5.1. Современные подходы к построению дерева жизни
5.1. Современные подходы к построению дерева жизни
До 60-х годов прошлого века степень родства между видами определяли исключительно по анатомическим, морфологическим и поведенческим признакам. Принадлежность животных, как существующих, так и вымерших, тому или иному виду, отряду и т. п. определялась по деталям строения скелета, по отдельным костям и зубам. Эти подходы сохранили значение и в последующие годы. Приобретенный опыт столь значителен, что даже плохо сохранившиеся окаменелости позволяют достаточно надежно классифицировать животных, живших десятки и сотни миллионов лет тому назад. Так, по отпечаткам раковин четко классифицируют моллюсков, существовавших 400–600 млн лет тому назад. Достаточно точное датирование окаменелостей (как животных, так и растений) позволило определить не только последовательность образования видов, но и время их образования (точки разветвления) вплоть до возраста 1 млрд лет, когда появились губки и кишечнополостные.
А

Б
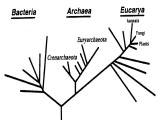
В
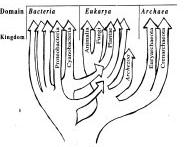
Г
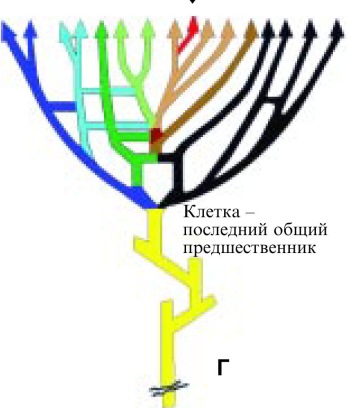
Рис. 4. Эволюция представлений о дереве жизни.
А. Дерево жизни, как его представлял Чарльз Дарвин. Б. Дерево жизни по Woese et al. (1990), ствол которого (последний общий предшественник) дает ветви бактерий и архе. В. Дерево жизни по Doolittle (1999), учитывающее горизонтальные переносы. Г. Дерево жизни, при построении которого учтены соображения о возникновении эукариотической клетки при акте симбиоза бактерии и архе.
Более древние отпечатки принадлежат исключительно одноклеточным организмам. Обнаружено значительное количество достаточно четких отпечатков прокариотических и эукариотических (ядерных) клеток возрастом до 2.5 млрд лет и 1.7 млрд лет, соответственно. Однако в более древних породах, главным образом вследствие их значительного метаморфизма, клеточные отпечатки – большая редкость. Своеобразный рекорд принадлежит Дж. Шопфу, который впервые опубликовал микрофотографии клеточных окаменелостей, обнаруженных им на Северо-Западе Австралии в осадочных породах, залегающих на глубине около 30 км (Schopf, 1993). Возраст этих пород и, соответственно, окаменелостей – около 3.5 млрд лет. Автор идентифицировал окаменелые клетки как близкие современным фотосинтезирующим грамотрицательным бактериям, которые имеют два названия: сине-зеленые водоросли и цианобактерии (Рис. 2). Главным доводом в пользу этого заключения стало присутствие в тех же отложениях слоистых известковых структур (строматолитов), поныне создаваемых колониями современных цианобактерий на мелководье. Но такой вывод равноценен утверждению, что уже 3.5 млрд лет тому назад на Земле осуществлялся процесс фотосинтеза, при котором с участием световой энергии присутствовавшие в атмосфере пары воды и СО2 использовались для органических синтезов и производства кислорода. Если придерживаться концепции зарождения жизни на самой Земле, то существование 3.5 млрд лет тому назад бактерий современного типа свидетельствует о том, что жизнь на Земле должна была присутствовать и значительно раньше (Awramik, 1992). Однако более древние не мета-морфозированные (не подвергавшиеся плавлению) породы, в которых могли сохраниться ранние клеточные формы, пока не обнаружены. Да и выводы Шопфа о биологическом происхождении сделанных им находок были оспорены как в отношении клеток (Brasier et al., 2002, 2006), так и строматолитов (Lowe D.R., 1994; Grotzinger and Knoll, 1999; Walsh, 2004). В отношении некоторых предъявленных Шопфом окаменелостей, сохранившихся совсем плохо, критика представляется обоснованной. Качество остальных отпечатков также не очень высокое. Однако сравнение с явно родственными, но более поздними и потому лучше сохранившимися отпечатками подтверждает их достоверность. В недавних исследованиях рядом авторов получены новые тому доказательства (Altermann and Kazmierczak, 2003; Schopf, 2006; Allwood et al., 2006; Hofmann and Bolhar, 2007). В частности, обнаружены следы фотосинтезирующих клеток в столь же древних отложениях на территории Южной Африки (Tice and Lowe, 2004). С другой стороны, Шопф в суммирующем обзоре (Schopf, 2006) фактически отказался от ранних утверждений, что обнаруженные им отпечатки древнейших клеток принадлежали фотосинтезирующим бактериям, похожим на современных цианобактерий. Он ссылается на существующих ныне хемотрофов, которые имеют большое сходство с теми же отпечатками. Таким образом, вопрос о времени появления фотосинтезирующих клеток вновь оказался на повестке дня.
Длительное время не удавалось обнаружить относящиеся к той же эпохе микроокаменелости, которые можно было бы с достаточной степенью уверенности отнести к хемотрофным бактериям, поныне преобладающим среди анаэробов (микроорганизмов, существующих в бескислородной среде). Эволюционно они предшествовали цианобактериям и, следовательно, не могли не быть их современниками. Наконец, в 2000-ом году было сообщено, что в Австралии в осадочных породах, формировавшихся более 3.2 млрд лет тому назад, обнаружены отпечатки нитеобразных микроорганизмов. Породы оказались богаты соединениями серы, что свидетельствовало о их формировании на океаническом дне вблизи выходов лавы и горячих серных источников. На этом основании окаменелые бактерии были отнесены к анаэробным хемотрофам (Rasmussen, 2000; Shen, 2001). Были получены и другие свидетельства присутствия более 3 млрд лет тому назад восстанавливающих CO2 до метана (метаногенных) (Ueno et al., 2006) и восстанавливающих окислы серы анаэробных микроорганизмов (Ohmoto et al., 1993). Таким образом, микроорганизмы, отличные от цианобактерий, также могли претендовать на роль последнего общего предшественника.
Проведенное выше обсуждение показывает, что ствол и ранние ветви дерева жизни могли формировать анаэробные хемотрофы, использующие наиболее примитивные пути накапливания энергии за счет преобразования неорганических соединений: восстановления двуокиси углерода и сульфата или окисления сероводорода. Фотосинтезирующие бактерии, использующие более сложные схемы метаболизма, возникли позже. Причем, по мнению ряда авторов, это были не цианобактерии, использующие в качестве восстановителя при фотосинтезе воду и освобождающие кислород, а фотосинтезирующие бактерии, которые в условиях восстановительной атмосферы могли использовать в качестве восстановителя свободный водород или сероводород (Des Maralis, 1998; Hoehler, 2005; Olson, 2006). По данным Олсона цианобактерии появились не ранее 2.9 млрд лет тому назад, а по другим данным (Kopp et al., 2005) еще позже. Такой подход позволяет объяснить сохранение весьма теплого климата на Земле в течение первого миллиарда лет развития на ней жизни сильным парниковым эффектом, который мог создаваться благодаря присутствию метана, поступавшего в атмосферу при выходах лавы и в результате жизнедеятельности метаногенных бактерий (Pavlov et al., 2000), а также присутствию двуокиси углерода (Hessler et al., 2004). Накопление и сохранение метана были возможны только при отсутствии в атмосфере молекулярного кислорода. С распространением цианобактерий и поступлением значительного количества кислорода в атмосферу содержание в ней метана было сведено, практически, к нулю. Именно с этим обстоятельством связывают не просто похолодание, но глобальное оледенение, имевшее место около 2.2 млрд лет тому назад (Kopp et al., 2005; Kasting and Ono, 2006). Мы видим, что вопрос о времени появления цианобактерий принципиален для интерпретации геохимических процессов в интервале 3.5–2.5 млрд лет тому назад и для правильного отображения дерева жизни.
Очевидно также, что данные палеонтологии не позволяют надежно датировать появление самых ранних ветвей дерева жизни и, соответственно, обоснованно судить об их очередности (см. Schopf, 1999). Успехи молекулярной биологии позволили разработать целый ряд новых подходов к решению этой проблемы. Базовую идею “молекулярных часов” высказали Цукеркандл и Полинг в 1965 г. (Zuckerkandl and Pauling, 1965). К тому времени уже была расшифрована аминокислотная последовательность ряда белков. Среди этих белков были и белки-гомологи, принадлежавшие разным видам. Оказалось, что чем дальше отстоят виды друг от друга, т. е. чем раньше произошло их разделение, тем больше различий в аминокислотных последовательностях гомологичных белков. После успехов в определении нуклеотидных последовательностей ДНК (секвенировании) появилась практическая возможность реализовать аналогичный подход непосредственно на генах, определяя происшедшие в них мутации (замены азотистых оснований, вставки, делеции, дупликации). Естественно, речь идет о полезных или практически нейтральных мутациях, которые сохраняются в потомстве и, следовательно, накапливаются с момента образования вида (Ohta T., 1987; Schuster P., 1993).
Первый вопрос, который следовало решить, возникают ли мутации в генах линейно во времени. С этой целью сопоставлялись данные о числе мутаций, накопленных у определенных видов, с палеонтологическими данными о возрасте этих видов. Такое исследование смогло охватить почти миллиард лет благодаря большому количеству палеонтологических находок, относящихся к этому периоду. Оно показало, что, по крайней мере, в пределах означенного периода линейность накопления мутаций во времени действительно имела место. Было сделано предположение, что эта закономерность выполнялась вплоть до тех давних времен, когда появились самые ранние из дошедших до нас (современных) видов. Учитывая, что речь шла о громадных промежутках времени, предстояло выбрать в клетке объекты (белки или гены), для которых характерен низкий темп изменчивости. Среди белков предпочтительными объектами являются компоненты мультимолекулярных комплексов, например белки рибосом. Структурное изменение в таком белке из-за несогласованности с другими компонентами дестабилизирует весь комплекс. Подобные мутации исключаются из популяции. По той же логике подходящими объектами при “тестировании” дерева жизни являются рибосомные и транспортные РНК. Параллельно с практическим использованием метода молекулярных часов продолжается его совершенствование (Lee, 1999; Douzery et al., 2003; M?ller and Reisz, 2005; Benton and Donoghue, 2007).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Истина одна, но подходы к ней – по множеству дорог
Истина одна, но подходы к ней – по множеству дорог Выскажем мнение, с которым, возможно, не все согласятся: гипноз, магия, религия – это все грани одного кристалла. Апологеты того или иного направления, спорящие друг с другом, напоминают четырех слепцов, описывающих слона:
ДОПОЛНЕНИЕ К ГЛАВЕ 4 Термодинамические подходы к сущности жизни. Второе начало термодинамики, энтропия и диссипативные структуры
ДОПОЛНЕНИЕ К ГЛАВЕ 4 Термодинамические подходы к сущности жизни. Второе начало термодинамики, энтропия и диссипативные структуры Нам придется начать издалека. В 1847 году Г. Гельмгольц сформулировал закон сохранения энергии (ЗСЭ). Следует помнить, что ЗСЭ является всего
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы
Глава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы Пер. А. НеизвестногоВ предыдущей главе мы обсудили возможные сценарии возникновения клеток и (будем надеяться) достигли
Зоотерапия на современном этапе. Подходы и перспективы развития…
Зоотерапия на современном этапе. Подходы и перспективы развития… Перед тем как более подробно познакомить читателя с подходом лечебной кинологии, хотелось бы несколько слов сказать непосредственно о зоотерапии, о терапии животными. В наши дни очень остро стоят
Зоотерапия на современном этапе. Подходы и перспективы развития…
Зоотерапия на современном этапе. Подходы и перспективы развития… Перед тем как более подробно познакомить читателя с подходом лечебной кинологии, хотелось бы несколько слов сказать непосредственно о зоотерапии, о терапии животными. В наши дни очень остро стоят
Из какого дерева был построен плот «Кон-Тики»?
Из какого дерева был построен плот «Кон-Тики»? Плот «Кон-Тики», на котором Тур Хейердал со спутниками в 1947 году пересек Тихий океан, был построен из бальзового (бальсового) дерева, распространенного во влажных тропических лесах Центральной и Южной Америки. Бальзовое
1.1. Современные представления о сущности жизни
1.1. Современные представления о сущности жизни Жизнь во всех ее формах и проявлениях изучает биология. Предметом биологии является многообразие вымерших и ныне существующих организмов, их строение и функции, происхождение и эволюция, размножение и развитие,
Нейробиологические подходы к проблеме сознания
Нейробиологические подходы к проблеме сознания Долгое время в науке господствовала идея наличия в головном мозге «командного центра», управляющего остальными отделами. Однако затем она была заменена концепцией «модульного мозга», согласно которой мозг имеет свои
Сладкие «слезы» дерева
Сладкие «слезы» дерева Острою секирой ранена береза, По коре сребристой покатились слезы… А. К. Толстой Чай и кофе вкусно пить с сахаром. Но где найти сахар в северном лесу? Сахарный тростник растет в тропических болотах. В жарких странах растут финики и другие сладкие
Экспериментальные подходы к изучению роли мозга в эмоциональных проявлениях
Экспериментальные подходы к изучению роли мозга в эмоциональных проявлениях Изучая функционирование лимбической системы и тех частей коры, которые связаны с эмоциями, экспериментаторы обычно используют два главных метода: 1) электрическую стимуляцию отдельных
5.4. Существовал ли последний общий предшественник, то есть был ли единый ствол у дерева жизни?
5.4. Существовал ли последний общий предшественник, то есть был ли единый ствол у дерева жизни? Прежде чем вернуться к дереву жизни, следует упомянуть высказывания, ставящие под сомнение само понятие “последний общий предшественник”. Группа авторов полагает, что
8.4.8. Терапевтические подходы, направленные на временное повышение мыслительных способностей
8.4.8. Терапевтические подходы, направленные на временное повышение мыслительных способностей Перспективным является терапевтический (медикаментозный) подход к улучшению на время памяти и других проявлений мыслительной деятельности. Уже находится в стадии испытаний
15. Современные представления о возникновении жизни
15. Современные представления о возникновении жизни Вспомните!Какие химические элементы входят в состав белков и нуклеиновых кислот?Что такое биологические полимеры?Какие организмы называют автотрофами; гетеротрофами?Теория биохимической эволюции. Наибольшее
Калабасы из дерева
Калабасы из дерева Как уже было сказано, калабаса может быть изготовлена не только из тыквы. Упоминалось, что она может быть деревянной. Но не каждое дерево подойдет для того, чтобы стать сосудом для мате. Так какие же возможны варианты?Palo Santo«Священное дерево». В данном