Попытки стать приматом – плезиадапиформы
Плезиадапиформы (отряд Plesiadapiformes или подотряд Plesiadapiformes отряда Primates) известны из палеоцена и эоцена Европы, Северной Америки и Азии (65–42 млн лет назад). Обликом и образом жизни они были во многом похожи на грызунов. Размеры с мышь или крысу, вытянутая мордочка, маленький и просто устроенный мозг, глаза, расположенные по бокам головы, короткие массивные лапы, длинный хвост – все это общие признаки всех некрупных млекопитающих. Плезиадапиформы, с одной стороны, в общем плане строения имеют много от приматов, с другой – отличаются рядом параметров. Например, плезиадапиформы имели когти на всех пальцах, по крайней мере у части видов отсутствовало окостенение слуховой капсулы, многие обладали специализациями в зубной системе. Благодаря отпечаткам из французского местонахождения Мена известно, что плезиадаписы владели пушистым хвостом – не слишком характерным для приматов.
При этом из палеоцена и эоцена неизвестны другие млекопитающие, которые могли бы быть предками приматов. Посему включение плезиадапиформов в приматов или исключение из этого отряда – вопрос совести, а не таксономии. В настоящее время их все же чаще выделяют в самостоятельный отряд Plesiadapiformes, но близость к приматам подчеркивается объединением тех и других в рамках надотрядной группировки Primatomorpha.
Несмотря на древность, известно весьма много разных плезиадаписовых, в основном по зубам, но есть находки и целых скелетов. Замечательно, что даже на ранних этапах эволюции они были чрезвычайно многообразны. Это тем более так, поскольку мы знаем лишь о малой доле из общего числа столь древних животных. Густой букет родов и видов, возникший чуть ли не мгновенно из единого предка, называется первой радиацией приматоморфов.
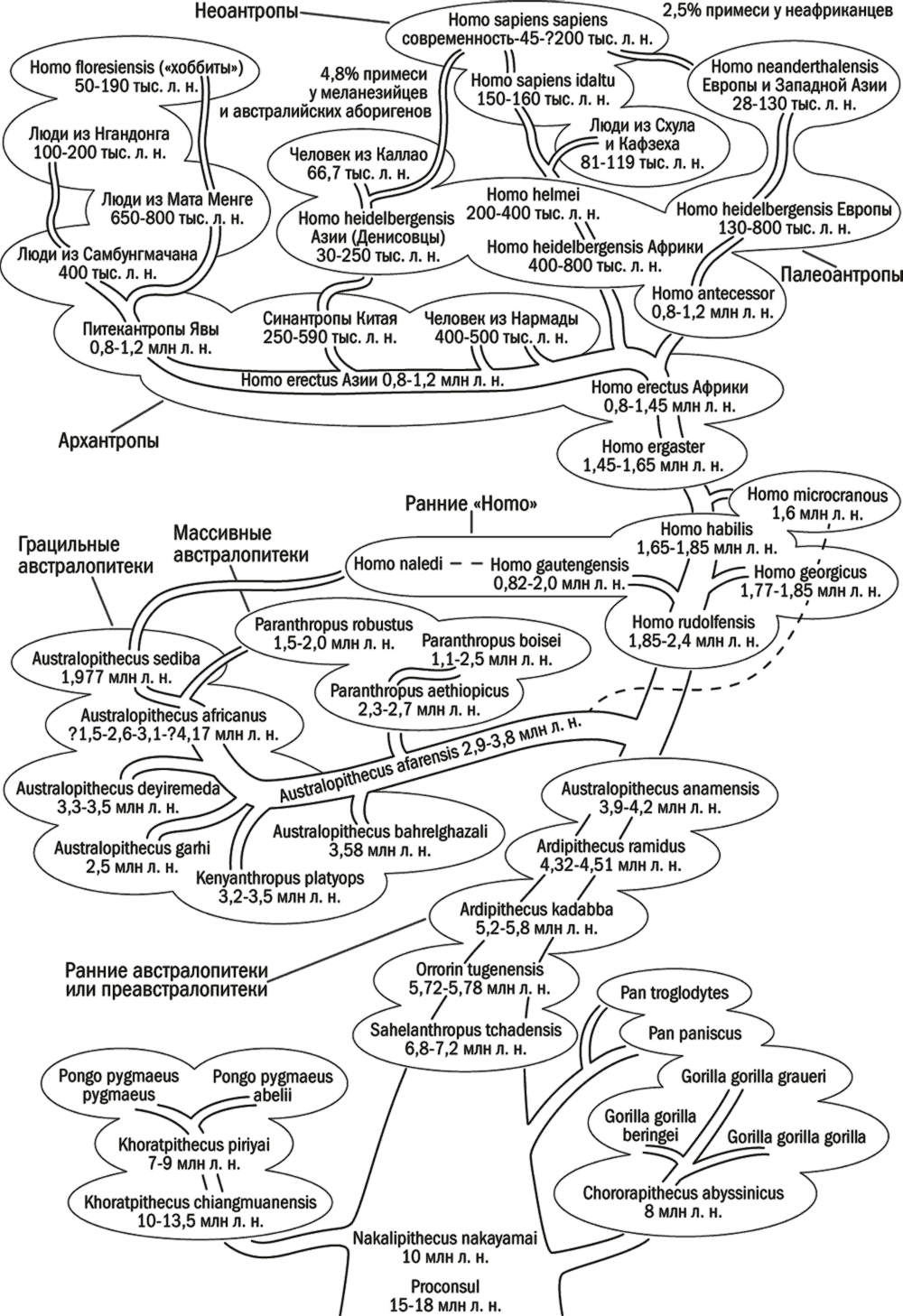
Рис. 26. Схема эволюции приматов
Относительно типа передвижения и образа жизни плезиадапиформов велись споры. Одни считали их наземными (Gingerich, 1976, 1984), другие – древесными (Szalay et Delson, 1979), однако ж отсутствие срастаний костей, гибкость конечностей и пропорции кисти однозначно свидетельствуют в пользу второго варианта. Впрочем, древесность древесности рознь: плезиадапиформы не умели бегать и скакать по веткам так же резво, как делают это сейчас обезьяны. Может, потому они и приобретали специализации зубов, подобные грызуньим, – при пониженной мобильности надо извлекать из доступных ресурсов максимум выгоды? Неспроста зубная система, аналогичная системе многобугорчатых – с резко выступающими резцами, крупным режущим премоляром и перетирающими или давящими молярами, – возникала неоднократно среди примитивных приматоподобных существ: у саксонеллид Saxonellidae, карполестоидов Carpolestoidea и фенаколемуриновых Phenacolemurinae. Наблюдая, сколь неряшливо едят обезьяны – без конца роняя что-то, выедая лишь самые вкусные кусочки и оставляя массу объедков, – можно задуматься. Обезьяны всегда могут быстро переместиться и найти новые вкусняшки, а медленные плезиадапиформы, грызуны и многобугорчатые вынуждены довольствоваться тем, что есть на месте, и грызть до победы.
Уголок занудства
Тип питания определяется по форме бугорков на молярах: высокие и острые типичны для насекомоядных животных, низкие и тупые – для растительноядных. Можно посчитать длину режущих гребешков эмали относительно длины самого зуба: у насекомоядных и листоядных гребешки будут относительно длинные (ведь надо пережевывать хитин или целлюлозу), у фруктоядных намного меньше, а у любителей нектара и древесных смол – совсем короткие. У насекомоядных эмаль обычно более гладкая, а у растительноядных – морщинистая.
Еще точнее можно определять диету по микростертости эмали: тут учитывается регулярность или беспорядочность царапин, их частота, направление, глубина и форма краев, микросколы, видные под электронным микроскопом (например: Calandra et al., 2012; Scott et al., 2012). Для анализа этих особенностей используются фракталы – вот зачем будущим биологам могут понадобиться уроки геометрии в школе!
У листоядов обычно очень крупные челюсти, так как листья малопитательны, их надо есть очень много; на углу нижней челюсти листоядов обычно развит большой дополнительный отросток.
Несмотря на внешнее сходство, зубная система специализированных плезиадапиформов не была идентична таковой грызунов. У представителей семейства плезиадапид Plesiadapidae верхние резцы имели бугорки и не были самозатачивающимися. У многих полностью исчезли нижние клыки и вторые нижние резцы, а у некоторых и верхние клыки, так что между резцами и премолярами имелся большой зазор – диастема. От ранних форм к поздним доля мягкой пищи – фруктов и особенно листьев – в рационе, вероятно, увеличивалась (Boyer et al., 2010a). Плезиадапиды были одними из самых успешных животных своего времени, в некоторых местонахождениях они встречаются чаще всех остальных млекопитающих. Это отразилось и в распространении: в Европе, Северной Америке, Пакистане и Китае с нижнего палеоцена по нижний эоцен. Размеры их сильно варьировали – самые мелкие были как белка, а некоторые были довольно крупными – размером с крупную кошку.
Не слишком родственны, но экологически схожи с плезиадапидами микросиопиды Microsyopidae, жившие с верхнего палеоцена по средний эоцен в Северной Америке; среди них встречались животные размером с бобра. Относительно микросиопидов, впрочем, есть сомнения: некоторые специалисты считают их грызунами, насекомоядными или группой в отряде шерстокрылов.
Паромомиоиды Paromomyoidea, включающие паромомиид Paromomyidae, одними исследователями считаются шерстокрылами, а другими – плезиадапиформами. В частности, род Ignacius на основании строения зубов относится к фенаколемуринам Phenacolemurinae – подгруппе паромомиид, типовой род которых Phenacolemur имел пальцы, почти неотличимые от пальцев современного шерстокрыла (Beard, 1990, 1993). Да и у самого Ignacius форма костей кисти тоже весьма шерстокрылья (Hamrick et al., 1999; Krause, 1991). Посему, пока были известны только изолированные кости конечностей, предполагалось, что эти существа имели летательную перепонку и были способны к планирующему полету. Однако обнаружение целых скелетов показало, что эти звери были больше похожи на белок, а не на летяг. Пропорции кисти к туловищу, строение основания черепа и внутреннего уха у Ignacius скорее как у современных обезьян (Bloch et Silcox, 2001; Bloch et al., 2007; Silcox, 2003). Такая мозаика признаков, запутывающая общую картину, типична для древнейших приматов и близких им групп. Паромомиоиды имели квадратные уплощенные моляры, приспособленные для перетирания фруктов, впрочем, в диете немалую роль могли играть древесный сок и насекомые. В последовательном ряду палеоценовых родов наблюдается быстрое удлинение и истончение нижних резцов с появлением промежутка – диастемы – между резцами и премолярами.
Карполестоиды Carpolestoidea почти полностью перешли на питание волокнистыми плодами, орехами и стеблями, благодаря чему приобрели плоские и высокие премоляры с пильчатым краем. У них имелись и продвинутые приматные черты: например, большой палец ноги был снабжен ногтем, а не когтем, что подразумевает и б?льшую чувствительность; впрочем, и карполестоиды были медлительными животными и не могли быстро прыгать по ветвям. Они существовали не так уж долго – в пределах среднего и верхнего палеоцена Северной Америки, а последние представители доживали свой век в нижнеэоценовых лесах Китая и среднеоэценовых – Пакистана. Карполестоиды никогда не были многочисленны, и исчезновение их может быть связано с климатическими колебаниями или конкуренцией с более крупными животными, поскольку карполестоиды были одними из самых небольших плезиадапиформов – размером с мышь или крысу.
Пикродонтиды Picrodontidae, с их морщинистыми многобугорковыми, очень широкими – особенно первыми – и совсем не стертыми молярами, стали очень специализированными фруктоядами, а может, даже питались преимущественно нектаром, пыльцой и древесным соком. Череп Zanycteris резко сужался спереди, этим пикродонтиды были похожи на современных мышиных лемуров Cheirogaleinae и хоботноголовых кускусов Tarsipedidae, питающихся нектаром. Пикродонтиды, видимо, дальше всех из плезиадапиформов ушли от исходной насекомоядности. Забавно, что зубы пикродонтид столь необычайно похожи на зубы летучих мышей, что в 1935 году Дж. Симпсон даже предполагал, что пикродонтиды относятся к рукокрылым (Simpson, 1935).
Все эти специализированные плезиадапиформы, конечно, не были предками приматов, но известны и более генерализованные их варианты, например средне-верхнепалеоценовые североамериканские палехтониды Palaechthonidae. У них сохранялась исходная для приматов зубная формула, не было диастемы между резцами и премолярами, отсутствовали гипертрофии каких-либо зубов. Строение черепа Palaechton nacimienti было самым примитивным среди всех плезиадапиформов: глазницы маленькие, ориентированы вбок и широко расставлены, заглазничное сужение не выражено, обонятельные луковицы большие; судя по увеличенному подглазничному отверстию, имелись длинные вибриссы. Ориентировались палехтониды в основном с помощью осязания, слуха и обоняния, а не зрения. Судя по всему, вели они в основном наземный образ жизни. В немалой степени все эти черты были связаны с малыми размерами тела и насекомоядностью палехтонид (Kay et Cartmill, 1977). Показательно, что группа палехтонид – вовсе не древнейшая среди плезиадапиформов; с одной стороны, она сохраняла исходную насекомоядность и экологически схожа с тупайями, с другой – по времени близка к настоящим приматам границы палеоцена и эоцена. Вряд ли настоящих приматов дали палехтониды с их особенностями, но истинными предками должны были быть очень похожие на них существа, только более древесные и всеядные.

Рис. 27. Черепа Plesiadapis (а) и Palaechton nacimienti (б).
Насекомоядность вполне сочеталась с древесностью у микромомиид Micromomyidae, которые были одними из самых мелких плезиадапиформов – весом 20–30 г (самыми лилипутскими были представители подсемейства тинимомиин Tinimomyinae), вдвое меньше современных мышиных лемуров. При их размере другая диета, нежели насекомые, млекопитающим резко противопоказана. Эти крошечные зверюшки жили в конце палеоцена и начале эоцена в Северной Америке.
Впрочем, и размеры микромомиид не предельны: поздние пикромомииды Picromomyidae побили все рекорды – Picromomys petersonorum весил ничтожные 10 г. Эти животные известны из среднего эоцена Северной Америки.
Позднейшей группой насекомоядных плезиадапиформов были толиапиниды Toliapinidae (Hooker et al., 1999). Они появились лишь в конце палеоцена и дожили до среднего эоцена. Большинство видов, включая примитивнейшего Sarnacius gingerichi, найдено в Европе, но Altiatlasius koulchii мигрировал в Марокко, а самый поздний – Seia shahi – обнаружен в Пакистане. Altiatlasius koulchii первоначально был описан как древнейший настоящий примат семейства Omomyidae, потом считался даже антропоидом-эосимидом (это было сенсацией, так как получалось, что человекоподобные приматы появились уже в конце палеоцена, 57–60 млн лет назад!), но позже пересмотрен как плезиадапиформ. Толиапиниды были весьма схожи с микромомиидами по образу жизни и являются их полным “старосветским” аналогом; к сожалению, от тех и других найдено очень немного останков.
Среди плезиадапиформов особняком стоит семейство Adapisoriculidae. Особенности его представителей позволяли разным исследователям включать его в сумчатых, насекомоядных, лептиктид или миксодектид, а также считать предковым для тупай. Адаписорикулиды назывались и как возможные предки афросорицид Afrosoricida, включающих современных златокротов Chrysochloridae и тенреков Tenrecidae (Seiffert, 2010). Несмотря на существенные различия с плезиадаписовыми в зубной системе, строение посткраниального скелета (то есть всего скелета, кроме черепа) у плезиадапиформов, шерстокрыловых и адаписорикулид весьма схоже (Smith et al., 2010); судя по всему, это были древесные насекомоядные животные. Более того, плечевая кость адаписорикулид имеет промежуточное строение между вариантами меловых “кондиляртр” и кайнозойских эуархонт, причем в ряду самих адаписорикулид обнаруживается постепенный переход от более примитивного варианта мелового Deccanolestes к более эуархонтовому у палеогеновых форм (Boyer et al., 2010b). Если считать адаписорикулид приматами или приматоморфами, а Deccanolestes hislopi – адаписорикулидом, то эта группа становится самой древней среди приматоморфов, поскольку указанный вид обнаружен в позднемеловых отложениях Индии. Учитывая, что Индия в то время была островом, удаленным от всех других земель, а Purgatorius в то же время жил в Северной Америке, пути миграций становятся совсем непонятными.
Замечательно, что адаписорикулиды найдены в нижнем и верхнем палеоцене и нижнем эоцене Европы, а также верхнем палеоцене и нижнем эоцене Северной Африки, но ни малейших их следов нет в хорошо изученных фаунах Северной Америки и главной части Азии. Таким образом, либо адаписорикулиды были до крайности приматоподобными сумчатыми или насекомоядными, либо приматы зародились не в Северной Америке. Сами адаписорикулиды могли возникнуть в разных местах: древнейшие представители найдены в верхнем мелу Индии (несколько видов одного или двух родов, отсюда они могли попасть в Европу через Восточную Африку или острова моря Тетис), но самое большое разнообразие обнаруживается в нижнем палеоцене Европы (они явно появились тут раньше, но меловые европейские млекопитающие известны плохо, так что в Индию и Африку могли попасть отсюда), а примитивнейший вид Afrodon chleuhi известен из верхнего палеоцена Марокко (хотя более ранних в Африке пока не нашли, потенциально они могли распространиться отсюда и в Индию, и в Европу; Smith et al., 2010; De Bast et al., 2012). Не исключено, что предки адаписорикулид жили в Европе и оттуда попали в Северную Америку, только там стали пургаториусами, а позже – плезиадапиформами. С другой стороны, адаписорикулиды могут быть предками современных шерстокрылов; из Индии они могли попасть в Юго-Восточную Азию, когда эти две части суши соединились (Smith et al., 2010). С третьей стороны, даже отнесение рода Deccanolestes к плацентарным вызывает сомнения; согласно некоторым кладистическим расчетам, он вместе с прочими адаписорикулидами может представлять очень архаичную линию (намного примитивнее, чем Cimolestes, Zalambdalestes и Purgatorius), восходящую к корням всех эутериев, поздние представители которой сохранили примитивные черты до весьма поздних времен (Goswami et al., 2011). Впрочем, адекватность результатов кластерного анализа, применявшегося в подобных исследованиях по выявлению филогении, мягко говоря, спорна, что можно видеть, например, при сравнении выводов двух групп палеонтологов: в одной работе адаписорикулиды оказались предками тенреков (Seiffert, 2010), во второй – ответвлением древнейших и примитивнейших эутериев, даже близко не родственных ни тенрекам, ни эуархонтам (Goswami et al., 2011). Как бы то ни было, существование адаписорикулид свидетельствует, во-первых, о возможности межконтинентальных обменов фаун в позднем мелу и раннем палеогене, а во-вторых, о существовании примитивных групп, близких к предкам приматов, на осколках Гондваны – в Африке и Индии.
Специфические признаки и особенности являются главным препятствием для признания плезиадапиформов в качестве настоящих приматов. Ведь они обладали маленьким мозгом (гораздо меньшим, чем у приматов таких же размеров, хотя и большим, чем у других зверей) с хорошо развитыми обонятельными центрами; зрение, судя по направленным в стороны глазам, не было стереоскопичным; имелись когти на пальцах, их большой палец не противопоставлялся, а кисть была больше “цеплятельной”, нежели “хватательной”. Поэтому, несмотря на тот факт, что плезиадапиформы уже ушли от сугубой насекомоядности, а некоторые стали специализированными листо– и фруктоядами, многие систематики склонны считать первыми приматами только эоценовых адапиформов Adapiformes и омомиформов Omomyiformes.
Имевшиеся же прогрессивные особенности плезиадапиформов не уберегли их от вымирания. Традиционно и по умолчанию считалось, что в их исчезновении повинны более продвинутые настоящие приматы. Однако изучение частот встречаемости останков рисует другую картину (Fleagle, 1999). Резкий спад численности плезиадапиформов начинается совершенно синхронно со столь же резким подъемом численности первых грызунов, тогда как лишь через некоторое время появляются и начинают распространяться полуобезьяны. Стало быть, плезиадапиформы проиграли эволюционную гонку грызунам, зато своим исчезновением освободили дорогу полуобезьянам; спасибо мышам – без них нас бы не было! Таким образом, отличия плезиадапиформов от приматов оказываются крайне существенными: то, чего не было у первых, не позволило им успешно конкурировать с грызунами; то же, что приобрели вторые, вывело их на новый уровень, недоступный грызунам.
Дальнейшее развитие группы привело к первой радиации приматов – возникновению множества новых видов, давших начало разным линиям приматов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК