лишь затем приобретения ими митохондрий, было отсутствие митохондрий у протист, рассматриваемых как примитивные эукариоты. Однако генетический анализ выявил у протист гены бактериального происхождения, которые могли принадлежать ранее присутствовавшим митохондриям (Hasegawa and Hashimoto, 1999). К аналогичному выводу привело исследование эукариотических клеток, у которых нет аппарата Гольджи, шероховатого эндо-плазматического ретикулума и некоторых других характерных для эукариот признаков. Их отсутствие достаточно убедительно может быть объяснено не исходной примитивностью этих эукариотических клеток, а утерей определенных признаков в ходе позднейшей эволюции (Clark, 1999).
Выше, при описании акта симбиоза бактерии и архе, положившего начало формированию эукариотической клетки, была использована модель проникновения архе в бактерию, т. е. бактерия, образовав цитоплазму химерной клетки, потеряла способность к автономному размножению. Потребовался дополнительный акт симбиоза – внедрение протеобактерии в формирующийся или уже сформированный эукариот, чтобы могли образоваться митохондрии.
Мартин и Кунин (Martin and Koonin, 2006) предложили в определенном смысле зеркальную схему, согласно которой при инициировавшем формирование эукариот акте симбиоза бактерия внедрилась в архе и приспособилась к автономному существованию (см. также Vellai and Vida, 1999; Martin, 2005). Так образовались митохондрии. В процессе их формирования многие бактериальные гены объединились с хромосомой архе, приняв участие в формировании ядерного генома. Согласно этой модели, формирование митохондрий и ядерных структур происходило параллельно, и отдельный акт симбиоза для появления митохондрий не потребовался.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
ПОЯВЛЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ
ПОЯВЛЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ Шишка, или цветок беннеттита (слева), ицветок магнолии (часть органов растенийудалена)В мезозойскую эру произошли два великих события в истории жизни на Земле, которые изменили и обновили органический мир Земли: появились первые
ПОЯВЛЕНИЕ ЧЕЛОВЕКА
ПОЯВЛЕНИЕ ЧЕЛОВЕКА Человек — это звучит гордо! М.
8.1. Появление эукариот
8.1. Появление эукариот Ранее мы обсуждали вопросы развития биотического круговорота на основе прокариот. Оказалось, что главные достижения в совершенствовании обеих ветвей цикла как синтетической, так и деструктивной связаны с улучшением энергетики, повышением
§ 32. Появление конечностей
§ 32. Появление конечностей На первый взгляд происхождение конечностей наземных позвоночных не вызывает каких-либо логических проблем. Их появление вторично по отношению к миграции предков наземных позвоночных на мелководье и формированию эффективного воздушного
Появление человека
Появление человека В Европе и Азии, в том числе и в нашей стране, учёные обнаружили громадное скопление костей — целые «кладбища» животных, живших несколько миллионов лет назад. Они раскопали многочисленные кости антилоп, газелей, жирафов, гиен, тигров, обезьян и других
Глава 7 Происхождение эукариот: эндосимбиоз, удивительная история интронов и исключительная важность единичных событий в эволюции
Глава 7 Происхождение эукариот: эндосимбиоз, удивительная история интронов и исключительная важность единичных событий в эволюции Пер. А. ГрызловаОрганизмы с большими, сложными клетками известны как эукариоты, что означает «обладающие настоящим ядром». Эти организмы
1. ПОЯВЛЕНИЕ ПСИХИЧЕСКОГО
1. ПОЯВЛЕНИЕ ПСИХИЧЕСКОГО Наиболее принятой в настоящее время точкой зрения является та, что психическое – атрибут живых систем, а неживая природа, включая созданные людьми сложные устройства, психикой не обладает. Это важное отличие живых систем от неживых отмечено П.
3.2. Генетический материал эукариот
3.2. Генетический материал эукариот Генетический материал эукариот сконцентрирован в ядре и представлен хромосомами, в которых молекула ДНК образует сложный комплекс с различными белками.Каждая клетка любого организма содержит определенный набор хромосом.
6.7. Регуляция экспрессии генов у эукариот
6.7. Регуляция экспрессии генов у эукариот Система регуляции экспрессии генов у эукариот связана с особенностями функционирования эукариотического генома. Хотя и у прокариот, и у эукариот функционируют системы регуляторных белков, наличие ядра и нуклеосомная
Мегасистема эукариот
Мегасистема эукариот Молекулярно-генетические подходы полностью изменили 4-царственную мегасистему эукариот. Подтвердилось мнение о сборном характере царства протистов: у них были обнаружены самые разнообразные типы структуры и функции клеток (распределение
Появление креодонтов
Появление креодонтов Когда приоткрываются двери прошлого, мы видим устремленные на нас бессмысленные взоры чудовищ. У этих фантастических порождений прошлой жизни размеры тела намного опережали развитие их нервной организации.На страницах нашей книги уже находили
5.3.1 Концепция формирования митохондрий и хлоропластов путем симбиоза бактериальной клетки и раннего эукариота
5.3.1 Концепция формирования митохондрий и хлоропластов путем симбиоза бактериальной клетки и раннего эукариота Около 2 млрд лет тому назад на Земле создалась критическая для дальнейшего развития жизни ситуация. Фотосинтезирующие бактерии, размножившись, стали