Новые приматы Старого Света: примитивнейшие узконосые обезьяны
Узконосые обезьяны (Catarrhini) – обезьяны Старого Света: Африки, Азии и Европы. Они появились уже в верхнем эоцене, а на границе с олигоценом стали достаточно разнообразными. Как всегда бывает, переходные группы трудно однозначно классифицировать, так как они зависают между предками и потомками. В данном случае палеоприматологи нашли элегантный выход: примитивнейшие узконосые, которые не могут быть однозначно определены как мартышковые или человекообразные, выделяются в парвотряд “ранних узконосых” Eocatarrhini, тогда как более продвинутые формы, имеющие непосредственное отношение к современным, обозначаются как “истинные узконосые” Eucatarrhini. Маленькая буковка в названии, но огромный скачок в морфологии!
“Ранние узконосые” (Eocatarrhini) сами по себе подразделяются на две группы: более древние и примитивные проплиопитековые (Propliopithecoidea) известны исключительно с границы эоцена и олигоцена Северной Африки (почти исключительно из Египта), а поздние и относительно продвинутые плиопитековые (Pliopithecoidea) – из миоцена Уганды, Европы и Азии.
Проплиопитековые (Propliopithecoidea) имели еще довольно много архаичных черт строения, поэтому их систематический статус неустойчив: разные палеоприматологи предлагали объединять их с парапитековыми или, напротив, включать в состав человекообразных. Самые ранние формы – самые загадочные, что, в общем-то, закономерно.
Олигопитековые (Oligopithecinae), названные по роду Oligopithecus из нижнего олигоцена (31–33 млн лет назад) оазиса Файюм в Египте (предположительно туда же включаются катопитек Catopithecus browni из более старых отложений того же Файюма и еще более древний талапитек Talahpithecus parvus из верхнего эоцена Ливии), разными исследователями определялись как самостоятельное семейство Oligopithecidae надсемейства проплиопитековых (Propliopithecoidea), как подсемейство семейств проплиопитецид (Propliopithecidae) или плиопитецид (Pliopithecidae; в варианте с выделением подсемейств Propliopithecinae, Pliopithecinae и Oligopithecinae), как синоним проплиопитецид или парапитецин (Parapithecinae), включались в понгид (Pongidae) и даже в надсемейство адаписовых (Adapoidea), а также в качестве семейства в инфраотряд Paracatarrhini гипотряда Anthropoidea. Можно надеяться, что новые находки позволят точнее определиться в этом сложном вопросе. В то же время и в том же месте, что олигопитеки, жил странный примат – протеопитек Proteopithecus sylviae, выделенный в собственное семейство Proteopithecidae; впрочем, по общему эволюционному уровню, размерам и образу жизни он не очень отличался от своих соседей.
Очень здорово, что приматы Файюма были массовыми животными, особенно многочисленны были катопитеки, на втором месте по частоте – протеопитеки. От них, да и других приматов сохранились не только зубы, но и черепа и части скелетов. Благодаря такому богатству нам известны многочисленные подробности строения и образа жизни египетских позднеэоценовых и раннеолигоценовых обезьян; ни о каких других синхронных приматах у нас нет столь обширных сведений.
Олигопитековые и протеопитек еще имели некоторые примитивные черты: например, барабанная пластинка у них не полностью окружала слуховой канал. Зато был и прогресс: скажем, в челюстях осталась всего пара премоляров – черта, не очень надежно, но наглядно разграничивающая узконосых и всех прочих приматов. Олигопитековые имели вес от трети до полутора килограммов, вели дневной образ жизни, питались преимущественно фруктами, дополняя их листьями и иногда насекомыми; протеопитек мог вдобавок к тому лакомиться смолами и нектаром.
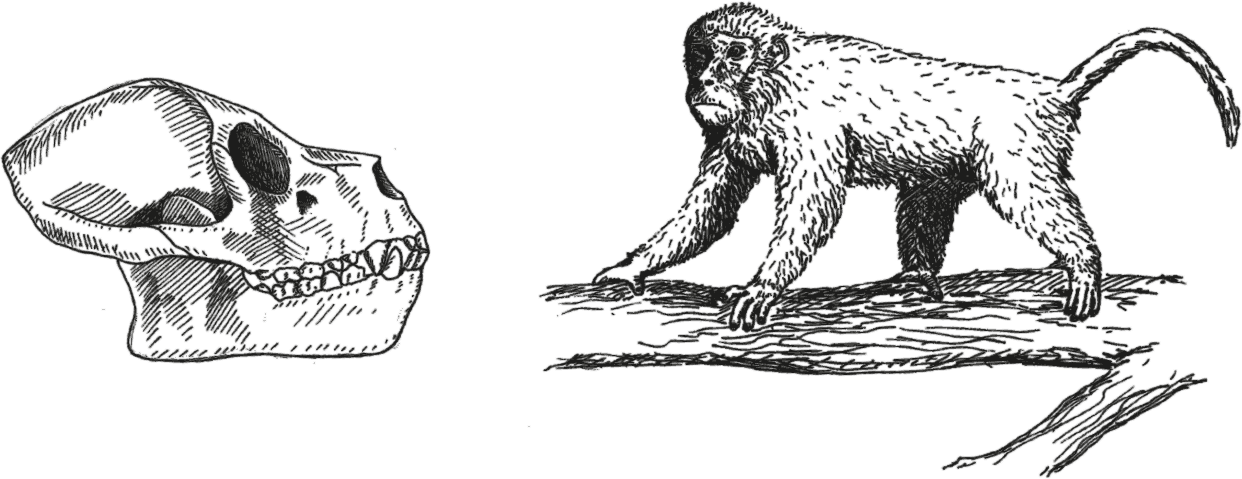
Рис. 33. Череп Aegyptopithecus zeuxis и реконструкция Propliopithecus.
Любопытно, что половой диморфизм по размерам клыков у катопитека и особенно протеопитека был выражен сильнее, чем у схожих по размеру тела современных обезьян (Simons et al., 1999). Крупные размеры клыков у самцов и мелкие у самок свидетельствуют о существенной межсамцовой конкуренции. Группы катопитеков и протеопитеков состояли либо из нескольких самцов со строгой иерархией и множества самок, либо вообще из одного самца с гаремом самок. Такая структура нетипична для полуобезьян, зато характерна для многих узконосых.
Пяточная кость протеопитека замечательна чудной смесью признаков, одни из которых почти идентичны чертам омомисовых, другие – видимо, конвергентно – мартышковых, а кои – так и вообще адаписовых (Gladman et al., 2013). Очевидно, на этом эволюционном уровне строение конечностей отставало от челюстей и зубов.
Позднейшим олигопитековым был некий неопределенный примат, живший в начале олигоцена в Египте. Он отличался карликовыми размерами – существенно меньшими, чем у Catopithecus и Oligopithecus (Seiffert et Simons, 2013). Трудно сказать, что довело его до жизни такой, а в конце концов и до вымирания.
Кстати, о Файюме…
Файюмский оазис широко известен среди искусствоведов файюмскими портретами, сделанными на крышках саркофагов (кстати, это отличный источник антропологической информации). Но палеонтологи любят Файюм совсем не за это: в нескольких карьерах они нашли и продолжают раскапывать сотни или даже тысячи окаменелостей эоценовых и олигоценовых животных. В Файюме найдены одни из древнейших слонов и слоновых прыгунчиков, даманов и сирен, антракотериев и китов, странных арсиноитериев и еще более странных птолемайид, летучих мышей и грызунов. Нынешняя пустыня 41–29 млн лет назад была цветущим садом, в болотах которого жизнь била ключом. Из Файюма известны самые разные приматы, в том числе предковые для нескольких крупных современных групп.
Проплиопитековые в узком смысле (Propliopithecinae) не слишком разнообразны – Aegyptopithecus zeuxis и четыре вида рода Propliopithecus – и известны только из Файюма в узком хронологическом интервале 29,5–31,5 млн лет назад, но чрезвычайно важны филогенетически. Один клык найден в Анголе.
Сложность разграничения парапитековых и “ранних узконосых” отлично видна на примере оманской находки в местонахождении Таках. Тут был обнаружен обломок нижней челюсти с пятью зубами. Первоначально он был описан как мэрипитек Moeripithecus markgrafi – вид, ранее известный из египетского Файюма (Thomas et al., 1991). Мэрипитек похож на проплиопитека, почему включался в проплиопитековых и даже иногда обозначался как Propliopithecus markgrafi. Зубы из Файюма, описанные как Propliopithecus ankeli, некоторыми палеоприматологами тоже определялись как принадлежавшие Moeripithecus markgrafi. Однако другие исследователи склонны обращать большее внимание на примитивные черты мэрипитека и включать его в парапитековых. В то же время оманская челюсть больше сближается с видом Propliopithecus ankeli, который признается самостоятельным и остается в проплиопитековых. И такая путаница сопровождает исследование почти каждой находки примитивных обезьян с границы эоцена и олигоцена.
Другой пример – алжирипитек Algeripithecus minutus: он описывался как парапитековая или плиопитековая обезьяна, однако с большей вероятностью является адаписовой полуобезьяной (Tabuce et al., 2009).
Из проплиопитековых лучше всего изучен египтопитек Aegyptopithecus zeuxis из олигоцена Египта (29,5–30,2 млн лет назад), от которого сохранились несколько целых черепов и фрагментарные скелеты. На первый взгляд египтопитек напоминал современных макак, благо он практически полностью соответствует им и размерами (2,5–6 кг), и пропорциями. Однако приглядевшись, в строении египтопитека нетрудно заметить массу примитивных черт: задняя стенка глазницы закрыта, да не вполне, барабанная пластинка окружает слуховой канал, но тоже не полностью, – тот и другой признаки развиты примерно как у широконосых обезьян. Совсем подкачал мозг: он у египтопитека вдвое меньше, нежели даже у карликовой мартышки (Radinsky, 1973; Simons et al., 2007). И относительный размер, и пропорции мозга египтопитека вообще оказываются как у мадагаскарских лемуров, хотя участки мозга, отвечающие за зрение, у египтопитека чуть больше, а за обоняние – чуть меньше; впрочем, на фоне современных обезьян эти “чуть” не считаются. Между прочим, это говорит о том, что увеличение мозга у широконосых и узконосых обезьян шло независимыми путями, коли уж общие предки по этому показателю находились на уровне развития полуобезьян. А это, в свою очередь, позволяет надеяться на появление в будущем новых разумных видов.
Судя по маленьким глазницам, египтопитек был дневным, а их разворот вперед говорит о хороших способностях к лазанию по деревьям. О том же свидетельствует строение конечностей: эти животные бегали по ветвям примерно как современные ревуны. В строении конечностей вообще много сходства именно с широконосыми обезьянами: руки и ноги были одинаковой длины, относительно толстые и короткие, неспособные полностью выпрямляться, зато очень цепкие. Далеко прыгать египтопитеки вряд ли умели, но хвост не утеряли, хотя вряд ли он был очень длинным.
Строение зубов свидетельствует, что египтопитеки и проплиопитеки питались почти исключительно фруктами, восполняя недостаток белков листьями. Таким образом, насекомоядное прошлое наконец-то было окончательно изжито.
Обилие черепов позволило оценить половой диморфизм и даже возрастные изменения египтопитеков: самки были существенно – вдвое – меньше и имели меньшие клыки и сагиттальные гребни на черепе. У проплиопитеков разница полов была далеко не такая значительная. С возрастом, как положено, росла морда и увеличивался сагиттальный гребень (Simons, 1987).
Плиопитековые (Pliopithecoidea), вероятно, были потомками проплиопитековых. Эта странная группа процветала от начала до конца миоцена (20–7 млн лет назад) в Китае, Таиланде, Пакистане и Европе, а одна спорная находка сделана в Уганде. Систематика плиопитековых остается предметом бурных дискуссий. С одной стороны, они объединялись с проплиопитековыми, с другой – с гиббонами. В одном из новейших вариантов плиопитековые делятся на дионисопитецид (Dionysopithecidae) и плиопитецид (Pliopithecidae); последние же – на более древнее подсемейство плиопитецин (Pliopithecinae) и более молодое крузелин (Crouzeliinae).
Кстати, о миоцене…
Граница олигоцена и миоцена ознаменовалась существенным потеплением и таянием льдов Антарктиды. Тепло, правда, не вернулось к эоценовому уровню, но все же существенно превосходило современные показатели. Карта мира в целом и Евразии в частности приобрела более-менее современный вид, только юг Европы еще существенно отличался: как бы Средиземное море тянулось аж до середины нынешних Украины и Казахстана, а посреди этих вод возвышались острова, которых не найти на современной карте, в числе коих знаменит Гаргано.
Примерно с середины миоцена опять стало холодать, причем ударными темпами, так что с климатической точки зрения границу между эпохами логично было бы проводить именно тут – порядка 12–14 млн лет назад.
Дионисопитециды (Dionysopithecidae) представлены всего тремя нижне-среднемиоценовыми (12–20 млн лет назад) видами из Таиланда, Китая и, видимо, Пакистана – Dionysopithecus orientalis, D. shuangouensis и Platodontopithecus jianghuaiensis, но известны фрагментарные и плохо изученные находки, которые могут представлять иные роды и виды. Сложность, как обычно, в том, что найдены почти исключительно изолированные зубы. Отдельными чертами эти животные уже настолько напоминали человекообразных обезьян, что некоторые палеонтологи объединяют их с проконсулами, хотя в целом они примитивнее плиопитецид. Дионисопитек числится в сонме вероятных предков гиббонов, благо и размером он был похож – около 15 кг. Вместе с тем пяточная кость платодонтопитека довольно примитивна и похожа на кости плиопитеков и проконсулид, а если выбирать между мартышковыми и человекообразными, то на мартышковых. Фаланги платодонтопитека очень вытянуты, по этому показателю их превосходят только гиббоны (Harrison et Yumin, 1999). Дионисопитециды были вероятнейшими предками плиопитецин, которые, таким образом, имели прародину в Азии.
Не исключено, правда, что плиопитековые возникли не в Азии, а в Африке. За это говорит существование ломорупитека Lomorupithecus harrisoni в Уганде 15–20 млн лет назад и возможного плиопитекового в Кении в местонахождении Форт Тернан, правда, уже в конце миоцена 12,5–14 млн лет назад (его зуб был отнесен к Limnopithecus – роду, раньше причислявшемуся к плиопитековым, но ныне считающемуся проконсулидом, то есть человекообразным). Ломорупитек известен по разбитому черепу, в целом похожему на плиопитековых Европы. Его древность – едва ли не старше дионисопитецид – и географическое расположение говорят, что корни плиопитековых могут быть глубже, чем нам известно.
Плиопитецины (Pliopithecinae) сыграли большую роль в истории палеонтологии, поскольку их останки впервые были найдены и описаны еще в середине XIX века. Благодаря этому они вошли во все учебники антропологии, хотя, строго говоря, наверняка были тупиковой группой приматов, имеющей к человеку лишь косвенное отношение. Они известны из Европы и Китая, где существовали не слишком долго – от 17 до 11,6 млн лет назад. Достоверно известен лишь один род Pliopithecus, в котором выделяются шесть-семь похожих видов. Некоторые роды от схемы к схеме кочуют между плиопитецинами и крузелинами.
Плиопитеки из всех ископаемых приматов в наибольшей степени похожи на гиббонов, причем как строением черепа, так и зубов и конечностей. Неспроста самый примитивный европейский вид Pliopithecus piveteaui был даже описан под названием Hylobates antiquus. Впрочем, это не значит, что плиопитеки больше всего похожи на гиббонов из всех современных приматов; при таком ракурсе сравнения плиопитеки сближаются скорее с широконосыми или мартышкообразными. Плиопитеки были довольно резвыми древолазами, но точно не брахиаторами.
Особенно здорово изучен Pliopithecus vindobonensis (15–15,5 млн лет назад), так как в Словакии были найдены многочисленные его останки, а Г. Запфе описал их в монументальном труде (Zapfe, 1960).
Уголок занудства
Плиопитек имел укороченную и суженную морду, сравнительно грацильную мозговую коробку округлой формы с более-менее вертикальным лбом, умеренно развитые височные линии, либо не сливавшиеся, либо образующие низкий сагиттальный гребень. Глазницы довольно крупные, ориентированные почти вперед, но все же несколько вбок, их край утолщен почти в виде кольца. Наглядное отличие от мартышкообразных – широкое межглазничное пространство. Скуловая область очень низкая. Угол нижней челюсти выступает назад. Особенности строения зубов выдают фруктоядность. Примитивной чертой является неполное срастание барабанной пластинки – для миоцена очень примитивная черта. В сравнении с гиббоном плиопитек имел практически той же длины ноги, но куда как более короткие и толстые руки. Если у гиббона руки заметно длиннее ног, то у плиопитека – наоборот; пропорциями он больше напоминал ревунов или павианов. Кисти и стопы плиопитека были удлинены, а фаланги сужены и изогнуты, хотя и не в такой степени, как у гиббона. Тело, судя по всему, было тонкое и длинное. Строение крестца наводит на мысли о сохранении хвоста.
Крузелины (Crouzeliinae): Plesiopliopithecus, Crouzelia, Barberapithecus, Anapithecus, Egarapithecus и Laccopithecus – более поздняя и продвинутая группа плиопитековых. Они известны почти исключительно из Европы (14,5–9 млн лет назад), только Laccopithecus robustus жил в Южном Китае (6–8 млн лет назад). Крузелия Crouzelia и лаккопитек Laccopithecus – очередные предполагавшиеся предки гиббонов.

Рис. 34. Череп Pliopithecus vindobonensis.
Больше останков сохранилось от анапитека Anapithecus hernyaki, особенно в венгерском местонахождении Рудабанья (9,5 млн лет назад). Это обезьяна весила 15 кг. Замечательно, что при малых различиях в размерах тела самцы и самки существенно отличались размерами клыков. Это говорит о своеобразной социальной структуре анапитеков. В черепе анапитеков вполне объемно повторяется план строения черепа более древнего плиопитека: крупные размеры, округлая мозговая коробка, сравнительно вертикальный лоб, чуть развернутые вбок глазницы с утолщенным краем, широкое межглазничное пространство, короткая морда с низкой скуловой областью. Судя по зубам, анапитеки, как и прочие плиопитековые, были фруктоядными (DeMiguel et al., 2013), хотя раньше предполагалась их листоядность. Есть и специфика: верхнечелюстные пазухи анапитеков уменьшены, хотя и не исчезли, как у мартышкообразных. Кости остального скелета тоже похожи на плиопитечий вариант, хотя фаланги изогнуты сильнее.
Кстати, о бедрах…
Сложность изучения фрагментарных останков нагляднейше видна в истории с бедром из Эппельсгейма (K?hler et al., 2002). В Германии еще в 1820 г. была найдена бедренная кость довольно большого размера. Первоначально она была описана как принадлежащая дриопитеку, хотя получила сразу три названия – Paidopithex rhenanus, Pliohylobates eppelsheimensis и Pliosymphalangus. После было показано, что она заметно отличается от варианта, типичного для человекообразных обезьян, зато весьма напоминает бедренную кость Pliopithecus vindobonensis. Однако европейские плиопитеки весили около 10 кг, а эппельсгеймский примат – 22. По размерам к нему приближается Pliopithecus zhanxiangi, но он найден в Китае. В 1935 г. в том же местонахождении был найден клык самца примата, описанный как Semnopithecus eppelsheimensis, который впоследствии определялся как Pliopithecus, Rhenopithecus или Dryopithecus. В итоге плиопитековая его природа была признана наиболее вероятной, но размеры зуба говорят о том, что его обладатель был вдвое меньше, чем владелец бедра, так что отнесение их к одному виду опять же совершенно необязательно. Далее, в рядом расположенном местонахождении Сальмендинген нашлись зубы, определенные как Anapithecus hernyaki – то есть опять же, плиопитековый примат. А анапитек весил 15 кг – уже ближе к размерам эппельсгеймской обезьяны. Однако в 1990-х гг. в плиопитековой природе этих зубов появились основательные сомнения, и исследователи стали склоняться к дриопитековой версии. Наконец, очередное подробное морфологическое исследование показало, что бедро наверняка не принадлежало дриопитеку, им мог обладать либо неизвестный доселе очень крупный плиопитековый примат, либо вообще совсем своеобразная узконосая обезьяна из “третьей группы” (первые две – плиопитековые и человекообразные).
Плиопитековые являют собой своеобразный аналог широконосых обезьян в Старом Свете, евразийский эксперимент: с наибольшей вероятностью они возникли в Азии, но дали всплеск разнообразия в Европе, где до этого крупных растительноядных приматов не было. Олигоценовые холода выкосили европейских жителей лесов, к середине этой эпохи тут исчезли и адаписовые, и омомисовые. Как и в случае с Южной Америкой, свободные экологические ниши просто-таки томились – когда же их займут новые жильцы, и те не заставили себя ждать. Как только в миоцене новые теплые леса заколосились от Испании до Таиланда, в рекордные сроки кустообразная эволюция заполнила европейские пущи крикливыми пожирателями плодов и листьев. Именно по этой причине быстрого видообразования из малого числа предков так трудно классифицировать плиопитековых – тут параллель с широконосыми тоже совершенно очевидна.
Правда, в это же время из Африки и с Ближнего Востока подтянулись человекообразные и мартышкообразные, составившие плиопитековым мощную конкуренцию. В конце же миоцена и вовсе стало грустно: леса сохли и заменялись саваннами. В Африке это привело к появлению австралопитеков, а в Европе – к глобальному вымиранию лесных приматов. Как-то не вставалось им на две ноги.
Последние крузелины – они же последние плиопитековые и самые специализированные среди прочих – исчезли 9 млн лет назад в Европе (Egarapithecus narcisoi) и 6–8 – в Китае (Laccopithecus robustus).
Минутка фантазии
Давным-давно никто не считает плиопитековых предками человека. Но это не отменяет того, что все задатки для появления разумности у этих обезьян были. Они обладали относительно большим мозгом и маленькими челюстями, не слишком сильным половым диморфизмом, не имели резких специализаций в строении конечностей. В каком-то смысле они находились даже в выигрышных условиях по сравнению с предками австралопитеков Африки, ведь условия Европы были более суровыми, а стало быть – располагающими к прогрессивной эволюции. В северных широтах раньше стали исчезать тропические леса, появляться саванны и степи. Тут бы плиопитекам и слезть с деревьев, заселить просторы и устроить марш на восток, юг и куда еще захочется… Что остановило их эволюцию – не вполне очевидно. С одной стороны, вымирание плиопитековых совпадает с концом миоцена и резким похолоданием, с другой – южнокитайские виды, жившие далеко на юге, потенциально могли развиваться дальше. Надо думать, основную роль сыграла конкуренция с мартышкообразными и человекообразными: “истинные узконосые” оказались быстрее, бодрее и нахальнее. Показательно, что у анапитеков постоянные зубы прорезывались заметно раньше, чем у мартышкообразных сопоставимого размера (Nargolwalla et al., 2005). Предполагалось, что раннее взросление было связано с усиленным прессом со стороны хищников или листоядностью, но новейшие исследования говорят скорее о фруктоядности анапитеков. Возможно, ускоренное развитие было способом конкуренции с “истинными узконосыми”. Как бы то ни было, укороченное детство означает меньшие возможности для обучения и в итоге интеллектуальное отставание от конкурентов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК