Глава 21. ЗНАЧЕНИЕ ФОСФАТОВ
Глава 21.
ЗНАЧЕНИЕ ФОСФАТОВ
Перед тем как обсуждать связь между фосфатными группами и метаболизмом, надо сначала немного поговорить о самом фосфоре.
В таблице Менделеева фосфор находится сразу же под азотом, что свидетельствует о сходстве атомного строения этих двух элементов. У атома азота две электронные оболочки, во внешней из них находится пять электронов, а у атома фосфора — три оболочки, во внешней из которых тоже находится пять электронов. Соответственно, в электронном изображении фосфор можно представить как

то есть точно так же, как и азот.
Атом фосфора может принимать в совместное пользование три электрона других атомов, предоставляя при этом в совместное пользование и три своих, так чтобы в целом достигать стабильного числа восемь. Так, например, с водородом фосфор образует соединение (фосфин, РН3) таким образом:
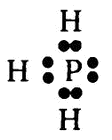
Точно так же образуется и аммиак (см. главу 17).
Кроме трех электронов, которые фосфор может предоставлять в совместное пользование, у него есть еще два, которые он никуда предоставлять не собирается (к примеру, в фосфине они остаются в полном распоряжении фосфора). Однако эти два электрона вполне могут быть предоставлены в пользование атому, которому только их и не хватает для обретения заветных восьми электронов во внешней оболочке. Естественно, первым из такого рода атомов нам приходит в голову кислород, во внешней оболочке которого всего шесть атомов.
Если сосредоточиться только на фосфоре и кислороде, то можно представить «разделяемые» его атомы как «просто связи» и рассматривать только «неразделяемые», таким вот образом:
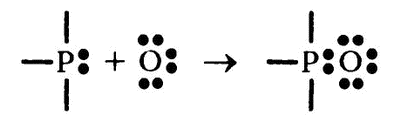
Как видите, теперь потребность атома кислорода в восьми электронах удовлетворена без каких-либо потерь для фосфора, который также сохраняет эти электроны в своей внешней оболочке.
Связь, образуемая подобным образом — когда оба электрона предоставляются одним и тем же атомом, — так же сильна, как и образуемая обычным способом, когда каждый атом предоставляет по одному электрону для образования пары. В химии она именуется «координационной связью», и ее часто изображают на схемах в виде маленькой стрелочки, указывающей от того атома, который предоставляет электроны, на тот, который их принимает. Таким образом, фосфор имеет три ковалентных и одну координационную связь, что можно изобразить так:
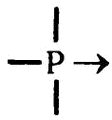
Зачастую, как в случае с фосфином, задействованными оказываются только три ковалентные связи. Однако, особенно в тех случаях, когда речь заходит об атомах кислорода, для координационной связи тоже находится работа[7]. Кстати, все вышесказанное в равной степени относится и к атомам азота.
Самым важным фосфорсодержащим соединением является ортофосфорная кислота, которую часто называют просто «фосфорной кислотой» (Н3РO4), в которой всеми четырьмя связями фосфор соединен с атомами кислорода. Ее можно представить таким образом:
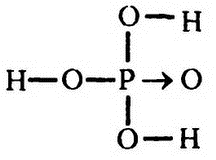
Где бы в живой ткани ни появлялся атом фосфора, он всегда оказывается входящим в состав соединения, родственного фосфорной кислоте. К примеру, фосфорная кислота может ионизироваться, сначала за счет ионизации одного атома водорода (Н+), в результате чего получается «первичный фосфат-ион» Н2РО4-, с отрицательным зарядом -1, а затем — второго и третьего атомов водорода с образованием вторичного и третичного фосфат-ионов: НРО42- и РО43-. Первые два из этих ионов обнаруживаются и в крови, и в тканях.
Кроме того, фосфорная кислота или получаемые из нее ионы могут вступать в соединение с другими веществами. В таких соединениях участвуют (иногда и более одной) гидроксильные (содержащие и водород, и кислород) группы, присутствующие в молекуле. Мы сосредоточимся именно на этой части молекулы фосфорной кислоты и, краткости и удобства ради, обозначим оставшуюся часть как (P). В таком виде краткая запись молекулы фосфорной кислоты будет выглядеть как Н — О — (P). В тех случаях, когда речь будет идти более чем об одной гидроксильной группе, к центральной (P) будут обозначены присоединенными вторая и третья гидроксильные группы.
При вступлении в соединение фосфорная кислота или ее ионы обычно непосредственно соединяются с гидроксильной группой другого вещества, что можно изобразить так: R — О — Н, где R — любая углеродосодержащая группа. Таким образом, запись R — О — Н может обозначать и спирт, и сахар или другие соединения, о которых мы не упоминали, — в общем, любое вещество, в состав которого входит гидроксильная группа.
Таким образом, реакцию фосфорной кислоты, скажем, с сахаром можно представить на рис. 45. В процессе присоединения отщепляется молекула воды, то есть мы имеем дело с реакцией конденсации, значит, реакцию обратную данной следует признать реакцией гидролиза.
Обычно уменьшение свободной энергии связано именно с гидролизом, так что точка равновесия данной реакции смещена далеко в сторону смеси сахара и фосфорной кислоты. При гидролизе сахарофосфата (примером такого вещества является упомянутый в предыдущей главе эфир Хардена—Янга) высвобождается от 2 до 4 килокалорий на моль вещества, в зависимости от конкретного вещества. То есть при гидролизе эфирной связи сахарофосфата высвобождается примерно столько же свободной энергии, сколько и при гидролизе пептидной связи белковой молекулы.
Естественно, для того, чтобы сформировать эфир путем конденсации сахара и фосфорной кислоты, необходимо затратить те же 2—4 килокалории на моль. Значит, чтобы создать такого рода эфирную связь, надо потратить столько же энергии, сколько на создание пептидной связи.
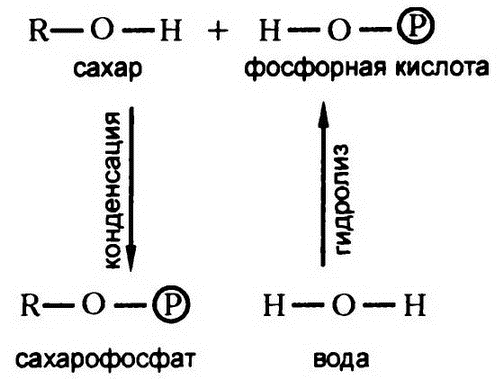
Рис. 45. Фосфорная кислота и сахар
Поскольку фосфорная кислота обладает гидроксильной группой, то одна молекула фосфорной кислоты вполне может конденсироваться и с другой такой же молекулой. В результате получается сочетание атомов типа — О — (|3), «пирофосфатная связь».
Конденсированные кислоты, приведенные на рис. 46, оставались бы вне поля зрения органической химии, если бы не тот факт, что в 1929 году немецкий биохимик К. Лохманн сумел выделить из мышц вещество, оказавшееся продуктом конденсации трифосфорной кислоты с органическим веществом аденозином, встречающимся в составе нуклеиновых кислот[8]. Соответственно, продукт конденсации получил название «аденозинтрифосфат».
Постепенно обнаруживалось участие аденозинтрифосфатов (теперь их принято обозначать общеизвестным уже сокращением АТФ) в различных реакциях, протекающих в организме. Они участвуют в процессе сокращения мышц; в передаче нервных сигналов; практически во всех стадиях обмена веществ.
Почему? Ну, начать хотя бы с того, что две фосфатные группы на удаленном от аденозина конце молекулы легче подвергаются гидролизу, чем фосфатные связи в целом. Это свидетельствует о том, что при гидролизе пирофосфатной связи уровень свободной энергии уменьшается сильнее, чем при гидролизе обычной фосфатной связи. Исследования подтвердили это предположение. Фриц Липман, американский физик немецкого происхождения, предположил по результатам своих исследований начала 40-х годов XX века, что существуют два типа фосфатных связей — обычные фосфатные связи, как в сахарофосфате, «низкоэнергетические», и «высокоэнергетические», примером которых является пирофосфатная связь.
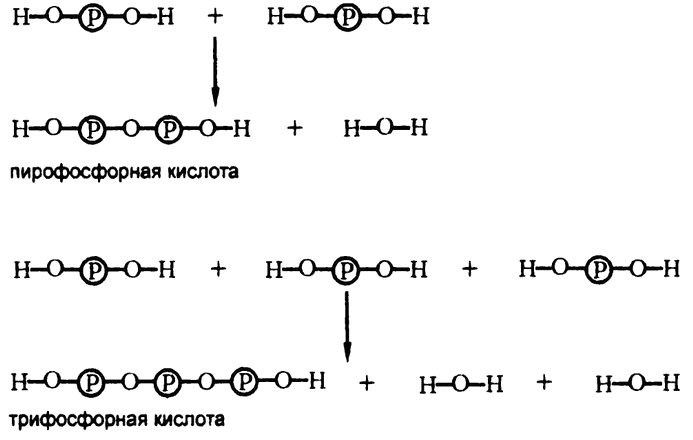
Рис. 46. Пирофосфатная связь
Сперва выяснилось, что при гидролизе высокоэнергетической фосфатной связи высвобождается от 11 до 16 килокалорий энергии на моль вещества, просто колоссальная цифра по сравнению с двумя—четырьмя килокалориями на моль, высвобождаемыми при гидролизе обычной фосфатной связи. Однако дальнейшие исследования показали, что эта цифра завышена. В 50-х годах XX века уточненные исследования показали, что в среднем при гидролизе высокоэнергетических фосфатных связей высвобождается от 5 до 10 килокалорий, в среднем — 8. А при гидролизе некоторых фосфатных связей, которые принято было считать низкоэнергетическими, могло высвобождаться и до 6 килокалорий на моль. Получается, никакого строгого разрыва между этими двумя группами нет вообще.
Но все же остается признать, что существует один тип фосфатных связей, при гидролизе которых выделяется в среднем 8 килокалорий на моль вещества, и другой, при гидролизе связей которого выделяется в среднем 4 килокалории на моль. Хотя различие между ними не так велико, как показалось сначала, и некоторые биохимики уже начинают колебаться, есть ли оно вообще, я все же считаю оправданным разделение фосфатных связей на высоко- и низкоэнергетические.
Многие биохимики по привычке разделяют высоко- и низкоэнергетические фосфатные связи графическим обозначением валентной связи в формуле. Низкоэнергетическая связь обозначается обычным образом R — О — (P), а высокоэнергетическая фосфатная связь — волнистой линией. Так, обозначив АТФ за А, можно представить формулу таким образом:
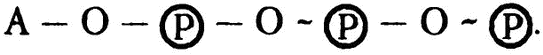
Последние две связи, как видите, — пирофосфатные и высокоэнергетические. Если одну из них разорвать посредством гидролиза, то останется «аденозиндифосфат», или АДФ, формула которого выглядит как А — О — (P) — О — (P). Если разорвать еще одну, то останется «аденозинмонофосфат», или АМФ (он же «адениловая кислота»), формула которого А — О — (P). Теперь осталась только низкоэнергетическая фосфатная связь. Ее тоже можно разорвать с помощью гидролиза, так чтобы остался только аденозин, но энергии при этом третьем по счету гидролизе выделится немного — в два раза меньше, чем в любом из предыдущих случаев.
Помимо иирофосфатной, существуют и другие высокоэнергетические фосфатные связи. Возьмем органическую кислоту в общем случае (общую формулу можно записать так:
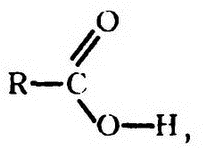
где R — любое из бесчисленного множества углеродсодержащих соединений, а СООН — «карбоксильная группа»), с которой мы уже не раз встречались.
Если фосфорная кислота конденсируется с гидроксильной частью карбоксильной группы (как на прилагаемой диаграмме), то в результате получается «ацилфосфат», изображаемый с волнистой связью, чтобы подчеркнуть ее высокую энергию (рис. 47).
Теперь давайте посмотрим, как высокоэнергетическая связь вступает в игру. Для этого вернемся к гликогену. Гликоген, содержащийся в печени и мышцах животных, в подробностях строения не совсем похож на крахмал, хранящийся в «запасниках» растений, но и гликоген, и крахмал формируются из молекул глюкозы, которые конденсируются с выделением при этом воды. Когда крахмал расщепляется в пищеварительном канале, амилазы катализируют процесс гидролиза единиц глюкозы на отдельные молекулы, добавляя воду обратно и разбивают крахмал на все более короткие цепочки, пока не получаются снова отдельные молекулы глюкозы.
И печень, и мышцы тоже должны уметь расщеплять гликоген на отдельные молекулы глюкозы, чтобы поддерживать должный ее уровень в крови. Так что логично предположить, что в тканях тоже работает некий фермент, подобный амилазам, который выполняет сходную работу — катализирует гидролиз.
Для проверки этого предположения необходимо было придумать такой эксперимент, который позволил бы понять, какие именно реакции происходят в живых тканях. Проще всего было бы сделать это, выбрав в качестве живой пробирки какое-нибудь подходящее животное вроде мыши. К сожалению, даже мышь — слишком сложное для такой задачи существо, в ее организме одномоментно проходят тысячи реакций.
Поэтому для упрощения задачи мышь придется убить и взять для исследования крошечные кусочки ее печени и мышц. Если правильно извлечь эти ткани и быстро подвергнуть обследованию, можно застать в образцах ферменты в работоспособном состоянии. Такие образцы помещали в растворы различных химических веществ. Действие ферментов настолько специфично, что если та или иная ткань катализирует определенную реакцию в лабораторных условиях, то вполне вероятно, что и в живом организме она катализирует ту же реакцию.
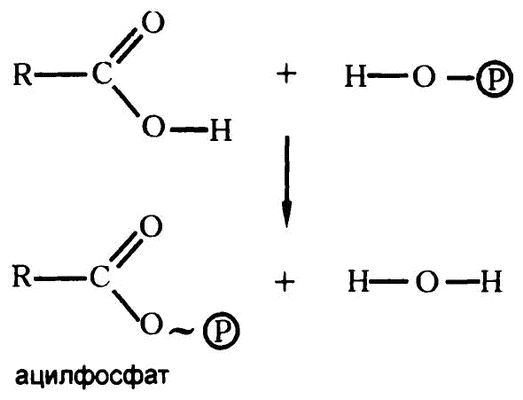
Рис. 47. Ацилфосфат
Если необходимо ускорить дело, то образец можно растолочь до полужидкой гомогенной кашицы («гомогената»), которую и поместить в субстрат. Тогда не придется ждать, пока субстрат проникнет в клетки—в процессе размешивания все ферменты лишились защиты клеточных мембран, и ничто не мешает им теперь проявлять каталитические свойства.
В гомогенате тоже содержится очень много ферментов, и обычно требуется сократить их число во избежание побочных реакций, которые могут помешать, и для того, чтобы концентрация необходимого фермента оказалась более высокой, чем в живом организме.
Это достигается путем подвергания гомогената условиям, о которых известно, что в них один фермент отделяется от другого. При помещении материала в некоторые растворы одни белки втягиваются в раствор, другие остаются в гомогенате. Отфильтровав после этого смесь, мы получим раствор, в котором содержатся только растворимые белки, «экстракт ткани», или «вытяжка из ткани». Получившуюся вытяжку можно дальше подвергнуть воздействию определенных кислот, ионов или некоторых органических растворов при низких температурах. При каждом из этих действий одни ферменты будут осаждаться, а другие останутся в растворе. Если повезет — можно дойти и до такого этапа, когда в растворе останется вообще только один, нужный, фермент.
Именно в результате длительной работы с вытяжками из ткани чета американских биохимиков чешского происхождения, Карл Кори и Герти Кори, сумели показать, что расщепление гликогена в тканях не имеет ничего общего с гидролизом и не катализируется амилазами.
В 1935 году К. и Г. Кори обнаружили, что если поместить вытяжку из мышц в гликоген, то никакого расщепления происходить не будет, пока в раствор не будет добавлен фосфат-ион. Естественно, первым делом исследователи предположили, что продукт распада должен будет вступить в соединение с фосфат-ионом, стали искать и в конце концов нашли в смеси сахарофосфат (не глюкозу!). В 1937 году они установили, что этот сахарофосфат, получивший к тому времени название «эфир Кори», является «глюкозо-1-фосфатом». (Шесть атомов углерода, входящих в состав глюкозы, имеют, согласно международной договоренности, номера от 1 до 6, и глюкозо-1-фосфат — это молекула, в которой фосфатная группа крепится к атому углерода № 1.)
Так оказалось, что гликоген в организме подвергается не гидролизу, а распаду с использованием фосфорной кислоты. Я поясню, что я имею в виду, на примере прилагаемой диаграммы. Пусть G — глюкоза, тогда гликоген — это цепочка G3, соединенная кислородными связями.
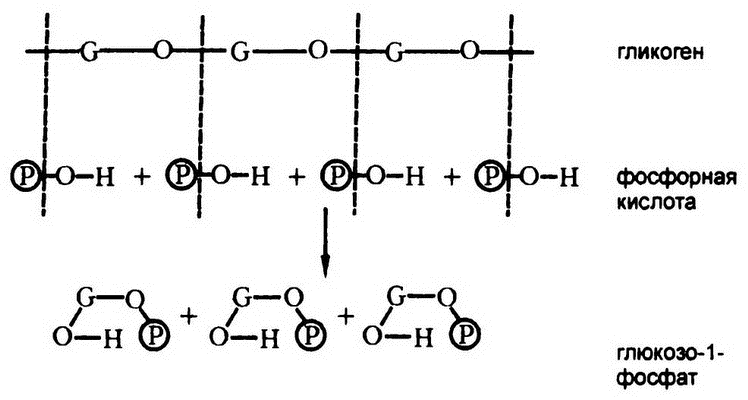
Рис. 48. Фосфоролиз гликогена
Длинная глюкозная цепочка гликогена расщепляется на одиночные молекулы сахара путем добавления наперерез связям элементов фосфорной кислоты. Если бы добавляемые элементы были молекулами воды, то речь шла бы о гидролизе. Раз вместо воды используется фосфорная кислота, то и реакцию эту естественно назвать «фосфоролизом» (рис. 48). Таким образом, гликоген в живых тканях расщепляется на глюкозо-1-фосфаты с помощью фосфоролиза.
Разница между этими двумя типами реакций имеет отношение к свободной энергии. Гидролиз гликогена на глюкозу подразумевает значительное уменьшение уровня свободной энергии, а для того, чтобы подобным же образом снова создать гликоген из глюкозы, потребуется такое же количество энергии приложить на каждое заново формируемое звено связи.
Фосфоролиз же гликогена на глюкозо-1-фосфат подразумевает крайне малое снижение уровня свободной энергии. Гликоген и глюкозо-1-фосфат находится почти на одинаковом энергетическом уровне. Соответственно, реакция фосфоролиза легко обратима, так что при равных концентрациях организм может легко переключаться между расщеплением и построением гликогена, в зависимости от потребностей, поддерживая таким образом необходимую для здоровья гармонию, что уже упоминалось в главе 19.
Однако производство глюкозо-1-фосфата всей полноты проблемы не снимает. Прежде всего, сахарофосфаты по какой-то причине не могут проникать через клеточную мембрану, так что любое соединение, содержащее глюкозо-1-фосфат, будучи созданным в клетке печени, обречено в ней и оставаться, пока не утратит фосфат. Поэтому при пищеварении образуется именно глюкоза, поскольку только она способна преодолеть кишечные мембраны и попасть в клетки тканей — глюкозо-1-фосфат не смог бы. И только глюкоза может присутствовать в крови, поскольку только она, а не глюкозо-1-фосфат, может проникать из крови в клетки и питать их. Поэтому именно глюкозу, а не глюкозо-1-фосфат, клетки печени должны выбрасывать в кровь.
Так и происходит. В клетках печени был обнаружен фермент, катализирующий превращение глюкозо-1-фосфата в глюкозо-6-фосфат (фосфатная группа при этом смещается от углерода-1 к углеро-ду-6, что не требует каких-либо заслуживающих упоминания перемен в энергии), а затем был обнаружен и второй фермент, действие которого заключается в отсечении фосфатной группы посредством гидролиза. Этот шаг высвобождает глюкозу, которая просачивается сквозь клеточную мембрану и попадает в кровоток.
Но обойти требования, касающиеся свободной энергии, невозможно. При переходе от глюкозо-6-фосфата к непосредственно глюкозе происходит ее высвобождение. То же самое произошло бы и при расщеплении гликогена на глюкозу напрямую; так не все равно — расщеплять гликоген напрямую или через посредническую цепочку из двух сахарофосфатов? Чем это лучше? Тогда перед тем, как образовывать гликоген из глюкозофосфатов, для образования глюкозофосфатов из глюкозы организму все равно придется где-то брать энергию — так не проще ли сразу образовывать его из глюкозы, минуя промежуточные стадии?
Нет. При формировании гликогена перед организмом встают две основные проблемы: надо создать большую и сложную молекулу из простых и маленьких и надо найти энергию для соединения каждой из связей в цепочке, каковых в каждой молекуле гликогена по тысяче. Если организм будет поставлен перед задачей создать гликоген сразу из глюкозы, ему придется решать обе эти проблемы одновременно, и очевидно, что организм с ними не справляется.
Если же сначала глюкоза преобразуется в глюкозо-6-фосфат, то энергию искать все равно приходится, но первая проблема при этом снимается — простое вещество переходит в другое, почти столь же простое. Затем, когда глюкозофосфат уже сформирован, организму остается только объединить единицы глюкозы в огромную молекулу, но энергии для этого уже почти не требуется. Таким образом, одна сложная задача для организма разбивается на две простые, и с ними поодиночке организм вполне справляется.
Описываемый баланс гликогена и глюкозы проиллюстрирован на рис. 49.
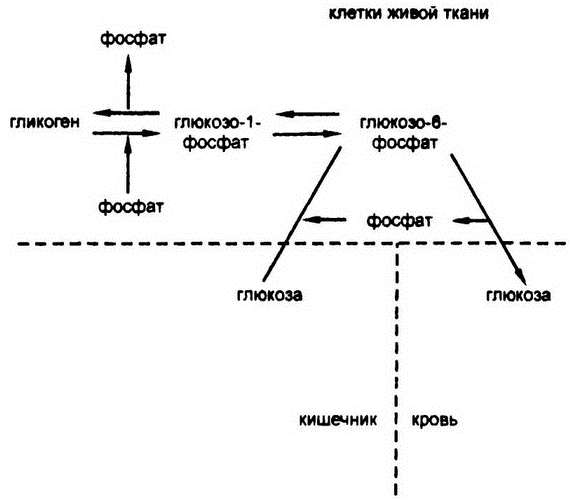
Рис. 49. Связь гликогена и глюкозы
Следующий вопрос таков: так или иначе, при переходе от глюкозы к гликогену организму все же необходимо найти-то источник энергии. Где же он ее берет? Иными словами, глядя на рис. 49, мы можем спросить: за счет чего организм переводит глюкозу в глюкозо-6-фосфат?
Вот здесь и проявляется роль высокоэнергетической фосфатной связи. Для перехода от глюкозы к глюкозо-6-фосфату требуются две вещи — фосфатная группа и энергия. АТФ обладает и тем и другим.
Реакцию глюкозы с АТФ можно выразить следующим образом:
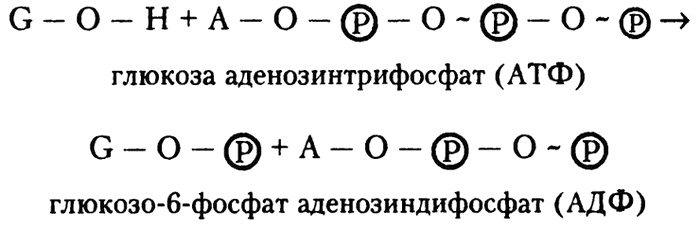
Представим себе, что это выражение представляет собой комбинацию двух реакций. Первая — это перевод глюкозы в глюкозо-6-фосфат. При этом формируется низкоэнергетическая фосфатная связь, для чего требуется затратить около 4 килокалорий на моль вещества. Вторая — перевод АТФ в АДФ путем гидролиза высокоэнергетической фосфатной связи, в процессе чего высвобождается около 8 килокалорий на моль вещества. Получается, что гидролиза АТФ более чем достаточно для того, чтобы глюкоза могла превратиться в глюкозо-6-фосфат.
Это пример специфической парной реакции, при одной части которой высвобождается больше энергии, чем при второй — тратится, так что в итоге энергия все равно высвобождается (второй закон термодинамики не обойдешь).
До последнего времени существовало мнение, что АТФ — это и есть энергетический концентрат организма, и везде, где требуется проведение энергоемкой реакции, подключается АТФ. Однако с помощью современных методик анализа ткани было установлено, что существуют и другие трифосфаты, и теперь считается, что для различных реакций существуют и различные источники энергии. Тем не менее все они действуют по тому же принципу, и то, что я рассказал об АТФ, в равной степени относится и к близким к нему веществам.
Фруктоза и галактоза, два других простых сахара, впитываемые организмом, тоже образуют соединения при энергетической помощи АТФ (и особых ферментов, разумеется, не тех, которые действуют в отношении глюкозы). Фруктозо-1-фосфат и галактозо-1-фосфат превращаются в тот же глюкозо-1-фосфат с помощью реакций, не требующих заслуживающей упоминания энергии, а дальше фруктоза и галактоза обычным путем превращаются в гликоген.
Формирование гликогена из простых Сахаров, таким образом, «называется «гликогенезом», а расщепление гликогена на простые сахара — «гликогенолизом». Схема с участием АТФ и всех трех Сахаров представлена на рис. 50.
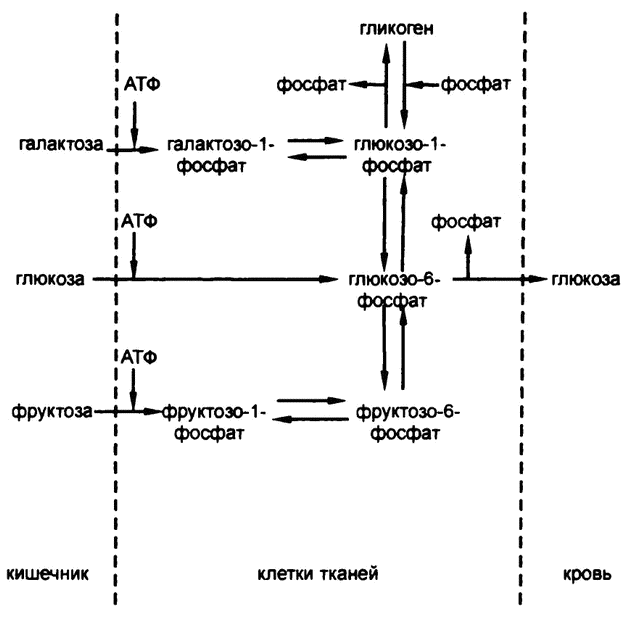
Рис. 50. АТФ и гликоген
Но что-то слишком гладко все получается. И правда, не можем же мы утверждать, что энергетический запас АТФ и подобных ему молекул — ответ на все задачи жизни? Что же происходит, когда он исчерпывается?
Естественно, он не исчерпывается, потому что если бы он исчерпался, то анаболизм прекратился бы, а вместе с ним — и жизнь. Поскольку жизнь не прекращается, значит, запас энергии каким-то образом возобновляется так же быстро, как и тратится.
Это означает, что организм каким-то образом строит АТФ из АДФ с той же скоростью, с какой АТФ расщепляется до АДФ в ходе множества парных реакций. Кажется, мы совсем запутались. Решив проблему энергоемких (4 килокалории на моль) процессов соединения глюкозы за счет гидролиза АТФ до АДФ, теперь мы должны решить проблему еще более энергоемких (8 килокалорий на моль) процессов создания АДФ из АТФ?
Что ж, они происходят в ходе анаэробного гликолиза, который я описывал в предыдущей главе.
На протяжении 30-х годов XX века биохимики выделяли промежуточные вещества реакций гликолизации дрожжей или живых тканей и искали ферменты, катализирующие реакции с участием этих промежуточных веществ. Выделение промежуточных веществ и изучение реакций позволило им шаг за шагом собрать, как мозаику, всю цепь катаболизма от глюкозы до молочной кислоты.
Сначала к глюкозе присоединяется фосфатная группа, затем, после перехода во фруктозу, — вторая. (Образованный таким образом фруктозодифосфат — это и есть тот самый «эфир Хардена—Янга», который я упоминал в прошлой главе.) Прикрепление каждого фосфата требует участия АТФ, так что может показаться, что цепочка гликолиза лишь усугубляет проблему, используя АТФ для своих целей, вместо того чтобы его создавать. Но мы ведь еще не закончили.
Фруктозодифосфат со своими шестью атомами углерода расщепляется на две половинки по три атома углерода в каждой (глицеральдегид-3-фосфата), в каждой из которых оказывается по одному фосфату. Что происходит с этим веществом дальше, показано на рис. 51.
Как видите, к нему добавляется вторая фосфатная группа, но не методом обычной конденсации. То есть в ходе этой реакции не образуется вода (Н — О — Н). Вместо этого выделяются два атома водорода — и все. Такой процесс называется дегидрогенизацией. При этом два выделяющихся атома водорода не образуют молекулы газообразного водорода, хотя я и записал их в выражении как просто 2Н. Их дальнейшая судьба немного сложнее, и мы вернемся к ним позже.
В результате дегидрогенизации образуется ацилфосфат, который, как я уже упоминал, является высокоэнергетическим соединением. Откуда же берется энергия для его образования? Из потери двух атомов водорода. Дегидрогенизация подразумевает снижение уровня свободной энергии на 35—70 килокалорий на моль вещества, и этого более чем достаточно, чтобы получить необходимые 8 килокалорий для формирования высокоэнергетической связи. (В строго анаэробных условиях такого масштабного пустого расходования энергии, что нам приходит в голову при виде приведенных цифр, не получается, потому что оба атома водорода в итоге потом возвращаются — вместе со своей энергией.)
Далее в цепочке формируется вторая высокоэнергетическая связь в ходе реакции, менее энергичной, чем дегидрогенизация, но тем не менее достаточной для превращения уже существующей низкоэнергетической связи в высокоэнергетическую.
Любое вещество, в состав которого входит высокоэнергетическая фосфатная связь (например, 1,3-дифосфоглицериновая кислота), может вступать в реакцию с АДФ для передачи ему этой фосфатной связи, образуя АТФ. На такой перенос связи никаких заслуживающих упоминания энергозатрат не требуется. Таким образом, формирование любой высокоэнергетической фосфатной связи равносильно формированию молекулы АТФ.
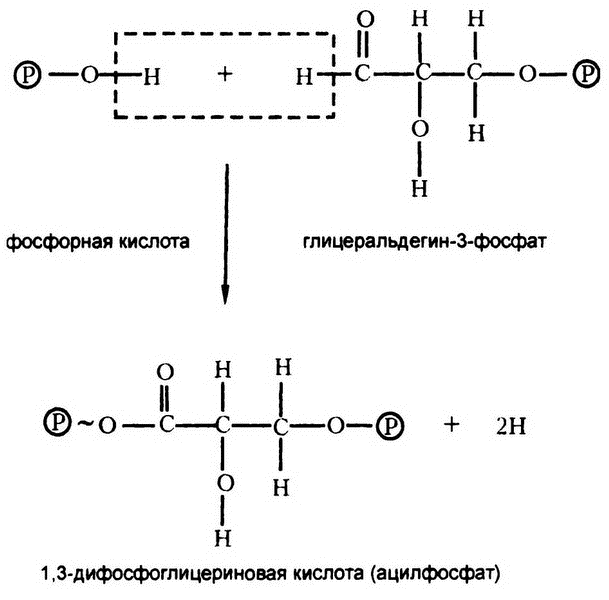
Рис. 51. Дегидрогенация
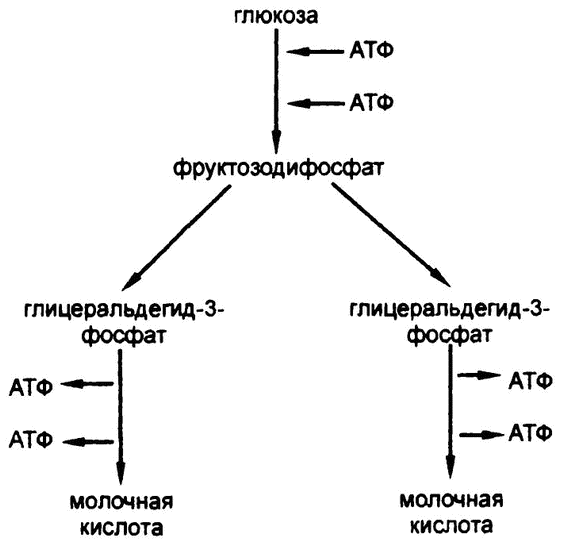
Рис. 52. Схема гликолиза
Итак, подытожим. При анаэробном гликолизе каждая молекула глюкозы начинает свое превращение в молочную кислоту с помощью двух молекул АТФ. Однако в конечном итоге глюкоза распадается на соединения с тремя атомами углерода, каждый из которых к моменту окончательного превращения в молочную кислоту доставляет в организм две молекулы АТФ.
Следовательно, шесть атомов углерода, содержащихся в глюкозе, используя две молекулы АТФ, создают их четыре, таким образом принося организму доход в две молекулы АТФ (рис. 52), которые потом можно использовать в парных реакциях для осуществления процессов анаболизма.
Но, как я уже говорил, гликолиз — неэффективный способ производства энергии. Если бы организм жил на одном гликолизе, он вел бы такое же тупое существование, как и дрожжи. Нас интересует большее, поэтому мы сейчас обратимся к кислороду.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 3 Наша потребность в животных и ее роль для «культурной лестницы». Ручные животные и их значение. Новое положение женщины — снижение ее статуса.
Глава 3 Наша потребность в животных и ее роль для «культурной лестницы». Ручные животные и их значение. Новое положение женщины — снижение ее статуса. Какая же, по моему мнению, черта нашего биологического вида направила его по новому пути и привела к чрезвычайно
Глава 4 Наше самое важное ручное животное и его значение для возникновения земледелия. Примечательное развитие хлебных злаков.
Глава 4 Наше самое важное ручное животное и его значение для возникновения земледелия. Примечательное развитие хлебных злаков. Ответить на вопрос, когда человек впервые приручил крупный рогатый скот, так же трудно, как на вопрос о времени одомашнивания других животных.
Глава 17 Гибкое тело человека. Особые свойства человеческого мозга. Значение образного мышления. Пещерные рисунки. «Сверхтучная» женщина. Был ли кроманьонец обнаженным? Появление религии. Шаманы и служители божьи.
Глава 17 Гибкое тело человека. Особые свойства человеческого мозга. Значение образного мышления. Пещерные рисунки. «Сверхтучная» женщина. Был ли кроманьонец обнаженным? Появление религии. Шаманы и служители божьи. Ясно, что наш вид Homo sapiens sapiens неизмеримо превосходил все
Глава 1.1. Значение техники искусственного выкармливания новорожденных хищных млекопитающих для формирования поведения взрослых
Глава 1.1. Значение техники искусственного выкармливания новорожденных хищных млекопитающих для формирования поведения взрослых Поведенческие реакции, сформированные к моменту рождения незрелорождающихся животных и способствующие их первому контакту с окружающей
8 Значение культуры
8 Значение культуры Эпиграф к этой главе — переложение мифа о сотворении, взятое из книги Артура Коттерелла «Энциклопедия мировой мифологии» (Arthur Cotterell. Encyclopedia of World Mythology. — Paragon, Bath, 1999).Сокращенную версию бортового журнала Кука с корабля «Резолюшн» можно найти в книге
Глава 1. История и значение генетики
Глава 1. История и значение генетики Генетика – это сердцевина биологической науки. Лишь в рамках генетики разнообразие жизненных форм и процессов может быть осмыслено как единое целое. Ф. Айала, американский генетик, автор учебника «Современная генетика» Генетика
Регуляция обмена ионов кальция и фосфатов
Регуляция обмена ионов кальция и фосфатов Кальций и фосфаты являются структурными компонентами костной ткани. Ионы кальция участвуют в свертывании крови, мышечном сокращении, проведении нервного импульса, влияют на работу ионных насосов, способствуют секреции
Глава 1 Размер имеет значение
Глава 1 Размер имеет значение Сказал я девушке кротко: — Простите за нетактичность, Но бюст ваш, и торс, и походка Напомнили мне античность. Она в ответ мне со вздохом: — Простите, но ваше сложение Напомнило мне эпоху Упадка и разложения. Олег Григорьев Любовь и смерть —
Глава XI. БИ, ГОМО, ГЕТЕРО: ЭТО, ВООБЩЕ, ИМЕЕТ ЗНАЧЕНИЕ?
Глава XI. БИ, ГОМО, ГЕТЕРО: ЭТО, ВООБЩЕ, ИМЕЕТ ЗНАЧЕНИЕ? ЯЛВ: Твои исследования привели к удивительному выводу: у женщин бывает больше гомосексуальных эпизодов, чем у мужчин. Можешь сказать, почему так?ЗИ: У женщин бывают разные мотивы для гомосексуальных эпизодов. Например,
Глава 1 Значение и измерение интеллекта
Глава 1 Значение и измерение интеллекта 1. Определение интеллекта 2. Иерархическая модель интеллекта 3. Коэффициент интеллекта (IQ) 4. Эффект Флинна Расовые различия в интеллекте начали анализироваться с научной точки зрения с
Глава 2 Значение и формирование рас
Глава 2 Значение и формирование рас 1. Формирование рас, разновидностей и пород 2. Разновидности в животном мире 3. Таксономии рас 4. Расовые различия в заболеваемости 5. Существуют ли расы? Имея дело с расовыми
Глава 5. Экологическое значение геофизических электромагнитных полей
Глава 5. Экологическое значение геофизических электромагнитных полей Медицинская сторона экологического значения естественных ЭМП прежде всего находит выражение в корреляции между изменениями ЭМП и обострением различных заболеваний, хотя здоровые люди тоже
Глава 11. Размер имеет значение Как люди получили такой большой мозг?
Глава 11. Размер имеет значение Как люди получили такой большой мозг? Для того чтобы расселиться по всей планете, нашим предкам должно было понадобиться что-то большее, чем настойчивость и удача. Чтобы раз за разом избегать вымирания, нужны были еще и мозги. Человеческий
Глава 11: Размер имеет значение
Глава 11: Размер имеет значение Gould, Stephen Jay. Wide Hats and Narrow Minds // The Panda’s T umb. W. W. Norton, 1980.* Исключительно увлекательная трактовка истории об аутопсии Кювье.Isaacson, Walter. Einstein: His Life and Universe. Simon and Schuster, 2007.Jerison, Harry. On Theory in Comparative Psychology // The Evolution of Intelligence. Psychology Press, 2001.Tre f ert, D., Christensen D. Inside the