7.2.1. Монорегиональная гипотеза формирования вида Homo sapiens исключительно в Африке и его заместительного распространения на другие континенты
7.2.1. Монорегиональная гипотеза формирования вида Homo sapiens исключительно в Африке и его заместительного распространения на другие континенты
Наиболее известной является версия, согласно которой вид Homo erectus, просуществовавший почти 2 млн лет, дал начало всем последующим Homo. По этой версии Homo sapiens (человек разумный, он же современный человек) – последнее ответвление от стволовой линии Homo erectus, появившееся, согласно палеонтологическим данным, не позже 200 тыс. лет тому назад в родовом гнезде Homo на стыке нынешних Эфиопии, Конго и Уганды (иногда говорят об Африке “под Сахарой”) (Nei and Takezaki, 1996; Stringer, 2002; Clark et al., 2003; White et al., 2003; McDougall et al., 2005; Ray et al., 2005; Bouzouggar et al., 2007). Что особенно интересно, описаны окаменелости (черепа), принадлежавшие переходной форме между архаичным типом и анатомически современным человеком (Lieberman et al., 2002; White et al., 2003). Этим опровергаются довольно частые заявления об одномоментности появления современного человека.
Существуют, по крайней мере, еще две версии родственных связей людей. В Африке и ряде областей Западной Европы были обнаружены костные останки, возраст которых 500–800 тыс. лет. По мнению некоторых исследователей, они принадлежали ближайшему общему предшественнику неандертальцев и современных людей, названному Homo heidelbergensis (Гейдельбергский человек). В свою очередь, Homo heidelbergensis произошел от Homo antecessor (человек-предшественник). Согласно этой версии (Rightmire, 1998; Stringer, 2002; Lahr and Foley, 2004) Homo erectus не был предшественником современного человека. Как “боковая” ветвь Homo erectus происходил от Homo ergaster, который дал также начало виду Homo antecessor (Рис. 8Б).
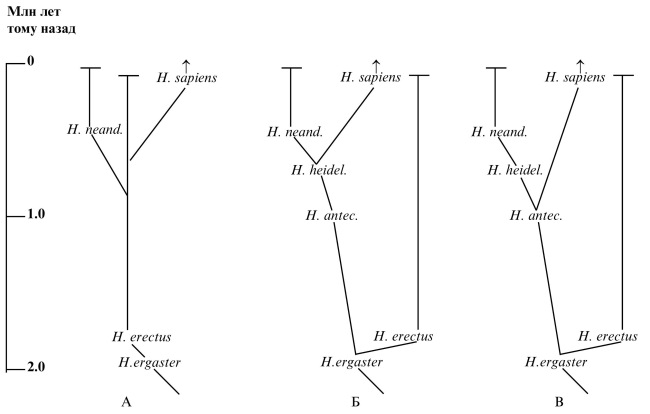
Рис. 8. Варианты происхождения современного человека (Homo sapiens).
Еще одна версия основана на результатах изучения недавних находок, сделанных в северной Испании. Их возраст по разным оценкам 650–800 тыс. лет. Костные остатки принадлежали существам, которые появились раньше гейдельбергского человека, но при этом имели больше общих черт с современным человеком и, следовательно, были ближайшими предшественниками современных людей. Этот вид также был назван предшественником (Homo antecessor). От него непосредственно происходит современный человек. Другая ветвь, исходящая от Homo antecessor – гейдельбергский человек, породила неандертальцев (Bermudez de Castro et al., 1997; Gibbons, 1997; Carbonell et al., 2005). Согласно этой версии (Рис. 8В), как и предыдущей (Рис. 8Б), Homo erectus не является предшественником современного человека, а формирует самостоятельную ветвь, исходящую, как и Homo antecessor, от Homo ergaster. Здесь уместно отметить, что многие авторы фактически отождествляют Homo ergaster и Homo erectus, полагая первого несколько более ранним вариантом второго. С учетом этого не будет ошибкой рассматривать в схемах Рис. 8Б и В Homo erectus как предшественника Homo antecessor, а следовательно, и всех последовавших линий Homo. Схемы Рис. 8Б и В допускают формирование вида Homo Neanderthalensis (неандертальцев) в Европе как ветви, исходящей от архантропов, обосновавшихся там не менее 800 тыс. лет тому назад. Очень близкие предшественники неандертальцев или они сами обитали в Европе 300–150 тыс. лет тому назад (Krause et al., 2007a). Несмотря на обнаружение предшественника Homo sapiens (по версии Рис. 8В) также в Европе, факт появления там людей современного типа около 45 тыс. лет тому назад как мигрантов из Африки в научной литературе является общепринятым (см. выше). Раздельной эволюцией линий, которые отделились от общего предшественника 800–500 тыс. лет тому назад (Green et al., 2006) и привели к неандертальцам и современным людям, можно объяснить данные, свидетельствующие о случайности их анатомических различий: сравнительное обследование черепов показало, что эти различия обусловлены скорее генетическим дрейфом, чем отбором в сходных условиях среды (Weaver et al., 2007).
Происхождение современного человека из Африки подтверждено недавними генетическими исследованиями как мужской линии (по мутациям непарной половой Y хромосомы), так и женской линии (по мутациям в ДНК митохондрий, передаваемых от матери детям с цитоплазмой яйцеклетки). Во всех случаях речь идет о нейтральных мутациях, которые практически не влияют на работу содержащих их “полиморфных” генов и, следовательно, не являются субъектами естественного отбора. Отличием “непарной” Y хромосомы и митохондриальной ДНК от парных хромосом является отсутствие генетических рекомбинаций, т. е. межхромосомных обменов участками ДНК. Благодаря этому у них легче проследить “вертикальную” составляющую, т. е. накапливание мутаций в последовательных поколениях (см. Хуснутдинова и Лимборская, 2005). При сопоставлении нуклеотидных последовательностей Y хромосом современных популяций (народов) обнаружено, что Y хромосомы всех мужчин происходят от Y хромосом, обладатели которых жили в Африке около 100 тыс. лет тому назад (Ke Y. et al., 2001; Pakendorf and Stoneking, 2005; Stringer, 2002; Ray et al., 2005). Исследования мутаций митохондриальной ДНК также привели в Африку как родину праматери всех ныне живущих людей, что с учетом первого результата вполне логично (Cann et al., 1987; Vigilant et al., 1991; Penny et al., 1995; Passarino et al., 1998; Quintana-Murci et al., 1999; Mishmar et al., 2004; Macaulay et al., 2005; Pakendorf and Stoneking, 2005; Cruciani et al., 2002, 2007; Бермишева М.А. и др., 2003; Gonder et al., 2007; Rowold et al., 2007). К выводу о происхождении современного человечества из Африки пришли разные группы исследователей, анализировавшие распределение полиморфных Alu повторов и разнообразие ряда микросателлитов у разных популяций: самые сильные расхождения со всеми остальными популяциями оказались у коренного населения Африки (Batzer et al., 1994; Jorde et al., 1997; Tishkoff et al., 2000; Watkins et al., 2001, 2003; Antunez-de-Mayolo et al., 2002; Witherspoon et al., 2006; Xing et al., 2007).
Эти же исследования позволили выделить Центрально-Восточную Африку (Африку “под Сахарой”) как регион, в котором обитали наши прародители, что полностью соответствует приведенным выше палеонтологическим данным (Gonder et al., 2007). Генетический поиск, осуществленный в этом регионе, позволил, как полагают некоторые исследователи, сузить исходный ареал до места обитания племени, поныне занимающегося собирательством и охотой. Именно здесь не менее 100 тыс. лет тому назад жили “научные” Адам и Ева. Это был, возможно, самый критический период для вида Homo sapiens, находившегося на грани вымирания после существования к тому времени уже около 50 тыс. лет. Не исключено, что сохранилось и затем размножилось потомство единственного клана. Можно предположить, что “научные” Адам и Ева передали своим детям наследуемые качества, обеспечившие им преимущества перед соплеменниками и шанс на выживание. Это могли быть более высокие умственные способности, позволившие лучше организовать охоту, отпор врагам, либо повышенный иммунитет к разразившейся тогда эпидемии и т. д.
За последующие тысячелетия люди размножились до многих тысяч особей. Около 70 000 лет тому назад, когда Homo sapiens был распространен во многих регионах Африки, группа людей двинула в сторону Аравии и покинула родной материк. Анализ мутаций, накопившихся в Y хромосоме у мужского “коренного” населения разных регионов Земли, и митохондриальной ДНК позволил проследить пути ранней миграции людей, покинувших Африку. Пока мигранты двигались вместе, новые мутации распространялись во всей популяции, а когда пути расходились, у каждой группы формировался дополнительно к уже существовавшим мутациям свой набор мутаций. Люди, оставшиеся на месте, оказались самой ранней отделившейся от всех других ветвью, и, соответственно, их потомки имеют наименьшее число общих мутаций со всем остальным населением Земли. На Рис. 9 весьма схематично показаны различия наборов мутаций в Y хромосомах, которые характерны для разных регионов Земли. Цифрой “1” обозначена “базовая” последовательность. Другие цифры обозначают мутации, характерные для народов, заселивших различные регионы. Именно эти данные позволили восстановить маршруты этой ранней и, как считают многие исследователи, единственной народообразующей эмиграции современного человека из Африки на другие континенты (Quintana-Murci et al., 1999; Mellars, 2003; Palanichamy et al., 2004; Macaulay et al., 2005; Sun et al., 2006; Fagundes et al., 2007; Underhill and Kivisild, 2007). Форсировав Красное море в его южной части и продвигаясь вдоль побережья Индийского океана, люди пришли в Индию, а оттуда на острова в южной части Тихого океана (Sun et al., 2006) и в Австралию, где обосновались около 50 тыс. лет тому назад (Bowler et al., 2003; Ingman and Gyllensten, 2003; Macaulay et al., 2005; Hudjashov G. et al., 2007). Продолжая движение на Восток, современный человек появился в Китае не позже 40 тыс. лет тому назад (Shang H. et al., 2007).
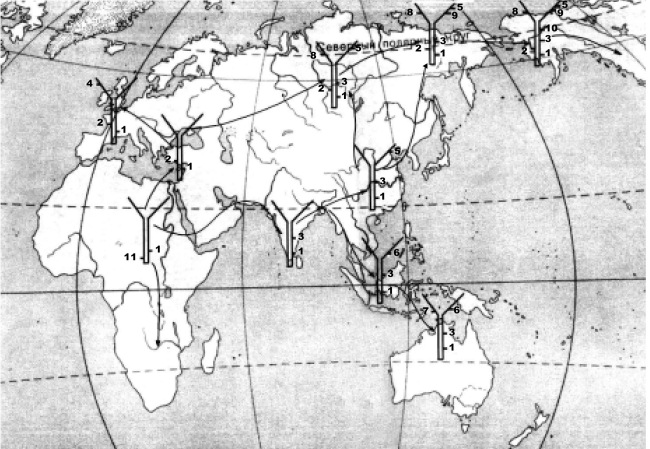
Рис. 9.
Рис. 9. Пути «народообразующей» миграции современного человека из Северо-Восточной Африки по данным мутационного анализа.
Отметки на схематических изображениях «Y» хромосомы обозначают характерные мутации, позволяющие определить очередность появления популяций и пути их миграции. Присутствие мутации «1» у всех популяций означает, что эта мутация – самая древняя. Отсутствие у Африканской популяции всех более поздних мутаций, присутствующих у других популяций, указывает на первичность именно Африканской популяции. Рисунок создан при участии Ф. В. Горбенко.
Другие группы людей направились на Север, закрепились на Ближнем Востоке, а затем, через Балканы и Кавказ стали распространяться на бескрайних просторах Евразии (Torrini et al., 2006). Около 40 тыс. лет тому назад анатомически современные люди пришли в Западную Европу, где уже более 100 тыс. лет обитали неандертальцы (Churchill, 2001; Niewoehner, 2001; Gravina et al., 2005). Несколько позже люди появились в Казахстане, на Алтае и в Южной Сибири. И они встретились с давно жившими там неандертальцами (Krause et al., 2007). Некоторые исследователи полагают, что относительно медленное продвижение людей в этих направлениях (по сравнению с их движением через Аравию в Индию и Австралию) связано именно с необходимостью преодолевать сопротивление аборигенов-неандертальцев. Это сосуществование продолжалось тысячелетиями (в Европе до 15 тыс. лет) пока неандертальцы не исчезли с лица Земли.
Несмотря на то, что неандертальцев причисляют к архаичным людям, они умели пользоваться огнем и мало уступали современным людям в искусстве изготовления орудий труда (Churchill, 2001). Они делали украшения и, по-видимому, были способны к членораздельной речи. Последнее предположение следует из того факта, что у неандертальцев был тот же вариант гена FOXP2, который присутствует у современных людей, обеспечивая членораздельную и осмысленную речь (Krause et al., 2007b; Trinkaus, 2007b). Объем мозга неандертальцев был даже несколько больше, чем у современного человека. Возможно, неандертальцы смогли бы создать свою цивилизацию. Однако 28 тыс. лет тому назад они исчезли с лица Земли (Finlayson et al., 2006). Высказывались предположения о вымирании неандертальцев из-за похолодания на Земле, в частности, на Юге Европы. Однако до того неандертальцы более сотни тысяч лет (а с ближайшими предшественниками почти миллион лет) жили в Европе и восточнее до Сибири, пережив не одно длительное похолодание (Ледниковые периоды). Поэтому к очередному похолоданию они не могли быть подготовлены хуже, чем пришедшие с Юга современные люди.
Установлено, что 40 тыс. лет тому назад в Арктической области Европейской России уже жили люди (Pavlov P. et al., 2001). Современный человек появился на Юге Европы не намного раньше. С учетом сказанного выше велика вероятность, что Европейскую Арктику осваивали неандертальцы. Похожая ситуация в Сибири. Стоянка людей, возраст которой около 30 тыс. лет, обнаружена на реке Яне далеко за Полярным кругом (Pitulko et al., 2004). Однако самое раннее проникновение современного человека на юг Сибири датируется этим же периодом. С другой стороны, неандертальцы уже обитали там более 100 тыс. лет, и логично предположить, что именно их стоянка обнаружена в Заполярной Сибири. Окончательно вопрос, кто первыми осваивал Арктику – неандертальцы или современные люди, можно будет решить после обнаружения костных останков этих первопроходцев. Однако в любом случае версия гибели неандертальцев из-за похолодания неубедительна.
Предложены две гипотезы, объясняющие исчезновение неандертальцев их контактами с людьми современного типа (кроманьольцами).
Согласно одной из них неандертальцы были вытеснены кромань-ольцами в буквальном смысле этого слова, т. е. лишены возможности добывать пищу, уничтожены в столкновениях, а возможно, просто съедены. Случаи каннибализма с обеих сторон подтверждены находками (Defleur et al., 1999; Fernаndez-Jalvo et al., 1999; Dalton, 2006). При этом, на основании исследования костных останков неандертальцев и кроманьольцев и мутационного анализа их митохондриальной ДНК сторонники указанной версии подчеркивают, что генетического смешивания не происходило (Krings et al., 1997; Ovchinnikov et al., 2000; Scholz et al., 2000; Ponce de Leon and Zollikofer, 2001; Arsuaga et al., 2002; Caramelli et al., 2003; Serre et al., 2004).
Согласно другой гипотезе, наоборот, важную роль в замещении сыграл генетический фактор: кроманьольцы смешались с неандертальцами и поглотили их. Такой вывод согласуется с данными ряда авторов, обнаруживших в геноме современных людей признаки, которые могли прийти от неандертальцев (Lahr and Foley, 1998; Plagnol and Wall, 2006; Foley and Lahr, 2007; Trinkaus, 2007a,b; Hawks et al., 2008). В этом отношении большой интерес представляет ген микроцефалин, который появился у современного человека около 37 тыс. лет тому назад и распространился во многих популяциях. Предполагается, что пришел этот ген от архаичного носителя, предположительно неандертальца (Evans et al., 2006b; Torroni et al., 2006). Проверка этой гипотезы скоро станет возможной. В настоящее время по сохранившимся фрагментам ДНК костной ткани ведется расшифровка генома неандертальцев. Уже можно сказать, что различие геномов современных людей и неандертальцев по составу нуклеотидов лежит в пределах 0.5 %, в то время как различие каждого из них с геномом шимпанзе составляет 1.2 % (Cheng Z. et al., 2005; Noonan et al., 2006; Kehrer-Sawatzki H. and Cooper D.N., 2007). В данном случае интерес представляет вопрос, будет ли среди генов неандертальцев выявлен ген микроцефалина. Если это произойдет, то гипотеза приобретения этого гена современным человеком от неандертальца станет практически доказанным фактом. Если смешивание генетических признаков было возможно, то замещение происходило, по крайней мере, частично, путем ассимиляции.
Рассмотренная здесь во многом драматическая история замещения неандертальцев современными людьми в Европе отличается от других подобных историй, происходивших в разных регионах Евразии, куда ступала нога современного человека, только большим числом известных деталей. Однако в последнее время ситуация меняется. В самых разных регионах от Китая до Северной Африки делаются все новые находки, подтверждающие вклад местных архантропов в генетику формировавшихся там рас современного человека. Обнаружены черепа, принадлежавшие людям современного типа, которые несут также определенные признаки, свойственные архантропам (Stringer, 2002). После повсеместного исчезновения архаичных людей часть этих признаков сгладилась, но другие, особенно те, которые оказались полезными, должны были сохраниться в генетическом пуле человечества, как это имеет место с геном микроцефалина, предположительно перенятым от неандертальцев (см. выше). Благодаря этим новым данным вклад архантропов в генетику современного человека стал рассматриваться как заметный (Satta and Takahata, 2002) и даже чуть ли не как определяющий (Eswaran et al., 2005).
В связи с этими новыми веяниями напрашивается вопрос, насколько правомерно считать Homo sapiens и других представителей Homo, по меньшей мере начиная с Homo erectus, разными видами, если они сохраняли способность к продуктивному скрещиванию. Логично полагать эту часть Homo единым видом, за которым можно было бы закрепить название “человек разумный” (Homo sapiens). Все его разновидности сохранили бы наименования, обозначенные на Рис. 8, и только человека современного типа следовало бы переименовать, например, в Homo sapiens sapiens, как это уже делают некоторые авторы. Такой подход кажется тем более логичным, что по некоторым оценкам формирование вида – весьма длительный процесс, требующий не менее 2 млн лет (Curnoe and Thorne, 2003), и просто не было достаточно времени для образования разных видов. Естественно, решать этот вопрос будут специалисты по генетике популяций. Имея в виду эти соображения, мы сохраним в дальнейшем за современным человеком наиболее часто используемое наименование Homo sapiens.
Естественно, возникает вопрос, как и когда современное человечество разделилось на расы, если оно первоначально образовалось в определенном регионе Африки как единая популяция. Одним из важных факторов в процессе дивергенции популяций с образованием народов и рас рассматриваются ситуации, именуемые “горлышком бутылки” (bottleneck), которые сводят популяцию к минимуму. Вследствие этого при последующем восстановлении численности популяция оказывается генетически более однородной, чем была ранее. Об одной такой ситуации, поставившей человечество на грань исчезновения, упоминалось выше. Выйдя из этого кризиса, человечество как потомство одного клана существенно повысило свою генетическую однородность. Следует полагать также, что вышедшие из Африки в район Ближнего Востока 80–60 тыс. лет тому назад люди принадлежали близким кланам, что также определяло их высокое анатомическое сходство. Дивергенция началась позже, когда люди разбились на группы, направившиеся в разные стороны, в том числе в удаленные и труднодоступные места Европы и Азии, а также в Австралию. Связи между ними если и существовали, то были сведены к минимуму. Длительной изоляции способствовали природные катаклизмы, периодически ввергавшие ту или иную популяцию в ситуацию “горлышка бутылки” (Ambrose, 1998; Bjorklund, 2003; Ramakrishnan et al., 2005). К числу таких катаклизмов относят самый мощный известный современной науке взрыв вулкана Тобо на Суматре, который произошел в период первоначального расселения людей. Выброшенный в стратосферу материал образовал сферический слой, который закрыл Солнце и вызвал сильное похолодание. Полагают, что это похолодание повлекло наступление Ледникового периода, длившегося тысячелетия (Ambrose, 1998). В условиях повышенной обособленности и значительного снижения численности из-за суровости условий вновь появлявшиеся мутации быстро охватывали всю популяцию, но из-за ее изолированности не распространялись за ее пределы. Постепенно между изолированными популяциями накапливались различия в анатомическом строении, в цвете кожи, волос.
Обособленность как одно из условий дивергенции популяций не всегда следует понимать как территориальную разобщенность. В Африке сформировались и сохраняются множество проживающих-бок о бок народностей. Достаточно упомянуть таких соседей как маленькие пигмеи и очень крупные банту. Их дивергенция началась 60 тыс. лет тому назад, и близость проживания ей не помешала. Даже в наше просвещенное время почти непреодолимым препятствием для контактов могут быть племенные, кастовые или религиозные отличия.
Процесс образования ныне существующих рас и народов не сводится исключительно к изоляции и дивергенции. Весьма существенный генетический вклад, как уже говорилось выше, могли вносить местные архантропы. Разнообразие в наборы признаков вносили встречные переселения. Так, происходила обратная миграция людей в Африку (Cоrte-Real H.B., 1996; Olivieri, et al., 2007; Gonzаlez et al., 2007). Поэтому неудивительно, что у некоторых африканцев обнаруживают черты европейцев или монголоидов. На Алтае и в Сибири встретились два людских потока – из Китая-Монголии и со стороны Кавказа (Derenko et al., 2000; Деренко и др., 2001). Признаки кавказской линии обнаруживают сейчас у американских индейцев (Brown et al., 1998).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава 1 Немного истории о том, как Homo sapiens перестал быть тунеядцем
Глава 1 Немного истории о том, как Homo sapiens перестал быть тунеядцем «На руку земледельца опирается мир», — говорит французская пословица. Но человек существует на нашей планете много сотен тысячелетий, а земледелие — всего один-два десятка. Что же служило опорой
Глава XII ЗВЕРИ И КОНТИНЕНТЫ
Глава XII ЗВЕРИ И КОНТИНЕНТЫ Наконец-то мы вступаем в привычный для нас мир — мир знакомых травяных ландшафтов с ярким солнцем и сильными ветрами. Стада пасущихся лошадей, мастодонтов и антилоп обычны для этого степного пейзажа. Палеонтологи хорошо изучили этих
Глава 2 Лес – древнейшая обитель. Как живет первозданный Homo sapiens в наше время?
Глава 2 Лес – древнейшая обитель. Как живет первозданный Homo sapiens в наше время? Почему же я избрал сегодня индейца в девственном лесу, чтобы составить себе представление о «первозданном человеке» и начальном образе жизни Homo sapiens sapiens? Вроде бы в мире – в Африке, Индонезии, на
Глава 8 Моя собственная «водяная гипотеза». «X-pithecus». «Водяные дети» Чарковского. «Грудная гипотеза» Морган и моя. Сексуальные гипотезы Морриса. Прямохождение. Краткий обзор развития икспитека.
Глава 8 Моя собственная «водяная гипотеза». «X-pithecus». «Водяные дети» Чарковского. «Грудная гипотеза» Морган и моя. Сексуальные гипотезы Морриса. Прямохождение. Краткий обзор развития икспитека. Десмонд Моррис – зоолог и директор Лондонского зоопарка. Кстати, после
Глава 16 Неандерталец и возможная причина его внезапного исчезновения. Homo sapiens и его предполагаемая родина. Всемирный потоп, его причины и последствия.
Глава 16 Неандерталец и возможная причина его внезапного исчезновения. Homo sapiens и его предполагаемая родина. Всемирный потоп, его причины и последствия. Как уже говорилось, во время ледникового периода с его оледенениями и межледниковьями трансформация наших
Триумфальное шествие Homo sapiens
Триумфальное шествие Homo sapiens Судя по археологическим и палеоантропологическим находкам, широкое распространение гомо сапиенс за пределы Африки и Ближнего Востока началось не раньше 50–60 тыс. лет назад (рис. 36). Сначала, по всей видимости, были колонизированы южные,
Белокурые, близорукие и подверженные ревматизму: еще больше оплошностей допустила матушка-природа, создавая Homo sapiens
Белокурые, близорукие и подверженные ревматизму: еще больше оплошностей допустила матушка-природа, создавая Homo sapiens Если внимательно посмотреть на цепь эволюции, то человека мы обнаружим на самой верхней ступени истории развития. Он — воплощение кроны эволюции,
У Homo erectus была походка Homo sapiens [32]
У Homo erectus была походка Homo sapiens[32] В 1978–79 гг. на берегу озера Туркана (в прошлом Рудольф) в Кении были найдены следы древних людей возрастом 1,43 млн лет (то есть на 2,2 млн лет моложе открытых Мэри Лики следов австралопитеков в Лаэтоли). В 2009 году там нашли еще три цепочки следов:
В Африке
В Африке Сам я в Африке не был, но в студенческие годы встречался и подолгу беседовал с одним из наших зоологов Владимиром Васильевичем Троицким, который в 1912–1914 годах путешествовал по Центральной Африке в районе озера Виктория-Ньянца, занимаясь сборами зоологических
7.2.2. Мультирегиональная гипотеза формирования Homo sapiens
7.2.2. Мультирегиональная гипотеза формирования Homo sapiens Хотя автору представляется предпочтительной представленная выше концепция формирования Homo sapiens в Африке и народообразующего выхода людей современного типа из Африки 80–60 тыс. лет тому назад, необходимо отметить, что
Без них мертвы континенты
Без них мертвы континенты Обитатели зеленых храмовТеперь обратим внимание на отношения между миром насекомых и растениями суши.Земля — «зеленая планета». Если взглянуть на нее со стороны, то окажется, что поверхность континентов и островов, обращенную во вселенную и
6. Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens
6. Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens Делались попытки оценить интеллект обезьян, человекообразных обезьян и гоминид до Homo sapiens, основываясь на теории Пиаже о развитии интеллекта у детей. Согласно теории Пиаже, дети проходят четыре стадии
Архаичный Homo sapiens
Архаичный Homo sapiens Первую остановку по пути к рандеву № 1 мы сделаем в позапрошлом ледниковом периоде – около 160 тыс. лет назад. Этот пункт я выбрал, чтобы осмотреть окаменелости из деревни Херто-Бури в Афарской котловине в Эфиопии. Люди из Херто интересны тем, что, по
Homo erectus и анатомически современные Homo: диета и взаимоотношения между полами
Homo erectus и анатомически современные Homo: диета и взаимоотношения между полами Примерно 1.9-1.8 млн. лет назад человек широко расселился по территории Африки и впервые покинул ее пределы. Началась колонизация Евразии. Освоение новых территорий стало возможным благодаря