Глава 6. Жизнь в карикатуре
Глава 6. Жизнь в карикатуре
Несколько полушутливых строчек, предваряющих каждую главу нашего повествования, уже стали, как нам кажется, своеобразной традицией: хорошей или дурной — судить читателю. Но, честное слово, готовясь к рассказу о функциональной роли белковых молекул в основных жизненных процессах, поневоле сбиваешься на высокоторжественный и даже патетический слог, более подходящий для выступления на юбилейных торжествах, чем для «оживления» сухого, в сущности, околонаучного трактата.
Так что на этот раз придется обойтись без привычного зубоскальства, неуместного в столь ответственный момент перехода к святая святых естествознания — жизни.
Белок — основа жизни
В другой ситуации такой заголовок показался бы слишком выспренным, но читатель, предупрежденный о недопустимости всяких ухмылок, наверняка поймет нас правильно. И это несмотря на то, что пока мы рассмотрели только один аспект деятельности белковых молекул в организме — ферментативный катализ.
Называя именно этот аспект наиболее важным, мы, вне сомнения, понимаем всю условность такого утверждения. Зачем же противопоставлять ферменты, скажем, сократительным белкам, от которых зависит подвижность наших мышц? Нет, конечно, умолчание о многих функциях белков в организме диктуется вовсе не «второстепенностью» этих функций, а исключительно особенностями структуры настоящей книги. И все же мы должны хотя бы вкратце остановиться на роли белков в процессах самосборки надмолекулярных структур клетки.
Простейший пример «самособирающегося» надмолекулярного комплекса — четвертичная структура белка — уже был нами вскользь описан. Но благодаря белкам специфической пространственной структуры оказывается возможным самопроизвольное формирование многих, гораздо более сложных структур — элементов клетки. При этом в структуры такого типа наряду с молекулами белков вовлекаются также и соединения иной природы.

Так, белки в сочетании с РНК особого сорта образуют надмолекулярные частицы — рибосомы, играющие первостепенную роль в процессе биосинтеза белковых молекул на матричной РНК. Другие белки совместно с соединениями жировой природы, липидами, самопроизвольно формируют на поверхности клеток мембраны — тончайшие образования, регулирующие материальный обмен клетки с наружной средой.
И все эти и другие сложнейшие структуры целиком определяются и удерживаются невалентными межмолекулярными взаимодействиями, причем образуются они, повторяем, самопроизвольно. Этот факт можно объяснить только способностью белковых молекул принимать в растворе строго определенную пространственную форму, что, как видим, оказывается определяющим фактором также и для процесса самосборки.
С большим трудом удерживаются авторы от триумфального восклицания: «А мы что говорили?!» И в самом деле, слова о том, что именно в пространственном строении молекул белков следует искать объяснение почти всех их удивительных свойств, оказались без малого пророческими. Читатель имел возможность убедиться в их правоте: избирательность ферментов, их стереоспецифичность, быстродействие в качестве катализаторов, способность упорядочить запутаннейшие цепи химических реакций в организме, участие молекул белков в самосборке надмолекулярных структур — все это прочно связано с представлением о третичной структуре белков — соединений, удивительнейших во всех отношениях.
И в самом деле, даже если ограничиться лишь двумя функциями белка — ферментативной и структурообразовательной, — поражаешься разнообразию возможностей белков. Подумать только: комбинируя в линейной последовательности двадцать аминокислотных остатков, природе удается «получить» ферменты, контролирующие тысячи химических реакций самой разнообразной природы или блоки надмолекулярных структур (подчас очень причудливых), число которых и вовсе уж не поддается оценке.
Именно белки являются тем удивительно пластичным инструментом и материалом одновременно, обеспечивающим существование и функционирование всех живых структур любого организма. Ведь даже когда основная масса организма образована веществами небелковой природы, как, например, у большинства растений, собственно жизненные процессы протекают лишь в тех структурах, где содержится много белка. Ибо в древесине, пробке, оболочке взрослой растительной клетки практически никаких процессов обмена веществ не происходит, это мертвые образования, выполняющие чисто механические функции. А функциональной основой жизни, обусловливающей всякое активное ее проявление, всегда оказываются белки.
И однако, будем справедливыми. В одной из важнейших функций всякого живого организма белковым молекулам верно и очень изобретательно ассистируют также и молекулы другого важнейшего класса биополимеров. Ибо, как справедливо отметил один известный биохимик, белки могут все, кроме одного: они не могут копировать самих себя.
Назад к нуклеиновым кислотам
Наше возвращение к нуклеиновым кислотам вызвано вовсе не тем, что авторские пристрастия вдруг изменились и белки перестали ходить у нас в любимчиках. Просто для дальнейшего прославления роли белков в жизненных процессах нам необходимо познакомиться со схемой их синтеза, и здесь никак не обойтись без деталей некоторых молекулярных механизмов, где первостепенную роль играют молекулы РНК и ДНК. В первой главе об этих процессах — самокопировании молекул ДНК, размножении РНК-овых копий, синтезе белка — уже шла речь, но тогда мы вынуждены были обойтись чисто формальным, символическим их описанием (пусть даже и хореографическим). Теперь же, вооруженные запасом необходимых сведений о молекулярных структурах и взаимодействиях, мы готовы заново рассмотреть и молекулы нуклеиновых кислот, и процессы их воспроизведения.
Итак, прежде всего о структуре комплементарных парных комплексов молекул ДНК, о которых шла речь в начале книги. Комплементарные пары оснований, напоминаем, образуют аденин с цитозином и гуанин с тимином. Аденин и гуанин относятся к числу так называемых пуриновых оснований; эти основания представляют собой два сочлененных цикла — шестичленный и пятичленный, образующих одну плоскость. Два других основания, цитозин и тимин — пиримидиновые, содержат только шестичленный цикл. Таким образом, схема соединения оснований такова, что большое основание образует комплементарную пару с малым, но никогда — большое с большим или малое с малым. На вопрос же о том, почему большой гуанин объединяется именно с малым цитозином, но не с малым тимином, также ответить сравнительно легко. При сближении определенным образом ориентированных ароматических колец гуанина и цитозина между ними возникают три водородные связи, причем группы, участвующие в их образовании, хорошо соответствуют друг другу. При сближении аденина и тимина также появляются водородные связи, но только две. А вот между аденином и цитозином водородные связи либо не образуются вовсе, либо, если и возникают, то очень слабые.
С учетом этих подробностей становится ясной природа сил, удерживающих комплементарные нити ДНК вместе: пара комплементарных оснований образует общую плоскость из двух колец, стянутых водородными связями. При этом такие вот плоские элементы, складываясь друг с другом, образуют как бы стопку правильной формы. Эта стопка удерживается невалентными силами, а вдоль нее, закручиваясь в форме спирали, тянутся две нити регулярной, повторяющейся части молекулы — сахаро-фосфатный остов. Поэтому такая структура и называется двойной спиралью ДНК.
Впрочем, кто же в наши-то дни этого не знает! Знаменитая двойная спираль. В некотором роде символический знак новой биологии. На фасаде главного корпуса Академии наук БССР, возведенном несколько десятилетий назад, есть барельефы с изображением символов науки, имевших хождение в те годы, — глобуса, реторты и электрофорной машины. Так вот, если в ближайшее время президиум академии решит заменить их чем-то более созвучным эпохе, то, по нашему мнению, скорее всего это будут спутник, стилизованный атом лития (три электрона), и, конечно, двойная спираль — символ чего-то биологического.
Здесь надо отметить, что очень широко распространено даже среди части биологов совершенно ошибочное представление, будто образовывать спиральные структуры могут только биологические полимеры и что именно в этом заключен сам таинственный смысл их «биологичности». Однако спираль — наиболее естественное состояние почти всякого полимера при невысоких температурах; в кристаллической форме до 90 процентов полимерных молекул свернуты в спираль, в растворах также могут спирализовываться значительные участки зауряднейших, хорошо знакомых нам из повседневного быта полимеров — полиэтилена, полихлорвинила, нейлона и т. д. Так что способность молекул белков и ДНК к образованию спиральной структуры не является каким-то загадочным свойством, выделяющим их в ряду прочих полимеров. Но существование именно такой пространственной структуры спирали ДНК — факт в высшей степени замечательный, и его открытие Дж. Уотсоном и Ф. Криком в 1953 году по праву считается одним из главных событий биологии XX века.
Собственно говоря, «открытие» не вполне подходящее слово. Дж. Уотсон и Ф. Крик предсказали эту структуру, исходя из самых общих положений кристаллографии и рентгенограмм, истолковать которые можно было очень и очень по-разному. Их работа не только положила начало триумфальному (как, по крайней мере, кажется со стороны) шествию молекулярной биологии — она утвердила право биологов на теоретические исследования, считавшиеся до тех пор почти неприличными. Сам великий Э. Чаргафф — звезда первой величины в области исследования нуклеиновых кислот — отнесся к намерению Дж. Уотсона и Ф. Крика расшифровать структуру ДНК чисто умозрительными методами с великолепным ироническим презрением. Буквально накануне публикации Дж. Уотсоном и Ф. Криком их эпохальной статьи он справлялся в письме к руководителю лаборатории Дж. Кендрью, чем там занимаются его клоуны от науки. Такова была участь биолога-теоретика в недавнем прошлом (к сожалению, также и значительное время спустя — авторам известны многочисленные примеры).
Уже сама структура двойной спирали подсказывает способ реализации процесса репликации. Ведь если основания обладают столь четко выраженным попарным сродством, значит, у одиночной нити ДНК каждый свободный нуклеотид будет «стараться» занять место напротив своего партнера. А после этого стоит только последовательно соединить их, выстроенных таким образом, в одну нить. Так оно примерно и происходит на самом деле. В процессе репликации двойная спираль раскручивается, и на каждой отделившейся ее нити «нарастает» новая комплементарная нить.
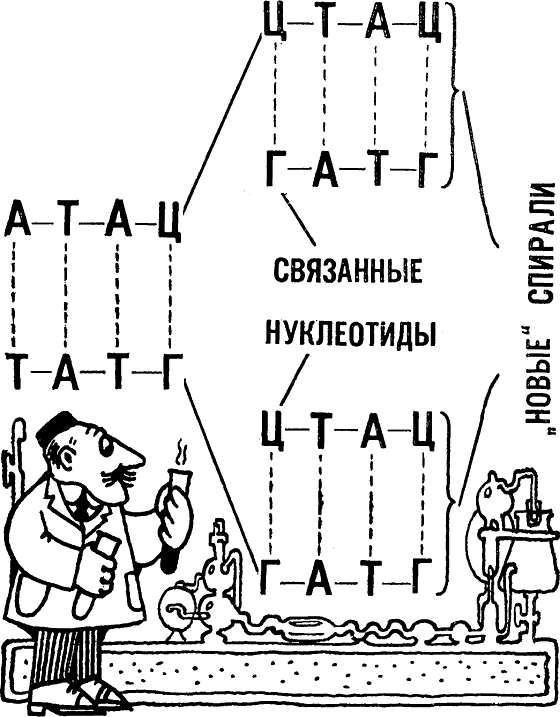
Присоединившиеся нуклеотиды «сшиваются» с остальной частью наращиваемой нити при помощи специального фермента. Вот и все. Раскручивание старой и наращивание новых спиралей продолжается до тех пор, пока спираль не раскрутится полностью, а каждая из ее нитей образует со свежесинтезированной комплементарной копией новую двойную спираль.
Схема процесса транскрипции — синтеза молекул РНК на матрице ДНК — в принципе сходна с репликацией; это и неудивительно, если вспомнить значительную структурную близость молекул ДНК и РНК. Наиболее существенное отличие заключается в том, что комплементарная последовательность РНК копирует не всю молекулу ДНК, а только ее фрагменты; важно также, что в качестве матрицы для синтеза РНК используется только одна из нитей, образующих двойную спираль, но не ее комплементарная копия. В искусственных условиях удается получить смешанную двойную спираль, образованную молекулами РНК и ДНК; такие спирали характерны для некоторых вирусов.
Как синтезируются белки
И процесс репликации, и родственный ему процесс транскрипции способны вызвать немало удивления совершенством своей организации даже в столь бледном и лишенном подробностей описании. Однако по сравнению с молекулярными механизмами биосинтеза белка они выглядят сравнительно простыми и почти очевидными.
В процессе синтеза молекул нуклеиновых кислот расположение нуклеотидов в определенном порядке происходит как бы само по себе, за счет сродства каждого из нуклеотидов к своему комплементарному партнеру. Первоначально биологи предполагали, что и при синтезе белка на матричной молекуле РНК происходит нечто подобное: каждая аминокислота стремится «примкнуть» к кодирующему ее триплету, а последовательное соединение аминокислот осуществляется особым ферментом.
Начались поиски принципов структурного соответствия между аминокислотами и кодирующими их триплетами, более того, кое-кому такие принципы удалось даже установить. Многие значительные открытия побуждают исследователей действовать по аналогии и по этой самой аналогии открывать несуществующие вещи. Это относится, конечно, не только к биологам. Скажем, очень часто в качестве примера блестящего успеха теоретической научной мысли приводят открытие французским астрономом У. Леверье планеты Нептун — знаменитое «открытие на кончике пера».
В этой связи редко вспоминаются факты менее известные: когда У. Леверье опубликовал свою работу и астрономы действительно нашли в указанном им месте неизвестную ранее планету, как из рога изобилия посыпались сообщения об открытии «по методу Леверье» еще примерно полутора десятков абсолютно несуществующих планет. Некоторые из них исходили — увы! — от самого У. Леверье, до конца своей жизни не отказавшегося от попыток обнаружить на орбите, находящейся внутри орбиты Меркурия, планету Вулкан, существование которой им было предсказано.
Словом, процесс «считывания» нуклеотидной последовательности молекулы РНК, приводящий к образованию белковой молекулы с соответствующей аминокислотной последовательностью, оказался организованным совершенно иначе и намного сложнее, чем процессы копирования молекул нуклеиновых кислот.
Взять хотя бы то, что отнюдь не все типы молекул РНК, находящиеся в клетке, несут информацию о первичной структуре каких-нибудь белковых молекул. Молекулы РНК с такой функцией называют информационной РНК. Есть, однако, и иного типа молекулы РНК, играющие первостепенную роль как раз в процессе сопоставления каждому триплету информационной РНК соответствующей ему аминокислоты. Этот тип молекул РНК носит название транспортной РНК.
Такое название, на наш взгляд, не вполне точно отражает функцию этих молекул. Может сложиться впечатление, что это молекулы-«подвозчики», в то время как никакой специальной деятельности по перемещению аминокислот к информационной РНК они не выполняют. Правильнее, по-видимому, рассматривать их как своеобразные молекулы-«переводчики». Судите сами. В клетке есть двадцать типов молекул транспортной РНК — по числу аминокислот, используемых в белковом синтезе. Каждая из аминокислот может образовать некоторое промежуточное соединение со «своей» транспортной РНК. При этом само «узнавание» аминокислотой подходящей молекулы транспортной РНК не происходит непосредственно: и в этом случае нет никаких сил специфического сродства двух молекул, которые определяли бы подобное связывание. Эту функцию выполняет специальный фермент — аминоацилсинтетаза. Ферментов этого типа в клетке также двадцать.
Мы уже много говорили о специфичности ферментов, так что первый этап акта связывания аминокислоты с молекулой подходящей транспортной РНК — образование промежуточного комплекса фермента со «своей» аминокислотой — вполне понятен. Однако аминоацилсинтетаза обладает еще одним типом специфичности: она способна также «узнавать» и соответствующую данной аминокислоте транспортную РНК. «Захватив» молекулу аминокислоты, фермент взаимодействует с определенным участком молекулы транспортной РНК и присоединяет к ней молекулу аминокислоты. Обратите внимание, что на этом этапе единственным элементом, наделенным способностью к «узнаванию», является именно белок — фермент аминоацилсинтетаза, обладающий избирательным сродством, с одной стороны, к определенной аминокислоте, с другой — к молекуле соответствующей ей транспортной РНК.
Образование комплекса аминокислоты с молекулой транспортной РНК без посредничества ферментов совершенно невозможно. Ибо если бы даже эта парочка и научилась «распознавать» друг друга (такое вполне мыслимо), то соединяющая их связь уж никак не могла бы возникнуть сама по себе. На ее образование нужно затратить некоторое количество энергии, а реакции такого типа, как мы знаем, в организме выполняются только при участии ферментов.
Итак, первый этап биосинтеза белковой молекулы завершен: аминокислота прочно связана с соответствующей транспортной РНК. Важнейший же инструмент второго этапа — сравнительно крупные частицы, называемые рибосомами (мы уже упоминали о них, когда шла речь о процессах самосборки). Конечно же, «крупные» они только по молекулярным масштабам: рибосома примерно в сто раз «тяжелее» средней белковой молекулы. Ее размер — около двухсот ангстрем. Рибосома состоит из двух неравных субъединиц, различающихся по величине примерно вдвое. «Материал», из которого построены рибосомы, — это белки и РНК, но опять-таки РНК совершенно особого типа.
Рибосомная РНК, так же как и транспортная, не содержит информации об аминокислотной последовательности каких-либо белков; кстати, и вообще не очень понятно, зачем она присутствует в рибосомах (как видите, молекулярная биология тоже кое-чего не знает!).
Термин «инструмент», примененный нами выше в отношении рибосомы, возможно, не очень точен. Правильнее было бы, наверное, назвать ее рабочим столом, на котором идет синтез белка. Происходит это следующим образом. Рибосома связывается с нитью информационной РНК в том месте, где начинается запись аминокислотной последовательности некоторого белка. Механизм «узнавания» этого места легко себе вообразить: в состав рибосомы входит РНК, и участок, комплементарный какой-то «метке» на информационной РНК, может уже известным нам образом связаться с упомянутой «меткой». После этого рибосома готова к приему первого комплекса транспортной РНК с аминокислотой.
Этот комплекс взаимодействует как с самой рибосомой, так и с триплетом информационной РНК, следующим за «меткой». «Сцепление» транспортной РНК с информационной основано на том, что в определенной части структуры молекулы транспортной РНК есть так называемый антикодон — тройка оснований, комплементарная триплету, — который кодирует данную аминокислоту в информационной РНК. Взаимодействия, возникающие внутри комплекса рибосомы с обеими молекулами, приводят к некоторым его перестройкам. Молекула аминокислоты отщепляется от молекулы транспортной РНК и связывается с рибосомой; последняя, в свою очередь, несколько перемещается вдоль молекулы информационной РНК. После этого транспортная РНК отделяется.
(Такой образ действий молекулы транспортной РНК позволяет понять происхождение другого варианта ее названия — «мессенджер-РНК», или просто «мессенджер». О том, что это иноязычное заимствование, нечего и говорить. Остается только объяснить, откуда он взялся.
«Стэнли Мак-Брайд еще раз взглянул на часы. Со времени звонка Мейсона прошло уже около двадцати минут, но посыльный все еще не появлялся. Если он не придет в течение последующих трех минут, Стэнли вынужден будет покинуть „Карлтон-отель“: он знает, что полиция не слишком снисходительна к нарушителям сухого закона.
В дверь постучали. Со вздохом облегчения Стэнли поднялся из кресла. На пороге стоял посыльный в расшитой галунами курточке:
— Пакет для вас, сэр…»
Так вот, если подобного рода сочинения читать в английском оригинале, то выяснится, что этот самый посыльный с его непременными галунами и есть мессенджер. Вот откуда берет свое начало термин «мессенджер-РНК»; понятно, что и к нему могут быть предъявлены те же упреки, которые страницей раньше мы предъявляли к выражению «транспортная РНК».)
В результате перемещения рибосомы напротив того места, где происходит посадка на нее транспортной РНК, оказывается уже другой триплет, и теперь та же история повторяется с новой молекулой транспортной РНК. Различие заключается единственно в том, что «доставленная» ею молекула аминокислоты при участии специальных ферментов вовлекается в образование пептидной связи с уже имеющейся на рибосоме аминокислотой. Опять сдвиг рибосомы относительно молекулы информационной РНК ровно на один триплет, опять с рибосомой связывается новая молекула транспортной РНК, к которой «прикреплена» очередная аминокислота, и так далее. До того момента, когда рибосома дойдет до очередной «метки» на молекуле информационной РНК, «метки», сигнализирующей окончание последовательности белковой молекулы. Затем рибосома отделяется от нити РНК, а готовая белковая молекула — от рибосомы; синтез завершен.
Иными словами, если уж сравнивать процесс синтеза белковой последовательности на матрице информационной РНК, как это часто делается в популярной литературе, с переводом с одного языка на другой, то нужно говорить о двойном переводе: сначала, скажем, японский текст с помощью японско-английского словаря переводится на английский язык, после чего уже с помощью англо-русского словаря — на русский.
Образование комплекса аминокислоты с транспортной РНК является, по этой схеме, лишь первым этапом перевода, а роль «словаря» здесь выполняет набор обоюдоспецифичных ферментов. (Возможно, некоторые из наших читателей, разобравшись в описанном процессе, еще раз посетуют на излишнюю сложность и запутанность молекулярных механизмов биологического кодирования. Но, во-первых, все эти механизмы — повторим еще и еще раз — обслуживаются практически никогда не ошибающимися ферментами, а во-вторых, попробуйте-ка сами перевести без ошибок хотя бы несколько японских фраз!)
На втором же этапе роль словаря играет транспортная РНК, поскольку именно она поставляет каждому триплету соответствующую аминокислоту. Кстати, здесь может возникнуть вопрос: большинство аминокислот, согласно генетическому коду, кодируется несколькими триплетами, значит, и транспортных РНК должно быть не двадцать типов, а гораздо больше? Так оно и есть на самом деле, по крайней мере, в некоторых организмах. Правда, до сих пор не удалось установить, действительно ли в клетке всегда присутствуют все типы различных транспортных РНК (по числу триплетов, кодирующих аминокислоты), но ясно, что их, во всяком случае, больше двадцати.
Изложенная «принципиальная схема» биосинтеза белка вызывает, помимо переводческо-лингвистической, и иную, чисто производственную аналогию. В самом деле, так и кажется, что если попытаться снять фильм о работе этого уникального по совершенству механизма, созданного природой, то «звукоряд» такого фильма будет состоять из четкого ритма шумов огромного производственного комплекса: вот лязгнула аминокислота, присоединенная аминоацилсинтетазой к транспортной РНК, щелкнул «контакт» транспортная РНК — информационная РНК, рибосома со скрипом продвинулась по информационной РНК на один триплет и так далее.
Такая аналогия тем более оправдана, что синтез молекулы белка на рибосоме осуществляется сравнительно быстро, особенно учитывая филигранную точность работы: на это, в зависимости от размеров молекулы, расходуется от нескольких секунд до нескольких десятков секунд. При этом надо учесть, что в целях повышения «производительности» на одной молекуле информационной РНК одновременно «сидит» несколько рибосом, и тем самым синтезируется сразу несколько молекул белка. Были даже получены доказательства синтеза белка на молекуле информационной РНК, которая еще сама находилась в процессе изготовления. Иными словами, конец этой молекулы еще не был готов, а начало уже вовлечено в работу по синтезу белка!
Из последнего факта следует, кстати, один важный вывод: направление наращивания новых звеньев молекулы РНК в процессе ее синтеза совпадает с направлением движения вдоль нее рибосомы в процессе синтеза белка. А именно, молекула белка строится в направлении от конца, несущего свободную аминогруппу (N-конца), к концу, несущему карбоксильную группу (C-концу).
Вот откуда берутся белки. Заметим, в нашем описании мы не использовали никаких представлений, не упоминали никаких процессов, которые не могли бы быть описаны на языке химии и физики. По современному состоянию биологической науки это описание в некоторых местах может быть более подробным, в некоторых — менее, модели одних процессов представляются вполне очевидными, в отношении других, как говорится, возможны варианты, но не подлежит сомнению главное: чтобы объяснить самые существенные, самые (эх, была не была!) откровенные процессы, происходящие в живой клетке, нам на сегодняшний день уже абсолютно нет необходимости предполагать существование каких-то сил, отличных от уже известных химикам и физикам!
В том, что эта пламенная тирада — не пустой звук, сможет убедиться всякий по-настоящему терпеливый читатель (впрочем, только такому и удастся дочитать ее до конца). Авторы берут на себя смелость утверждать: того, что мы уже знаем о белках и нуклеиновых кислотах, достаточно (или почти достаточно) для понимания, по крайней мере в главных чертах, сущности явления, называемого жизнью. Правда, на «философские» вопросы типа «В чем смысл жизни?» мы, по присущей нам скромности, не дадим ответа, но вопросы биологические, поверьте, будут, хоть и схематично, рассмотрены на примере захватывающей истории о жизни и повадках хорошо (увы!) знакомого каждому читателю существа — вируса.
Кое-что о вирусе
Биологи, которые решились бы, вдохновленные примером знаменитого немецкого зоолога А. Брема, создать многотомный труд «Жизнь вирусов», наверняка бы не уставали с обидой в голосе повторять, что перед их великим предшественником стояла задача неизмеримо более благодарная.
В самом деле, столько увлекательных возможностей: тут и коварные охотничьи повадки пумы, и хитроумные брачные игры птиц, и диковинной формы глубоководные рыбы. Чего стоят одни цветные вклейки с изображениями попугаев! А для придания вящей достоверности А. Брем еще приводит красочные рассказы путешественников-натуралистов, своими глазами видевших, например, схватку орла с гиеной.
Про вирус что можно рассказать занимательного? Форма у него не очень диковинная, а часто и весьма простая — палочковидная или многогранная, очевидцев (тех, кто видел вирус, скажем, в электронный микроскоп) немного, а что до экзотичности, то почти каждый из нас сталкивался с этими «милыми» существами — хотя бы в период зимне-весенних эпидемий гриппа. И все же попытаемся описать экстерьер, образ жизни и «привычки» вируса.
Прежде всего, вирус весьма-весьма мал. По своим линейным размерам он всего-то в тысячу или несколько тысяч раз больше молекулы водорода, то есть можно говорить, что он имеет размеры, сравнимые с молекулярными. В самом деле, средняя молекула белка выглядела бы рядом со средним вирусом примерно как пачка сигарет рядом с платяным шкафом.
Вирусная частица состоит из белковой оболочки, внутри которой заключена нуклеиновая кислота — в зависимости от типа вируса это может быть ДНК или РНК. Вот и весь организм.
Вирусы ведут омерзительно паразитический образ жизни, не изобилующий, однако, никакими впечатляющими подробностями. Сталкивается этакий вирус, например, с бактериальной клеткой, прилипает к ее стенке. Через некоторое время нуклеиновая кислота проникает внутрь клетки, оставив белковую оболочку снаружи. Часто в описаниях этого процесса используется выражение «впрыскивание ДНК», однако не следует понимать его буквально: вирус начисто лишен какой-либо способности к активным действиям; речь идет просто о перестройке белковых структур вируса под влиянием физико-химических взаимодействий с поверхностью клетки и о диффузии ДНК внутрь клетки.
ДНК, проникшая в клетку, находит там (естественно!) весь комплекс ферментов, необходимых для обеспечения процесса репликации (не забыли? Это процесс воспроизведения ДНК!). И вот в клетке начинается массовое изготовление копий вирусной ДНК, которые ей, клетке, мягко говоря, совершенно не нужны. Ибо дальнейшие события уже легко предвидеть: другие ферментные системы клетки, ответственные за транскрипцию (а это процесс производства РНК по ДНК), принимаются за синтез комплементарных копий РНК, которые служат матрицами для изготовления (снова при помощи ферментных систем клетки) молекул вирусных белков. Десятки или даже сотни молекул белка вирусной оболочки самопроизвольно образуют определенной структуры комплекс с одной из имеющихся в клетке молекул вирусной ДНК (наподобие того, как это происходит при формировании четвертичной структуры белка — подробней об этом чуть попозже). Получившийся комплекс и есть не что иное, как новая вирусная частица. Такой процесс, осуществляющийся за счет ресурсов клетки и силами ее ферментативных систем, протекает весьма быстро.
При этом изощренное издевательство вируса над беззащитной клеткой не исчерпывается описанными вероломными штучками. Помимо белков оболочки вируса, на вирусных РНК синтезируются и некоторые другие белки. Структура их такова, что, образуя прочные комплексы с некоторыми ферментами клетки, они блокируют процесс синтеза клеткой собственных белков и нуклеиновых кислот с тем, чтобы все ее оставшиеся силы были направлены только на воспроизводство новых вирусов. (Какое хищное, жестокое коварство — причем, заметьте, коварство на молекулярном уровне!)
Кроме того, на той же вирусной РНК часто идет еще и синтез фермента, разрушающего клеточную оболочку, чтобы к тому моменту, когда полностью истощенная, кишащая вирусами клетка погибнет, мириады зловредных существ смогли беспрепятственно устремиться наружу. Правду, правду говорят врачи: во время эпидемий гриппа нужно пользоваться марлевыми повязками.
Впрочем, мы вынуждены разочаровать любителей самолечения: никаких новых рекомендаций по борьбе с вирусными заболеваниями мы не предложим — ни всеисцеляющей гимнастики йогов, ни лечебного голодания. Наши цели в другом: описав полный жизненный цикл вируса, мы еще раз хотим подчеркнуть роль, которую играют в этом цикле белки. «Прикрепление» белковой оболочки вируса к стенке клетки, проникновение вирусной ДНК в клетку, сопровождающееся перестройкой белков мембраны клетки и оболочки вируса, использование различных ферментных систем для размножения, самосборка вирусной частицы — без белковых молекул, без их удивительных свойств, «заложенных», как мы знаем, в пространственной структуре молекулы, все эти процессы попросту немыслимы.
Кстати о самосборке: вирусные частицы в клетке формируются, как упоминалось, самопроизвольно, без какого-либо внешнего направляющего начала. При этом нуклеиновая кислота вируса как бы экранируется белковым слоем, образованным сотнями или тысячами молекул-субъединиц, которые располагаются в слое геометрически правильно. Так, палочковидная частица одного из вирусов имеет оболочку, в которой отдельные белковые субъединицы расположены по спирали; на поверхности частиц, имеющих форму многогранников, субъединицы образуют рисунок, напоминающий паркет, выложенный из шестиугольных плиток. Все эти структуры, повторяем еще раз, собираются самопроизвольно: если в определенных условиях разделить белковую и нуклеиновую части вирусов, а затем опять их смешать, получатся вполне нормальные вирусы. Таким образом удается даже получать «гибридные» вирусы, у которых нуклеиновая кислота принадлежит одному виду, а белковая оболочка — другому.
Непосредственное влияние третичной структуры белковых субъединиц на процесс самосборки можно представить себе, например, так: предположим, что «боковые» поверхности субъединиц несут много гидрофобных групп, благодаря чему в воде они стремятся «сойтись» именно этими поверхностями. Наоборот, «наружный торец» имеет много полярных групп и хорошо смачивается водой, а «внутренний торец» обладает сродством к нуклеиновой кислоте. Под влиянием соответствующих межмолекулярных взаимодействий и осуществляется самосборка вирусной частицы. Естественно, помимо всего прочего, пространственное строение белковых субъединиц должно быть таким, чтобы образуемая ими оболочка имела плотную и правильную структуру.
Однако мы вновь отвлеклись (хотя и по благодарному поводу: не правда ли, всегда приятно еще раз затронуть тему о пространственной структуре белковых молекул?) от нашего главного направления: схематического объяснения феномена жизни на языке физики и химии — наук, иногда называемых точными (боже, какая несправедливость в отношении биологии!). Что ж, вернемся к нашим вирусам.
Такова жизнь
Авторы прекрасно отдают себе отчет: читатель вправе быть разочарован. После долгих и зачастую нудных объяснений, относящихся к самым разнообразным наукам, после многозначительных ссылок на квантовую механику, историю и даже лингвистику, наконец, после твердого обещания показать, как же разрешается эта самая «многовековая загадка жизни», читателю было предложено едва ли три странички текста, на которых, как утверждают авторы, и содержится описание основных жизненных процессов. Да полноте! Неужели эта цепочка химических реакций и физических явлений и есть жизнь? И стоило ли ради этих трех страниц городить предыдущие пять глав?
Безусловно, стоило. Как мы видим теперь, весь жизненный цикл типичного вируса представляет собой последовательность физико-химических процессов из числа рассмотренных нами ранее. Конечно, по сравнению с циклом прочих живых организмов он чудовищно примитивен; недаром одно время в среде биологов разгорелся несколько схоластический спор на тему о том, называть ли вирус существом или веществом. Сразу видно, однако, что это спор скорее на лингвистическую (опять!), чем на биологическую, тему: в действительности вирус подпадает и под то и под другое определение, так что речь может идти лишь о границах применения обоих терминов.
И все-таки хочется вслед за каким-нибудь квартирным склочником воскликнуть:
— Разве это жизнь?
Давайте взглянем, чем же вирус отличается от прочих живых существ. Прежде всего, будучи способным к размножению, он использует для этой цели не принадлежащие ему «химическое сырье» и ферментные системы клетки-хозяина. В случае же более сложно организованных существ (опять это слово!) в наследственной инструкции — геноме — должна содержаться также в конечном счете запись (в виде цепочки нуклеотидов) аминокислотной последовательности всех ферментов, необходимых для обеспечения процессов репликации, транскрипции, белкового синтеза, а также ферментов, производящих необходимые для этих процессов исходные соединения.
Конечно, количество информации, записанной в виде некоторой последовательности ДНК, в результате этого возрастет неимоверно, однако такой организм в отличие от вируса уже будет полностью самостоятельным, автономным. И все же столь ли уж принципиальным будет это отличие, заключающееся, в конце концов, всего-то в величине общей протяженности ДНК-овых цепочек?
Проделаем мысленный эксперимент: сначала удлиним вирусную ДНК за счет последовательности нуклеотидов, обеспечивающей синтез одного из ферментов обслуживающих систем. Существо такого рода, очевидно, по-прежнему будет для своего размножения нуждаться в ферментных системах клетки-хозяина. Однако, включая в состав генома вируса последующие ферменты, мы будем все более усиливать независимость вновь создаваемого организма от услуг ферментов клетки, так что на каком-то этапе можно будет говорить о возможности самостоятельного его существования.
На первый взгляд это рассуждение, напоминающее известный парадокс древних о зерне и куче (одно зерно не есть куча, два зерна не есть куча, три зерна… и так далее до любого количества зерен, которые все никак не могут образовать кучу), легко уязвимо: ясно, что организм станет «настоящим» после включения в состав генома последнего из необходимых для воспроизводства ферментов. Именно этот момент перехода к абсолютной самостоятельности можно было бы, казалось, назвать решающим в отнесении изучаемого объекта к живой или мертвой материи.
Но вот незадача: микробиологической практике известно сколько угодно примеров существования мутантных форм бактерий-уродцев, лишенных способности синтезировать какой-то один фермент. Если это, например, один из ферментов, участвующих в синтезе определенной аминокислоты — скажем, аланина, — то такой организм не сможет существовать вполне самостоятельно, но будет прекрасно развиваться в среде, содержащей аланин. И уж конечно, ни у кого не хватит совести отказать ему в праве называться живым. (Точно так же, как никто не усомнится в праве завода, который занят лишь сборкой изделий из агрегатов, поставляемых смежниками, называться, скажем, заводом холодильников или телевизоров.)
Словом, не подлежит сомнению, что вирусы — одно из проявлений жизни. А то обстоятельство, что для современной науки не являются секретом существеннейшие детали его бытия, позволяет утверждать, что нам, людям второй половины XX века, стало наконец известно в принципе, что такое жизнь.
На примере вирусов мы рассмотрели некую минимальную совокупность процессов, характерных для всех без исключения живых существ. Эти процессы и составляют самую основу жизни. Более высокоразвитые организмы, отличающиеся неизмеримо более сложным устройством и чрезвычайным разнообразием жизненных отправлений, используют в точности те же механизмы кодирования, синтеза макромолекул; в них работают ферменты точно такой же природы. Не следует, конечно, на этом основании думать, что существует угроза «закрытия» биологии как «науки о жизни» по той причине, что ей уже все известно. Как раз наоборот — в этой области мы на сегодняшний день не знаем неизмеримо более, чем знаем. Однако установление центральных молекулярных механизмов организации и функционирования всякого живого существа навсегда останется заслугой биологии 50–70-х годов нашего и без того очень бурного столетия.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава пятая Жизнь человекообезьян в сообществах
Глава пятая Жизнь человекообезьян в сообществах Тот, у кого в роду нет ни глупцов, ни плутов, ни попрошаек, был зачат ударом молнии. Томас Фуллер (1608–1661) В Гомбе-Стрим, в Танзании, самец-шимпанзе играет с маленьким братом. Во время игры малыш легко стоит на двух ногах, что
Глава III. Есть ли жизнь на Земле?
Глава III. Есть ли жизнь на Земле? Оро:…Как сказал один шутник, по теории вероятностей мы все должны были быть мертвецами. Тем не менее, мы живы. Чаргафф: Но мы все-таки умрем. Мора: В том-то и беда. Из дискуссии на Флоридской конференции по происхождению жизни Там, где вечно
ГЛАВА 13. Есть величие в этом взгляде на жизнь
ГЛАВА 13. Есть величие в этом взгляде на жизнь В отличие от своего деда-эволюциониста Эразма, чьими стихами на научную тематику (несколько неожиданно, я бы сказал) восхищались Вордсворт и Колридж, Чарлз Дарвин не был известен как поэт, но он создал лирическую кульминацию в
Глава 6 ЖИЗНЬ РАСТЕНИЙ, УВЕЛИЧЕННАЯ В 100 МИЛЛИОНОВ РАЗ
Глава 6 ЖИЗНЬ РАСТЕНИЙ, УВЕЛИЧЕННАЯ В 100 МИЛЛИОНОВ РАЗ а восточном побережье Индии, в старинном штате Бенгал, на полутора гектарах земли неподалеку от Университета Калькутты расположился комплекс зданий из серо-лилового песчаника в классическом до мусульманском
Глава 3 Дыхание и жизнь
Глава 3 Дыхание и жизнь Из всего разнообразия веществ, находящихся в окружающей среде, воздух, или, точнее, кислород, является самым жизненно необходимым. Если нужно, мы можем несколько дней обходиться без воды, а без пищи неделями. Я не хочу сказать, что это приятный
Глава 1. Что такое жизнь?
Глава 1. Что такое жизнь? Не так уж много времени прошло с тех пор, как генетика и биохимия стали самостоятельными науками, каждая из которых… пытается подобрать ключ к феномену жизни. Биохимики обнаружили ферменты, а генетики — гены. Уильям Хеш, "Генетика бактерий и
Глава 4. Есть ли жизнь на других планетах?
Глава 4. Есть ли жизнь на других планетах? Тем не менее большинство планет, несомненно, обитаемы, а необитаемые со временем будут населены. Таким образом, я могу все изложенное выше выразить в следующем общем виде: вещество, из которого состоят обитатели различных планет, в
Глава 8. Жизнь в Солнечной системе
Глава 8. Жизнь в Солнечной системе Нам суждено спускаться вновь и вновь. В тот край, откуда началась дорога, Чтобы опять взглянуть… и в изумленьи Его увидеть, словно в первый раз. Т. С. Элиот, "Легкое головокружение" Убежденность в существовании жизни на планетах Солнечной
Глава 20. ЖИЗНЬ БЕЗ ВОЗДУХА
Глава 20. ЖИЗНЬ БЕЗ ВОЗДУХА Как видите, многое о процессах, происходящих в живой ткани, можно сказать на одной лишь основе рассмотрения веществ, которые мы имеем на входе в организм и на выходе из него. Мы решили, что должен существовать некий общий строительный «кирпичик»,
Глава 23. ЖИЗНЬ С ВОЗДУХОМ
Глава 23. ЖИЗНЬ С ВОЗДУХОМ При рассмотрении реакций, проходящих с участием атмосферного кислорода, естественно возникает желание разобраться в самом процессе впитывания кислорода живой тканью (ну, наполняет он легкие, и что дальше?).Из таких разных существ, как картошка и
Глава 10. Как давно могла зародиться жизнь?
Глава 10. Как давно могла зародиться жизнь? До сих пор мы обсуждали, где могла бы появиться жизнь во Вселенной и насколько маловероятным могло быть это событие. Мы совсем не обсуждали, когда она могла зародиться и сколько времени потребовалось на совершенствование древних
Короткая жизнь для человека, но длинная жизнь для человечества
Короткая жизнь для человека, но длинная жизнь для человечества Бактерии и люди многим отличаются. Если одно поколение бактерий живет двадцать минут, то между одним поколением человека и следующим проходит много лет. Если человек зарождается за счет слияния яйцеклетки и
Глава 16: Жизнь, какой мы (не)знаем ее
Глава 16: Жизнь, какой мы (не)знаем ее Caplan, Arthur. What If Anything Is Wrong with Cloning a Human Being? // Case Western Reserve Journal of International Law. 35 (Fall 2003): 69–84.Segerstr?le, Ullica. Defenders of the Truth. Oxford University Press, 2001.Wade, Nicholas. Before the Dawn. Penguin, 2006.* Среди прочих, Николас Уэйд предлагал добавить еще одну пару
Глава 17 Голубая планета и новая жизнь
Глава 17 Голубая планета и новая жизнь Земная жизнь в безбрежном лоне вод Среди пещер жемчужных океана Возникла, получила свой исход, Росла и стала развиваться рано; Сперва в мельчайших формах все росло, Не видимых и в толстое стекло, Которые, киша, скрывались