Долгопятообразные Tarsiiformes
Внижнем эоцене из какой-то группы полуобезьян выделилась линия современных обезьян, но почти сразу она разделилась на долгопятообразных и антропоидов. Поэтому долгопятообразные имеют едва ли не большинство признаков полуобезьян. По этой же причине раньше их включали именно в полуобезьян, потом стали выделять в самостоятельную промежуточную группу, а ныне рассматривают как специализированных и во многом примитивных, но все же обезьян.
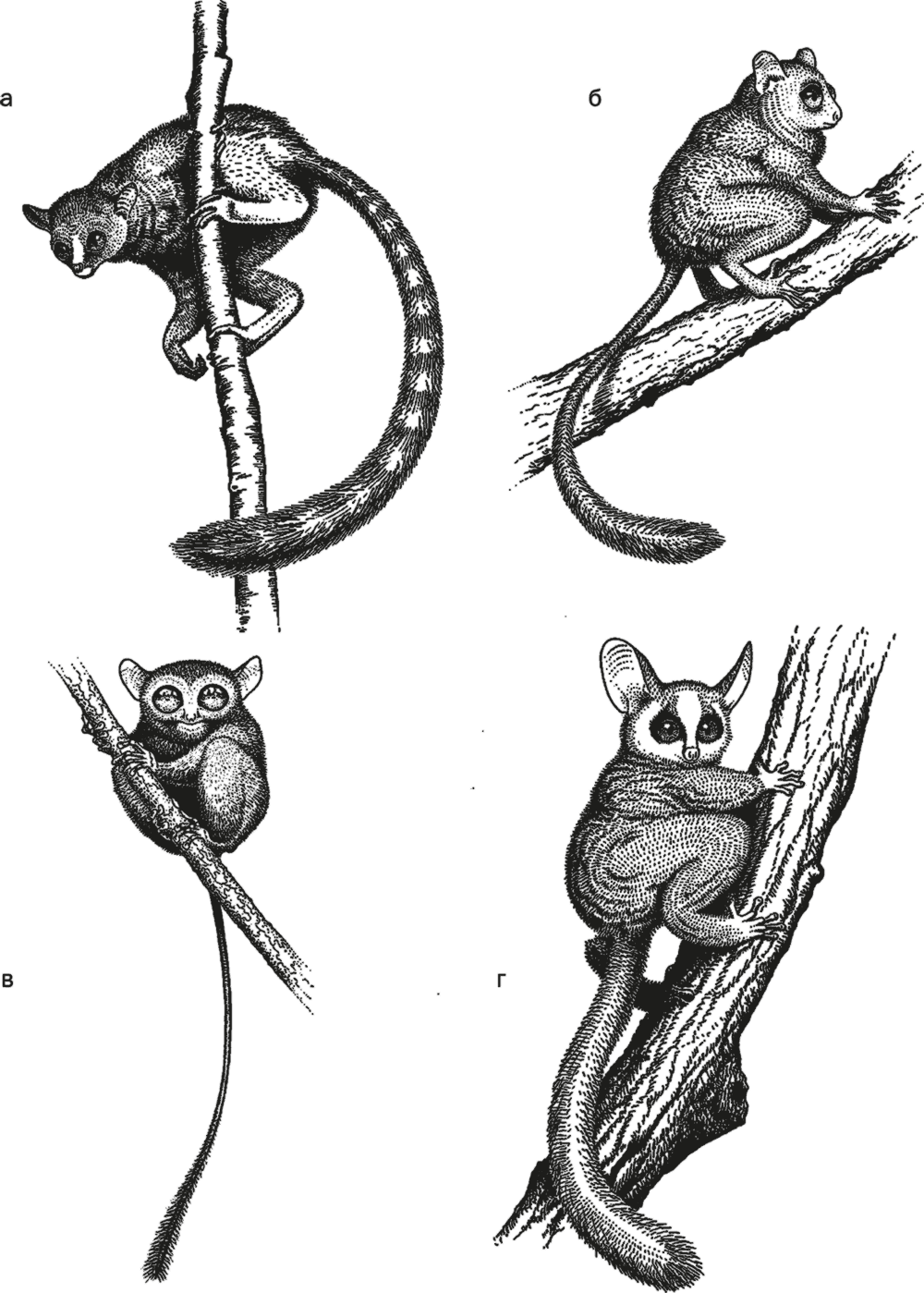
Рис. 30. Archicebus achilles (а), Necrolemur (б), долгопят (в) и галаго (г).
Омомисовые (Omomyiformes) объединяют великое множество эоценовых долгопятообразных. Они были, наряду с адаписовыми, основными участниками второй радиации приматов в раннем эоцене. Не исключено, что их заря зарделась еще в среднем палеоцене Азии, так как в эту эпоху в Китае жил декоредон Decoredon anhuiensis. Однако это существо известно крайне плохо, неспроста изначально декоредон был определен вообще как кондиляртр.
Несравненно лучше сохранился архицебус Archicebus achilles – самая древняя настоящая обезьяна, лучше всех прочих подходящая на роль общего предка долгопятообразных и антропоидов.
Великий Предок – древнейший, наицелейший, примитивнейший и просто красавец
А у Икарушки бедного,
Всеми братками забытого,
Только бледные ножки торчат
Из холодной зеленой водицы…
Д. В. Шагин
Находки целых скелетов древних животных – большая редкость. А если эти животные к тому же жили на деревьях и не любили воду (а скелеты сохраняются преимущественно в водных отложениях), то вероятность обнаружения их останков крайне мала. Потому древнейшие приматы известны плохо и почти исключительно по зубам. Потому сенсацией стала находка почти целого скелета одного из древнейших приматов – архицебуса Archicebus achilles.
В очередном номере журнала Nature китайско-американо-французская группа исследователей описала не только новый род и вид, но целое семейство древнейших приматов – архицебид Archicebidae (Ni et al., 2013). Новейшая находка уникальна по всем возможным показателям.
Во-первых, датировка: скелет сохранился в отложениях, образовавшихся в самом начале эоцена, 54,8–55,8 млн лет назад. Такого древнего и одновременно целого скелета обезьян еще никогда не находили. Были, правда, скелеты полуобезьян, но эти два главных подразделения отряда приматов – совсем не одно и то же. Как всегда, огромный интерес вызывает момент расхождения главных эволюционных ветвей, зарождение новой группы. Омомисовые известны со среднего палеоцена, но в подавляющем большинстве случаев от них сохранились лишь зубы размером с песчинку. Доселе примитивнейшим представителем группы была тейярдина азиатская Teilhardina asiatica, описанная в 2004 году из отложений границы палеоцена и эоцена Китая, а древнейшими – декоредон Decoredon anhuiensis из середины палеоцена (с его статусом есть большие проблемы, далеко не все палеонтологи признают его за примата), анеморисис Anemorhysis tenuiculus и тетониус Tetonius homunculus из конца палеоцена, еще две тейярдины Teilhardina brandti и T. magnoliana, баатромомис Baataromomys ulaanus и два вида вастаномисов Vastanomys gracilis и V. major, синхронные новоописанному архицебусу. Показательно, кстати, что всплеск разнообразия приходится на границу между эпохами палеоцена и эоцена, ведь эта граница потому и определима, что отражает существенные изменения климата. Однако все эти зверюшки известны по столь скудным останкам, что ученым приходилось напрягать все усилия, чтобы разобраться в их родстве и хоть как-то представить их облик.
Тут мы переходим ко второй уникальной особенности архицебуса – его сохранности. Скелет этого зверька сохранился не то чтобы целиком, но настолько, как палеонтологи могли только мечтать. Особенно радуют идеально сохранившиеся ножки и трогательный хвостик. С головой дела обстоят хуже, время не пощадило хрупкий череп, смяв его в не очень внятное пятно. Все же определимых и сравнимых признаков на скелете архицебуса на порядок больше, чем на любой другой находке древнейших обезьян (правда, сравнивать приходится опять же зубы, ибо от других приматов сохранились в основном они).
Благодаря этим сохранившимся признакам возможно оценить третью уникальность архицебуса – его крайнюю архаичность. По сочетанию признаков он оказывается максимально приближенным к ожидаемому первопредковому для всех обезьян состоянию: с одной стороны, он лишен явных черт полуобезьян, с другой – отсутствуют специализации. Например, у архицебуса нет сращения большой и малой берцовых костей, нет у него и крайнего удлинения пяточной кости. Эти особенности древних омомисовых и современных долгопятов всегда огорчали исследователей, ведь омомисовые в принципе подходят на роль предков прочих обезьян, но подобные специализации ставят на идее предковости крест. Доселе оставалось предполагать, что более поздние обезьяны произошли от ранних неспециализированных форм омомисовых, но внешний облик таких предков оставался в значительной мере гипотетическим. По одним зубам о строении конечностей много не скажешь. У архицебуса же ноги – какие надо ноги, из таких могли возникнуть как скакательные лапки долгопятообразных, так и хватательные – антропоидов.
Конечно, архицебус не мог не преподнести и сюрпризов. Например, в стопе предплюсна имеет умеренную длину, как у антропоидов, плюсна – большую, как у тупай и антропоидов (но в отличие от лемуров, адаписовых и долгопятов), а фаланги – тоже большую, как у долгопятов. Фаланги архицебуса сильно выпрямлены – специфическая особенность; самым длинным пальцем стопы был четвертый – это сближает архицебуса с адаписовыми и долгопятами.
На примере стопы отлично видна очередная уникальная особенность архицебуса – сочетание признаков разных групп обезьян. В нем есть основа от долгопятообразных, но немного от широконосых, немного от узконосых, а что-то – и от эосимид, которые сами сочетают черты узко– и широконосых. Много примитивного и мало специализированного – идеальное сочетание для идеального Великого Предка. Ранее известные приматы обычно без особых проблем могли быть классифицированы либо как долгопятообразные, либо как антропоиды. Если бы от архицебуса были найдены только зубы, его тоже без малейших сомнений причислили бы к примитивным долгопятообразным и его имя затерялось бы среди бесчисленных “-омисов”. Но теперь у нас слишком много информации, чтобы так упрощать картину. Становится очевидным, что ранние обезьяны уже имели, хоть бы в зародыше, массу особенностей, одни из которых развились и усилились в одних группах, а другие – в других. В очередной раз приятно видеть, что понятия примитивности и продвинутости крайне относительны.
Как же, наконец, выглядел архицебус и какой вел образ жизни?
Это было небольшое – с крупную мышь, 20–30 г – животное, с округлой головой, не слишком большими глазами, маленькой суженной мордочкой, крохотным носом и острыми зубами. Очень длинный хвост служил противовесом при прыжках. Скакать он мог благодаря очень длинным задним ногам, а сравнительно короткими передними лапками цеплялся за ветки и хватал насекомых. Впрочем, и способ передвижения, и тип питания не были специализированы. Архицебус прыгал совсем не так специфически, как современные галаго и долгопяты, а мог и бегать на четырех ногах по ветвям. Преимущественно насекомоядная диета также не исключает того, что архицебус мог есть и другую пищу. Судя по размерам глаз, он вел дневной образ жизни.
Самый целый из самых древних, наиболее примитивный и одновременно генерализованный неспециализированный, воплотивший в себе идеал Великого Общего Предка – о чем еще можно мечтать?! Приятно, когда в одной находке сочетается столько достоинств! Порадуемся же преодолению еще одной ступени познания!
На самом деле, не только архицебус может быть предком высших обезьян, просто он очень уж хорошо сохранился. Из Индии известен вастаномис Vastanomys (54–55 млн лет назад), который обладал едва ли не самым примитивным среди всех омомисовых строением и, судя по зубам, был не насекомо-, а фруктоядным. Но от вастаномисов сохранились одни зубы, так что мы не знаем о строении их ног или черепа.
Таким образом, омомисовые явились предками долгопятов и высших обезьян – такая версия называется тарзиоидной гипотезой. Слава эволюции, что в самом начале группы омомисовых нашелся не слишком специализированный примат, давший начало более продвинутым обезьянам. Судя по всему, омомисовые специализировались чрезвычайно быстро, не успей зародиться линия антропоидов в начале эоцена, они могли бы и вовсе не появиться. Или же эволюция воспользовалась бы материалом из других групп – адаписовых или, возможно, даже плезиадапиформов.
Основные группы омомисовых – омомиды Omomyidae и микрохэриды Microchoeridae – жили в Северной Америке, Европе, Северной Африке, Южной и Восточной Азии. Те и другие имели очень маленькую – укороченную и суженную спереди – мордочку и округлый череп с огромными направленными вперед глазами. В строении зубов многих видов сохраняется такой примитивный признак, как четыре премоляра. Ноги омомисовых были длинными, со сросшимися малой и большой берцовой костями и резко увеличенной пяткой, образующей цевку. Очевидно, это были ночные, насекомоядные и активно прыгающие животные. Правда, во всех этих чертах омомисовые не превзошли долгопятов, но стремились к идеалу как могли; в силу не слишком далеко зашедшей специализации омомисовые могли внешне напоминать скорее галаго, чем долгопятов.
Микрохэриды были очень разнообразными в размерах животными: от 45 г у Pseudoloris parvulus до 1775 г у Microchoerus erinaceus. Объединяются они благодаря специфическому строению зубов: в частности, первые верхние и нижние резцы несколько увеличены и выдвинуты перед вторыми резцами. Не очень ясно, зачем им нужна была эта специализация.
Омомиды чрезвычайно многочисленны. Большинство их весило от 100 до 200 г, но были и рекордсмены: Teilhardina magnoliana была размером с мышь – 28 г, а Anaptomorphus westi, Gazinius amplus и Stockia powayensis с крысу – 465–475 г, Hemiacodon gracilis тянул на целый килограмм, а представители родов Rooneyia, Ourayia и Macrotarsius – от килограмма до 2520 г! Часть омомид питались преимущественно насекомыми (например, многие в подсемействе Omomyinae и представители трибы Washakiini), однако, в отличие от современных долгопятов, очень многие были фруктоядными (например, почти все представители подсемейства Anaptomorphinae), а некоторые даже листоядными (например, Hemiacodon gracilis).
Кстати, о размерах…
Размеры ископаемых приматов определяются разными способами. Конечно, идеальный случай, когда в нашем распоряжении есть целый скелет. Но это бывает крайне редко. Чаще приходится довольствоваться отдельными зубами. Тогда в дело идет регрессионный анализ: из соотношений размеров зубов (например, площади первого верхнего или длины второго нижнего моляра) и веса тела у современных приматов высчитывается вес ископаемых животных. Дело Ж. Кювье живет и побеждает! Понятно, что погрешность таких вычислений весьма велика – у разных авторов и полученные разными методами цифры могут отличаться в разы, – да и индивидуальную изменчивость никто не отменял, так что публикуемые величины не должны восприниматься слишком буквально, а служат лишь примерным ориентиром.
Можно обратить внимание, что почти никогда речь не идет о линейных размерах – длине тела. Дело в том, что приматы – гибкие зверюшки, они удивительно пластичны, даже у человека рост при нескольких измерениях легко может различаться на несколько сантиметров. Так что палеоприматологи предпочитают пользоваться определениями веса.
Altanius orlovi – единственная полуобезьяна из Монголии; он определяет северную известную границу распространения этих животных в Азии. Меж тем фауны Северной Америки и Евразии весьма схожи. Либо обезьяны все же могли преодолевать Берингию, просто мы пока не нашли их в Сибири, либо они периодически форсировали расширявшиеся моря между Северной Америкой и Европой и между Европой и Азией, ибо Европа была островом между двух континентов. По этим же географическим причинам североамериканские омомисовые (как, кстати, и адаписовые) намного разнообразнее европейских: Северная Америка больше по размеру и богаче на условия, вот и животные были отличающимися, тогда как в Европе имелось меньше вариаций экологических ниш, отчего и приматы были схожими (Gilbert, 2005).
Омомисовые в Европе и Северной Америке дожили до границы эоцена и олигоцена (около 34 млн лет назад или чуть позже), после чего вымерли по тем же причинам, что свели в могилу адаписовых и многих прочих животных, процветавших в тропических лесах эоцена.
Долгопятовые (Tarsiiformes) современности обычно объединяются в один род Tarsius, хотя предложено разделить его на три, сохранив прежнее название лишь за сулавесскими видами, тогда как суматранско-калимантанские выделяются в род Cephalopachus, а филиппинские – Carlito (Groves et Shekelle, 2010). Впрочем, разница между ними очень невелика, даром что разошлись островные популяции как минимум еще в миоцене.
В строении долгопятов совмещаются признаки полуобезьян и настоящих обезьян. Это животные размером с крысу, с очень большими глазами и специализированными для прыгания ногами. Они активны ночью и питаются насекомыми. Во многих чертах строения проявляется резкая специализация: глаза столь велики, что не могут вращаться, отчего подвижность шеи резко увеличена, благодаря огромному позвоночному отверстию атланта (первого шейного позвонка) и хитрому строению межпозвоночных сочленений шея может поворачиваться задом наперед; мордочка крайне маленькая; кончики пальцев расширены в виде присосок, большая и малая берцовые кости срастаются, пятка резко удлинена в виде цевки для прыгания по ветвям, а второй и третий пальцы стопы снабжены когтями. Живут долгопяты семейными группами, общаясь друг с другом своеобразным щебетом, похожим на птичий.
О долгопятах подробно говорилось выше, тут помянем лишь их происхождение.
45 млн лет назад в Китае жили как минимум три вида долгопятов или чрезвычайно похожих на них существ, правда, известны они только по таранной и пяточным костям. Один из них весил всего 30 г – вдвое меньше самых маленьких долгопятов: карликовых Tarsius pumilus. Из того же времени Китая описан Xanthorhysis tabrumi, но от него найдены лишь зубы, так что совпадает ли он с одним из трех “таранно-пяточных” видов – непонятно.
Некий долгопятовый зверек жил в Таиланде около 34 млн лет назад, но известен он лишь по одному зубу. В это же время в Китае скакал Oligotarsius rarus.
Миоценовый Hesperotarsius sindhensis обитал 16–18 млн лет назад в Пакистане. Своим существованием он указывает, что древние долгопяты жили далеко за пределами своего нынешнего ареала.
К современному роду относятся три ископаемых вида, причем древнейший – Tarsius eocaenus – жил аж в эоцене, более 37 млн лет назад, в Китае. Почти современно выглядят Tarsius thailandica (18 млн лет назад) и Tarsius sirindhornae (13,1–13,3 млн лет назад) из Таиланда.
Выжить в Индонезии и на Филиппинах долгопятам помогла крайняя специализация, а позволил не слишком сильно менявшийся климат.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК