Какие генно-инженерные приемы подсмотрены в природе
Какие генно-инженерные приемы подсмотрены в природе
Благодаря перемещающимся элементам, генофонды всех организмов потенциально составляют общий генофонд всего живого. Реализация этой потенции, т.е. передача генов между разными таксонами, детально документирована. У млекопитающих и птиц практически идентичные провирусные последовательности появились уже после их эволюционного обособления. Поток генов между далекими организмами является реальностью. Вопрос заключается только в том, насколько часто организмы присваивали чужеродные гены в качестве «благоприобретенных собственных функциональных генов».
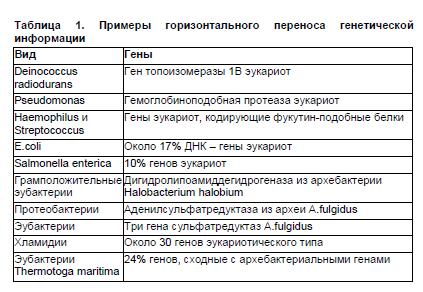
Наглядным примером горизонтального обмена между прокариотами и высшими эукариотами являются результаты исследований представленности в прокариотических геномах нуклеотидных последовательностей, типичных для эукариот (табл.1). Из этой таблицы следует, в частности, что в геноме симбионта человека, кишечной палочки, примерно 17% ДНК имеет эукариотическое происхождение. Хорошим примером естественной генетической трансформации является агробактериальная трансформация растительных клеток. Отличительная черта бактерий рода Agrobacterium (A.tumefaciens, A.rhizogenes) — способность вызывать развитие так называемых корончатых галлов (своего рода опухолей) у большого круга двудольных растений. При этом происходит перенос фрагмента ДНК агробактерии в геном растительных клеток. Такая клетка со встроенным участком агробактериальной ДНК продуцирует ряд органических веществ, служащих для агробактерии специфическими источниками углерода и азота. Такой перенос — уникальный природный процесс обмена генетической информацией между бактерией и растением, и именно его ученые взяли на вооружение для получения трансгенных растений, встраивая целевой ген в участок агробактериальной ДНК, переносимый в растение.
Эффективность трансформации растительных клеток может быть увеличена за счет использования штаммов A.tumefaciens, обладающих повышенной вирулентностью по отношению к данному виду растений. Тем не менее, частота трансформации весьма низка — только одна из 10 тысяч растительных клеток становится носителем рекомбинантной ДНК.
Возможно около 20 способов проникновения и межвидовой миграции генетических элементов, в их числе трансформация, трансдукция, перемещение транспозонов, плазмид, вирусов, неполовой обмен хромосомами и образование симбиотических ассоциаций. Информационная емкость переноса информации, выраженная в генах, варьирует от единиц до сотен и тысяч в случае плазмид и симбионтов.
Под влиянием стресса резко увеличиваются частоты горизонтального обмена материалом наследственности между бактериями, у растений — частоты перекрестного опыления у самоопылителей. В последние годы в различных моделях стресса у многих видов высших организмов наблюдают увеличение частот рекомбинационных событий, транспозиций, различных мутационных событий. Отчетливые данные о связи дестабилизации генетического материала с действием стрессирующих факторов, полученнные Б. МакКлинток, впоследствии привели к развитию представлений о системах «природной генетической инженерии» (Shapiro. 1992,1995).
Изначально термин «генетическая инженерия» применяли для обозначения целенаправленной манипуляции наследственными детерминантами с целью изменения существующих видов. В настоящее время этим термином обычно обозначают генетические манипуляции, с помощью которых формируется организм, имеющий новую комбинацию наследуемых признаков. Иначе ДНК-технологии можно определить как отрасль биологии, которая изучает явления и конструирование наследственности и изменчивости. Современный этап ДНК-технологий неразрывно связан с необходимостью увеличения источников благосостояния и здоровья человечества. ДНК-технологии стремительно увеличивают наши знания в одной из наименее исследованных областей — наследственности и законов ее изменения естественным и экспериментальным путем.
Свыкшись с материальностью генов, человек, естественно, тут же захотел заняться генной хирургией. Для этого в природе имеются ферменты рестриктазы, с высокой точностью разрезающие молекулу ДНК в определенных сайтах (сочетаниях нуклеотидов), и ферменты лигазы, «сшивающие» такие разрывы. Именно эти ферменты послужили основой для создания строго запланированных генных конструкций.
Использование рекомбинантных (перестроенных) ДНК различного происхождения составляет основу ДНК-технологий. Теоретически все 30-40 тысяч структурных генов человека и животных доступны теперь экспериментальному анализу. Поэтому желательна идентификация всех генов; составление карты тканеспецифичности их экспрессии; идентификация регуляторных областей генов; построение глобальной регуляторной карты генома; классификации генов по структурным и биохимическим функциям их продуктов; идентификация всех потенциальных белков и доменов; анализ распределения полиморфизма и мутаций; определение эволюционных и популяционных взаимосвязей; создание коллекции генетического материала и тд.
Устойчивость нити ДНК в составе хромосом регулируется целой системой ферментов, контролирующих три матричных процесса — репликацию, транскрипцию и трансляцию, и три собственно генетических процесса — репарацию, рекомбинацию и сегрегацию нитей ДНК и хромосом. Белковые продукты «генов метаболизма ДНК» образуют комплексы, которые следят за устойчивостью нитей ДНК, надежностью их репликации и рекомбинации, корректируют однонитевые и двунитевые повреждения. Степень активности этих комплексов весьма чувствительна к физиологическому статусу клетки. Ю.Я. Керкис (1940) впервые показал, что спонтанные наследственные изменения возникают за счет нарушения внутриклеточного метаболизма и физиологического гомеостаза. Устойчивость ДНК и темп мутаций могут в случае клеточного стресса меняться в десятки и сотни раз.
Началом эры генной инженерии растений принято считать 1973 год, когда впервые был проведен целенаправленный перенос гена. Фактически генная инженерия продолжает направление традиционной селекции сельскохозяйственных культур, однако достигает поставленных целей намного быстрее. Основные отличия генетической инженерии от традиционной селекции заключаются в том, что улучшение свойств культурных растений достигается либо улучшением существующей, либо созданием новой генетической вариации. При использовании традиционных методов скрещивания гарантия получения искомой комбинации генов, то есть желаемого признака у растения, практически отсутствует.
Прогресс современной науки во многом определяется и в решающей степени зависит от экспериментальной и практической реализации новых идей и подходов в клеточной и молекулярной биологии. Химерные и трансгенные животные и растения — это наиболее яркое подтверждение потенциальных возможностей фундаментальной и прикладной науки. Такие организмы стали основными инструментами в исследованиях функций генов, процессов дифференцировки, эмбрионального развития, клеточной гибели и старения. Несомненный прорыв в деле создания химерных и трансгенных организмов связан с разработкой ЭСК-технологий и микрохирургической техники работы на изолированных зародышах. В этих технологиях эмбриональные стволовые клетки стали связующим звеном между системами in vitro и in vivo, что дало возможность легко переносить результаты исследования с клеточного уровня на уровень целого организма. При этом значительно повысилась эффективность метода трансгеноза — до 40-50% по сравнению с 1% при использовании техники инъекции чужеродной ДНК (генов) в пронуклеусы зародыша на стадии зиготы.
ДНК-технологии позволяют исследовать и направленно изменять материал наследственности на разных уровнях его организации — генном, хромосомном, геномном, популяционно-генетическом. Интересно, что в смысле управления наследственностью «генетическую инженерию» использовали в течение тысячелетий безымянные селекционеры, благодаря которым еще в эпоху неолита и было введено в культуру абсолютное большинство возделываемых в настоящее время видов растений.
Переходя непосредственно к описанию методов генетической трансформации, отметим, что на сегодняшний день молекулярная генетика располагает значительным набором знаний и приемов для осуществления переноса генов из одних организмов в другие. Технология создания трансгенных растений включает большое количество этапов, среди которых можно выделить: получение целевых генов, создание векторов; трансформацию растительных клеток; подтверждение трансформации молекулярно-биологическими методами — обнаружение функционирующего целевого гена; регенерация целого растения из трансформированных клеток.
Подготовительный этап: конструирование вектора. На первом этапе конструирования рекомбинантной ДНК готовят вставки, пригодные для последующего соединения с вектором. В настоящее время наиболее часто используются 3 метода их получения: из фрагментов геномной ДНК; путем ферментативного или химического синтеза фрагментов ДНК; из сегментов ДНК, полученных с помощью ферментативного копирования РНК-матрицы in vitro.
В качестве вектора, которым может быть любой небольшой внехромосомный элемент (плазмида, ДНК фага или вируса), для трансформации растительных клеток обычно используют бактериальные плазмиды.
Следует отметить, что в большинстве случаев целевой ген подвергается модификации, поскольку, несмотря на универсальность генетического кода (он одинаков для всех организмов вне зависимости от уровня их организации), состав триплетов, кодирующих одни и те же аминокислоты у организмов, принадлежащих к разным видам, имеет некоторые отличия.
Замена кодонов никоим образом не сказывается на первичной структуре белка, в то время как экспрессия гена может быть усилена в сотни раз. Необходимый уровень экспрессии целевого гена в клетках растения достигается посредством использования соответствующих регуляторных элементов, контролирующих работу гена, — промоторов и терминаторов.
Следует отметить, что среди известных в настоящее время промоторов один из самых сильных — промотор 35S вируса мозаики цветной капусты, поэтому в большинстве случаев именно его используют в качестве регулятора экспрессии целевого гена.
Таким образом, вносимая генетическая конструкция (вставка или кассета экспрессии) — это группа функционально связанных участков ДНК, состоящая из высокоактивного промотора, непосредственно за которым располагаются соответствующий целевой ген и терминатор транскрипции. После получения вектора и вставки начинается процесс конструирования рекомбинантной ДНК. Полученные молекулы ДНК вводят в бактериальные клетки для клонирования, что приводит к накоплению рекомбинантной ДНК. Эффективное увеличение количества ее копий возможно лишь при обеспечении оптимальных условий существования вектора, использующего метаболиты, ферменты и другие белки клетки-хозяина, а также ее аппарат белкового синтеза, поэтому основной инструмент молекулярного клонирования — совместимая комбинация хозяина и вектора. Наиболее широко применяются такие сочетания, когда в роли хозяина выступает штамм E.coli, а в роли вектора — его плазмида. Проникновение вектора в живые клетки E.coli проходит наиболее эффективно при условии повышенной проницаемости клеточных мембран, обусловленной, например, их локальным разрушением. Нарушение их целостности достигается либо воздействием электрического тока — электропорацией, либо посредством обработки клеток определенными химическими веществами, после чего перенос вектора происходит в течение нескольких минут. Векторы обычно содержат маркерные гены, благодаря которым осуществляется отбор клеток с измененным генотипом. Например, клетки, чувствительные к определенному антибиотику или токсину, можно использовать в комбинации с векторами, содержащими гены устойчивости к этим агентам. Выращивая микроорганизмы в условиях, при которых проявляется зависимость от маркерных генов, можно отобрать и размножить клетки, несущие требуемый генетический материал.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Равновесие в природе
Равновесие в природе Эрозию почвы и потерю плодородия можно рассматривать как результат нарушения природного равновесия. Леса и луга могут оставаться неизменными на протяжении очень длительного периода; чтобы изменить течение рек, требуются миллионы лет. Число
Способы вызывания гипноза: физиологические и психические приемы
Способы вызывания гипноза: физиологические и психические приемы Обращаясь к вопросу о вызывании гипноза, необходимо иметь в виду, что для этого с пользою могут служить различные способы, из которых одни могут считаться физиологическими, так как они действуют
Дозы, потенции и повторные приемы средства
Дозы, потенции и повторные приемы средства ДозыВопрос дозирования гомеопатических лекарств представляет собой камень преткновения для понимания гомеопатии. В обычном понимании доза лекарства — это определенное количество таблеток, капель, шариков или миллилитров
Глава 12. НЕЧИСТОПЛОТНЫЕ ПРИЕМЫ ВОЗДЕЙСТВИЯ НА СОПЕРНИКА
Глава 12. НЕЧИСТОПЛОТНЫЕ ПРИЕМЫ ВОЗДЕЙСТВИЯ НА СОПЕРНИКА Некоторые из описанных в предыдущих главах приемов стимуляции организма могут быть названы допинговыми. Применение их для достижения успеха в конкуренции с другими собаководами за высокий выставочный ранг при
1. О природе М-полей
1. О природе М-полей Итак, согласно рассматриваемой гипотезе, фактор, определяющий образование форм на всех уровнях организации материи, — это морфогенетические поля. Утверждается, что это поля нового типа, не известного физике; соответственно, они не могут быть измерены
«Самая прекрасная в природе еда…»
«Самая прекрасная в природе еда…» Первым в массовую продажу было выпущено молоко, содержащее рекомбинантный бычий гормон роста, известный как rBGH. Это была генетическая манипуляция, запатентованная «Монсанто». Старательно придерживаясь доктрины о «существенной
Любовь к природе
Любовь к природе Первобытный собиратель, охотник, садовод были естественными членами экологических систем. Казалось бы, их влияние на природу не было разрушительным и они не нуждались в запретах поведения, нарушающего окружающую среду. Более того, обладай они сильными
Как это бывает в природе
Как это бывает в природе Возрастая численно, вид как бы усиливает свое давление на свою среду обитания, экосистему и биосферу. В ответ среда обитания, включающая в себя уйму других видов, в том числе пищевые объекты, конкурентов и потребителей того, о ком идет речь,
Глава II Животные в природе
Глава II Животные в природе
Почтение к природе
Почтение к природе Существуют достаточно разумные причины уважать естественный порядок вещей и не считать, будто люди способны так легко улучшить его путем непродуманного вмешательства. Это оказалось верным по отношению к окружающей среде: каждая экосистема есть
О природе и воспитании
О природе и воспитании Идея о том, что нас создает наше прошлое — главное прозрение Чарльза Дарвина. Он первым понял, что можно уйти от мысли о божественном творении видов, не отказываясь от идеи «творения» вообще. Каждое живое существо «сотворено», «сконструировано» без
РАВНОВЕСИЕ В ПРИРОДЕ
РАВНОВЕСИЕ В ПРИРОДЕ Понятие «равновесие в природе» стало одним из центральных положений экологической мысли, начиная с древних времен. Хотя это словосочетание несколько неопределенно (равновесие чего именно?), оно предполагает, что природа, если ее не тревожить, всегда
4. Методы и приемы дрессировки служебных собак
4. Методы и приемы дрессировки служебных собак Дрессировка служебных собак слагается из:1) общей дрессировки, 2) специальной дрессировки и 3) тренировки.Цель общей дрессировки — выработать у собаки навыки общего послушания и дисциплины, необходимые для управления ее
Методы и приемы общей дрессировки
Методы и приемы общей дрессировки Прием: хождение рядом. Цель приема — выработать у собаки стойкий навык безотказного движения рядом с дрессировщиком по команде и жесту (рис. 105), Условными раздражителями данного приема будут команда «рядом» и жест — легкий удар ладонью
Методы и приемы специальной дрессировки собак
Методы и приемы специальной дрессировки собак Дрессировку производят с тем, чтобы выработать у собаки специальные навыки, необходимые при использовании ее в том или ином виде службы (розыскной, сторожевой, пастушьей и др.).К специальной дрессировке собак приступают