Прямохождение
Широко известно полуанекдотическое определение человека как “двуногого без перьев”. Но человеку мало двуногости – он еще и ходит выпрямившись, в отличие от, скажем, птиц или тушканчиков, позвоночник которых расположен горизонтально. Прямохождение обеспечивается целым рядом специальных признаков.
Первый – положение большого затылочного отверстия на затылочной кости, через которое спинной мозг соединяется с головным. У четвероногих животных оно находится в задней части основания черепа и повернуто назад, так как позвоночник прикрепляется если и не строго сзади, то по меньшей мере наискосок; у прямоходящих существ отверстие расположено в центре длины основания черепа и открывается вниз, так как позвоночник подходит снизу, а передняя и задняя части черепа должны быть уравновешены. Соответственно, существенно меняются и места прикрепления шейных и спинных мышц, конфигурация затылка, укорачивается основание черепа.
Вариант промежуточного типа известен уже у Sahelanthropus tchadensis около 7 млн лет назад, заметно более продвинутый – у Ardipithecus ramidus 4,4 млн лет назад, а окончательно “прямоходящий” – у Australopithecus afarensis 3,5 млн лет назад.
Позвоночник четвероногих получает примерно равную нагрузку спереди и сзади, отчего размеры позвонков вдоль длины более-менее одинаковы, а центральная часть представляет собой равномерную дугу – аналог арочного моста. Крестец – часть позвоночника, к которой крепится таз, – имеет сильно вытянутую узкую форму, а его позвонки могут либо вообще оставаться независимыми, либо сливаться не полностью. У прямоходящих позвоночник ориентирован вертикально, а потому имеет характерные изгибы – лордозы вперед и кифозы назад; в итоге вся конструкция становится более-менее устойчивой и работает как пружина, а нагрузка стучит не строго по нижележащим позвонкам, а частично распределяется в стороны. Размеры позвонков у прямоходящих закономерно увеличиваются сверху вниз, самыми крупными оказываются нижние поясничные и верхние крестцовые, так как тут расположен центр тяжести организма. Крестец широкий и короткий, его элементы капитально сливаются между собой. У австралопитеков Australopithecus afarensis и Australopithecus africanus изгибы, вероятно, были как у современного человека, но некоторые детали строения позвонков (например, вытянутость тела позвонков спереди назад) сближают их с обезьянами.

Рис. 1. Основание черепа и большое затылочное отверстие шимпанзе (а), австралопитека (б) и человека (в).
Строение крестца у австралопитеков – начиная с Ardipithecus ramidus и Australopithecus afarensis – типично гоминидное. Конечно, не стоит абсолютизировать помянутые признаки. Например, у гориллы есть поясничный лордоз, но его появление связано не с прямохождением, а с огромным весом животного.
Строение таза очевидно разное при разных типах передвижения: у четвероногих обезьян таз узкий, высокий и вытянутый вдоль позвоночника, а у прямоходящих – широкий и низкий. Конечно, стоит помнить, что форма таза обусловлена еще и нуждами деторождения, но, судя по тому, что у австралопитеков голова была фактически обезьянья (а проблемы при родах создает как раз голова – она большая и плохо сжимается), а таз – человеческий, фактор локомоции играл определяющую роль.
Уголок занудства
На человеческой тазовой кости отлично выражена передняя нижняя подвздошная ость (костный выступ на переднем крае таза, читатель без труда может найти у себя верхнюю подвздошную ость – на ней держится ремень, нижняя скрыта мышцами; у обезьян нижней подвздошной ости фактически нет), а ушковидная поверхность и подвздошная бугристость, обеспечивающие соединение с крестцом, занимают значительную площадь. Седалищный бугор у человека могуч, а седалищная ость длинна и остра, тогда как у других обезьян первый короток, а второй вообще почти нет.
На передней нижней подвздошной ости крепится прямая мышца бедра (часть четырехглавой мышцы бедра), разгибающая колено. Но она же сгибает бедро в тазобедренном суставе, то есть поднимает ногу вперед (или опускает тело, если нога прочно стоит на земле). Получается противоречие: одна и та же мышца одновременно держит тело вертикально в одном суставе, но наклоняет его в другом. У четвероногих животных такой странной проблемы нет, ведь у них нога вертикальна, а тело горизонтально, мышца совершенно логично выполняет шаг вперед. Чтобы нога у человека была выпрямлена и в тазобедренном суставе тоже, четырехглавой мышце бедра противостоят целых три ягодичные мышцы – большая, средняя и малая, которые разгибают бедро, удерживая тело в вертикальном положении. Особенно преуспевает в этом большая ягодичная мышца. Но у обезьян она расположена не сзади, а сбоку и занята иным делом – отводит бедро в сторону, а потому развита умеренно, почему и зад у обезьян тощий, совсем нечеловеческий, а выпрямить ноги они не могут. Кстати, совершенно обезьяний вариант типичен для новорожденных младенцев. Поэтому они лежат в позе “куренка табака” – растопырив в стороны согнутые ножки, а свести вместе их практически не могут. Только потом четырехглавая мышца “переползает” на человеческое место – в “пятую точку”; если же по какой-то причине замедленный рост сухожилий и фасций препятствует этому перемещению, могут возникнуть проблемы с умением ходить.
Раз у человека пропала отводящая функция большой ягодичной мышцы, должна быть какая-то компенсация: отведением бедра в сторону у него усиленно занимаются верхняя и нижняя близнецовые мышцы, отчего место их крепления на седалищной кости чрезмерно увеличивается и становится седалищной остью. Но еще важнее человеку приводить бедро, то есть сводить ноги вместе, чтобы стоять прямо; этим, среди прочих, заняты квадратная мышца бедра и большая приводящая мышца, крепящиеся к седалищному бугру, который оказывается заметно крупнее, чем у обезьян.
Тазовые кости сохраняются плохо, но радует, что у Ardipithecus ramidus 4,4 млн лет назад мы видим строго промежуточный “четвероного-двуногий” вариант – одновременно и широкий, и вытянутый в длину. У Australopithecus afarensis 3,5 млн лет назад таз был не только полностью “прямоходящий”, но по расширенности пропорций даже более человеческий, чем у современного человека. Это не значит, что мы сколько-нибудь вернулись к четвероногости. Ясно, что сей парадокс связан с так называемыми аллометрическими закономерностями. При изменении общих размеров тела разные части тела меняются неодинаково: при росте от малых размеров к средним длиннотные размеры увеличиваются быстрее, чем широтные (а вот если увеличивать человека до масштабов слона, то, наоборот, широтные начнут расти быстрее, иначе скелет не выдержит огромного веса). Поэтому от австралопитеков к нам и высота таза несколько подросла.
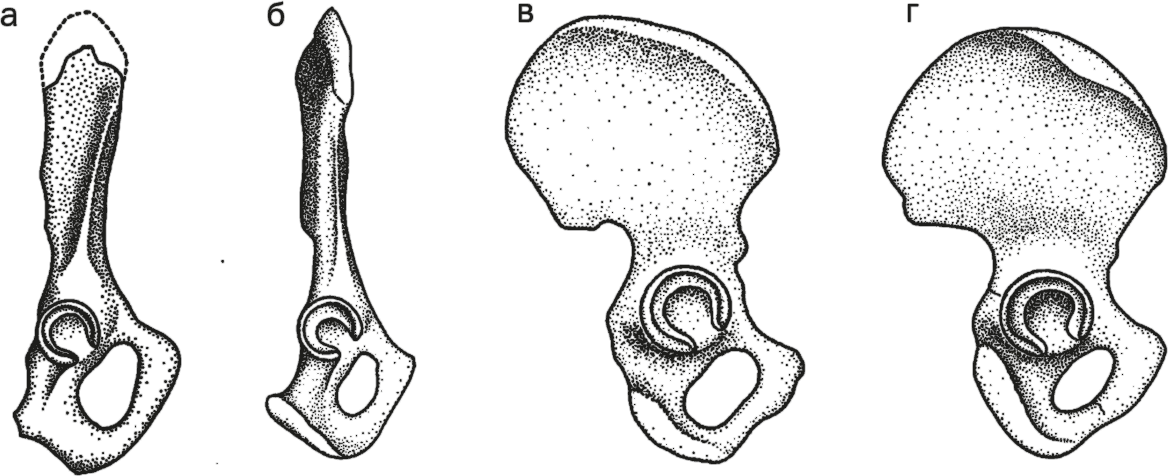
Рис. 2. Таз проконсула (а), шимпанзе (б), австралопитека (в) и человека (г).
Строение длинных костей ног отражает бипедию, наверное, лучше всего. У четвероногих приматов руки длиннее ног, колени разведены в стороны “колесом” и всегда полусогнуты, стопы разнесены друг от друга. Слабые мышцы не могут свести ноги вместе, да и форма нижних концов бедренных костей этому никак не способствует, туловище как бы “проседает”. Так как стопы при двуногом хождении оказываются далеко от центра тяжести, обезьяна компенсирует неустойчивость сильными боковыми колебаниями туловища, двигается очень неуклюже, враскачку, характерной походкой заправского бывалого морячка и того гляди норовит помочь себе руками. У прямоходящих ноги длинные, тазобедренные суставы сильно разведены друг от друга вследствие большой ширины таза, а колени сведены вместе, так что бедренные кости при взгляде спереди наклонены, а кости голени вертикальны, стопы сближены. Колени выпрямлены и при взгляде сбоку.
Более-менее бипедальное строение бедренных костей известно начиная с Orrorin tugenensis 5,88 млн лет назад.
Уголок занудства
У прямоходящих меняются места прикрепления и направления действия ягодичных и медиальных мышц, что очевидным образом отражается на рельефе как таза, так и костей ног. Новую форму приобретают верхний и нижний концы бедренной кости, а также межмыщелковое возвышение большой берцовой кости и поверхности голеностопного сустава. Шейка бедренной кости у человека длинная и ориентирована более вертикально; у обезьян она короткая и горизонтальная. Мыщелки бедра у человека повернуты относительно длинной оси кости, дабы колени могли сходиться, тогда как у обезьян они смотрят ровно вниз.
Большая ягодичная мышца обезьян отводит ногу вбок, а не выпрямляет ее. Также у них слабы мышцы медиальной группы бедра, отчего обезьяна не может свести ноги вместе, когда стоит на двух ногах. У человека эта способность развита несравненно сильнее, поэтому и соответствующие мышцы могучи. Это ярко отражается во внешнем виде: у шимпанзе бедро заметно сужается к тазу, а у человека оно чем выше, тем толще. Мышцы медиальной группы присоединяются к шероховатой линии на задней стороне бедренной кости, отчего у обезьян эта поверхность почти ровная, а у человека линия обычно развита в виде мощного костного гребня – пилястра. Показательно, что при врожденных параличах пилястр у человека не формируется, поскольку мышцы не используются и не воздействуют на кости. Большая ягодичная мышца у человека расположена сзади и крепится на небольшом участке в верхней части бедренной кости – на ягодичной бугристости, а у шимпанзе и гориллы она охватывает бедро сбоку и крепится намного ниже и почти по всей длине диафиза бедренной кости.
Строение стопы – отличный показатель типа передвижения, хотя в ископаемом виде эта часть тела сохраняется плохо. Четвероногие обезьяны имеют плоскую стопу с длинными изогнутыми подвижными пальцами, приспособленными для цепляния за ветви. Особенно выделяется большой палец, сильно оттопыренный в сторону, способный хватать даже лучше, чем большой палец руки. Это отражается в большей длине его мышц и характерной форме суставов.

Рис. 3. Стопа шимпанзе (а), австралопитека (б) и человека (в).
У прямоходящих продольный и поперечный своды (подъемы) стопы выражены хорошо, что позволяет перераспределять нагрузку тела более равномерно: половина на пятку, половина на пять головок плюсневых костей. Если для простоты расчета прикинуть, что человек весит 100 кг, то каждой ноге достается по 50 кг, а своды делят их так, что на пятку и переднюю часть приходится по 25 – условно по 5 кг на каждую плюсневую кость (в реальности, благодаря поперечному своду, на первую приходится львиная доля, немало и на пятую, а на три средних совсем чуть-чуть). Без сводов же – при плоскостопии – все 50 кг будут ударять в одну точку – переднюю часть пяточной кости, куда нагрузка в норме вообще не должна прилагаться. Нога будет болеть. У четвероногой же обезьяны и без сводов на каждую ногу будет ложиться по четверти веса, при общем весе 100 кг – по 25 кг, как у нас на каждую пятку, что вполне терпимо.
У двуногих пальцы стопы прямые, короткие, способные легко отгибаться наверх; большой палец не отведен в сторону и малоподвижен, а его суставы плоские; все плюсневые связаны мощными поперечными связками, чтобы не расползались в стороны.
Превосходство над Винни-Пухом, или Как все же украсть мед?
Тва и эфе любят мед.
Почему? Кто поймет?
Только чтоб его достать,
Надо ножки им сгибать.
В журнале Национальной американской академии наук в 2013 г. вышла статья, посвященная способностям людей к древолазанию (Venkataraman et al., 2013). Исследователи наблюдали за тем, как забираются на деревья представители разных экзотических племен охотников-собирателей. Особое внимание было обращено на пигмеев Центральной Африки, которые в поисках меда (чего же еще?!) регулярно карабкаются на высоту до полусотни метров (в статье приводится удивительно точная цифра 51,8; интересно, как ее измеряли?). Правда, пчелы тоже нечасто селятся на такой верхотуре, так что в среднем пигмеям приходится лезть только до отметки 19,1 м, что, конечно, тоже немало. В необоримом стремлении к меду доблестные конкуренты Винни-Пуха регулярно повторяют его способ спуска, так что, например, у пигмеев ака на падения с деревьев приходится 6,6 % смертности. Заметно успешнее аэта с Лусона, у которых аналогичная цифра составляет 1,7 % (а может, на Филиппинах просто мед правильнее, пчелы ниже гнездятся или веток на деревьях больше?). Но всех превзошли угандийские пигмеи тва. Они не падают никогда! Видимо, любят мед больше всех. В качестве контроля были взяты соседи тва – бакинга, “нелазающие земледельцы”, как они определены в статье, а также филиппинские собиратели аэта и земледельцы манобо.
Для того чтобы разобраться, как сии феноменальные способности тва отражаются на их ножках, был использован метод ультрасонографии. Измерялись длина мышечных волокон в мышцах ноги и углы сгибания суставов ноги. Длина волокон у тва и аэта оказалась заметно выше, чем у бакинга и манобо. Голеностопный сустав у собирателей сгибается намного лучше, чем у земледельцев, почти столь же сильно, как у шимпанзе. Обезьяны при взбирании на деревья сгибают ногу в среднем на чуть больший угол, чем это уже опасно для связок нетренированного (“индустриализированного”, по терминологии статьи) человека. Пигмеи при древолазании тоже попадают в “зону риска”, но редко превосходят опасные для горожанина величины сгибания лодыжки.
Авторы исследования померили также целых шесть больших берцовых костей пигмеев из леса Итури в поиске неких костных особенностей, позволяющих им столь сильно сгибать ногу при древолазании и столь успешно доставать мед. Измерялась, правда, почему-то “передняя дистальная ширина” этой кости (классики остеометрии плакали бы на этом месте). График получился на удивление непоказательный: собиратели занимают на нем крайнее положение вместе с “мальчиком из Нариокотоме” и южноафриканскими грацильными австралопитеками, в средней позиции находятся земледельцы с восточноафриканскими австралопитеками, а противоположный край оккупирован шимпанзе и гориллами.
Выводом из всего этого стало утверждение, что способности к древолазанию не обязательно отражаются на костях, а могут иметь основу в мышечных особенностях. Отсюда уже следует заключение, что мы можем недооценивать способности древних гоминид (в частности, австралопитеков) к древолазанию, рассматривая лишь их кости.
Красивые выводы! Но…
Во-первых, кто бы сомневался! Ясно, что на костях отражается далеко не все. Во-вторых, выводы звучат так, как будто пигмеи вообще живут на деревьях, а шимпанзе постоянно ходят прямо. А ведь на графике средний угол сгибания ноги у пигмеев все же недотягивает до такового у шимпанзе. Невозможно промолчать и о методе определения угла: измерялись фотографии, сделанные со случайного ракурса, от точек, определенных фактически произвольно. Например, на приведенной фотографии точка на колене взята, несмотря на надетые штаны! А ведь для измерения угла крайне важна ориентация объекта и точка зрения. При минимальном смещении искажение будет в несколько градусов – сопоставимое с обнаруженными различиями. Да и количество измерений было крайне малым. Далее: в исследовании было задействовано всего 6 костей современных пигмеев, причем не тех, кого наблюдали вживе. Вовсе не факт, что те пигмеи, чьи кости были исследованы, лазали по деревьям (может, это были женщины, например). Вообще-то, давно известен, скажем, замечательный “комплекс положения на корточках”. Когда человек много сидит на корточках, на всех костях таза и ноги появляются характерные изменения суставных поверхностей. В принципе, при сидении на корточках тоже происходит чрезмерное сгибание суставов, как и при древолазании. Думается, что у всех пигмеев поголовно этот комплекс развит в полной мере, ибо стульев у них не замечено. Запросто может быть, что развитие “комплекса положения на корточках” заметно способствует усиленному сгибанию стопы при лазании по деревьям. Взятые для контроля “индустриализированные” люди, ясно, такого комплекса не имеют, так что их способности к сгибанию ног вовсе не показательны. Думается, если бы были исследованы йоги, циркачи или гимнасты, у них способности были бы не хуже пигмейских. А уж если бы под наблюдение попали студенты биофака МГУ осенью, в сезон созревания яблок вокруг биофака, думается, выводы были бы совсем иными. А если бы на костях рассматривалась не “передняя дистальная ширина”, а изучались следы прикрепления мышц и особенности суставных поверхностей, то, возможно, были бы найдены соответственные изменения, следы которых можно было бы прослеживать и на останках древних гоминид.
Что, собственно, доказано? Что у пигмеев повышена подвижность суставов? Это было известно уже в XIX веке, а некоторые предполагают, что и древним египтянам. Оно и не странно, учитывая малые размеры тела и образ жизни пигмеев. Захочется медку в тропическом лесу – и не так ноги согнешь, и не туда залезешь. Функциональные изменения организма никто не отменял, а они не отменяют генеральных адаптаций, локомоторных в том числе. Кстати, гибкость мышц и подвижность суставов прямо зависят от возраста, а этот фактор совершенно не учитывался в исследовании. Пигмеи в среднем живут очень недолго, так что с большой вероятностью сравнивались молодые пигмеи с более пожилыми представителями контрольных групп. В любом случае этот момент остался покрытым мраком.
Так что, несмотря на пафосные выводы, статья не может служить образцом научного исследования. А способности наших предков к древолазанию от всего этого вовсе не меняются.
Кстати: для проверки положений статьи я, в жизни не залезавший на деревья выше трех метров и при наличии аллергии к меду, без малейшей подготовки согнул ногу на фантастический угол, побив рекорд всех пигмеев и шимпанзе, а также, возможно, орангутанов и гиббонов! Это если верить измерению угла на фотографии…
Наш большой палец стопы малоподвижен, но его мускулатура в целом осталась прежней, обезьяньей, независимой от мускулатуры остальных четырех пальцев. Поэтому при необходимости человек может разработать хватательную способность до невероятной ловкости. Известен удивительный прецедент, когда безрукая от рождения женщина Пелагея Семенова подарила И. В. Сталину обшитый бисером чернильный прибор – ручку, пресс-папье, подставку под чернильницу, причем низала бисер она ногами.
В некоторых культурах отдельные виды деятельности предполагали придерживание чего-либо большим пальцем ноги, например древка копья при его шлифовании. От этого палец разрабатывался и торчал в сторону очень далеко. Когда в XIX веке высокопросвещенные европейцы в белых пробковых шлемах приплывали на экзотический остров, скажем, на Филиппинах и обнаруживали там людей со стопой “как у обезьяны”, они глубокомысленно рассуждали о примитивных низших расах, застрявших на пути очеловечивания. Однако ныне те же аборигены забыли про копья, ходят в китайских кроссовках и имеют вполне стандартные человеческие стопы. Функциональная гипертрофия – великая сила!
У Ardipithecus ramidus 4,4 млн лет назад уже присутствовали своды стопы, но пальцы были длинные и изогнутые (это типично вообще для всех австралопитеков), а большой палец мог отводиться далеко в сторону. В стопе Australopithecus anamensis около 4 млн лет назад, судя по строению большой берцовой кости, большой палец был малоподвижен. У Australopithecus afarensis 3–3,9 млн лет назад своды стопы хорошо выражены, большой палец мог слегка противопоставляться другим, но намного слабее, чем у современных обезьян, отпечаток ноги был почти как у современного человека. В стопе Australopithecus africanus и Paranthropus robustus большой палец был несколько сильнее отведен от других, а пальцы – очень подвижные, что позволяет не считать их нашими предками. У Homo habilis стопа уплощенная, без выраженного свода (впрочем, она известна лишь по одной комплектной находке), но пальцы прямые, короткие, а большой палец полностью приведен к остальным. Видимо, совсем человеческая стопа сложилась лишь у рослых архантропов, хотя тут у нас имеется досадный пробел в материалах.
Стопа невиданного гоминида
Сложности изучения и интерпретации морфологии стопы хорошо видны на примере исследования стопы из эфиопской местности Ворансо-Милле. Этот район исследуется сравнительно недавно, но уже успел порадовать антропологов ценнейшими находками. Чего стоит один только скелет KSD – VP-1/1 – самый рослый среди афарских австралопитеков! Здесь же найдены челюсти, совмещающие черты анамских и афарских австралопитеков. Среди 54 тысяч окаменелостей нашелся повод и для новой сенсации – фрагмент уникальной стопы BRT-VP-2/73 из локального местонахождения Бартеле-2 (Haile-Selassie et al., 2012). Сохранились плюсневые кости с первой по четвертую и четыре фаланги правой стопы. К сожалению, нет никаких других останков скелета, которые могли бы быть отнесены к этому индивиду.
Находка имеет датировку 3,4 млн лет – самый средний возраст для афарских австралопитеков. Однако морфология костей заметно отличается от известного для этого вида. Причем отличается в примитивную сторону и во многом напоминает состояние ардипитека, жившего 4,4 млн лет назад – на целый миллион лет раньше! Новоявленная стопа обладает странным набором свойств: поперечный свод был выражен, а продольного не было, плюсневые и фаланги не слишком длинны и изогнуты, но большой палец короткий, сильно отведен и имел хорошую хватательную способность, а при ходьбе не загибался вверх, тогда как другие пальцы – загибались. Одна черта оказалась, по-видимому, очень примитивной: II плюсневая короче, чем IV; такое соотношение встречается у мартышковых и проконсула KNM-RU 2036, тогда как у современных понгид, ардипитека, афарских австралопитеков и человека соотношение всегда обратное.
В целом авторы первоисследования отмечают, что стопа BRT-VP-2/73 была столь же приспособлена к древолазанию, как и у Ardipithecus ramidus, хотя отличий от этого вида, как и от Australopithecus afarensis, тоже хватает. Главным выводом стало то, что в Эфиопии 3,4 млн лет назад существовали как минимум два вида: один – хорошо изученный прямоходящий Australopithecus afarensis, а второй – почти неизвестный полудреволазящий-полупрямоходящий, возможно, мало изменившийся потомок ардипитека. К сожалению, классификация гоминид строится в основном на признаках черепов и зубов, так что дать название новому виду авторы не решились.
Но, как всегда, возникают вопросы. К сожалению, авторы статьи не сравнили новую находку с Stw 573 – “Маленькой Стопой” или “Синдереллой” (то есть “Золушкой”) из грота Сильберберг Стеркфонтейна в Южной Африке. А ведь эта стопа имеет массу сходств с BRT-VP-2/73 при датировке от 2 до 4,2 млн лет назад со средней в 3,5 млн лет назад – практически синхронной с датировкой Бартеле (последняя опубликованная цифра для грота Сильберберг как раз и есть 3,67 млн лет назад). При первоначальном описании Stw 573 особое внимание уделялось значительному отведению большого пальца и его хватательной способности (Clarke et Tobias, 1995), хотя в последующем этот вывод был поставлен под основательное сомнение (Lovejoy et al., 2009). Показательно, что разнообразные индексы и показатели отдельных костей BRT-VP-2/73 весьма близки к таковым южноафриканских находок из Стеркфонтейна и Сварткранса, хотя таких сопоставлений сделано очень мало. Может, Australopithecus africanus, более архаичные, нежели Australopithecus afarensis, мигрировали из Южной Африки в Восточную?
Также несколько удивляет, что в многомерные анализы не попали никакие ископаемые материалы, кроме самой стопы BRT-VP-2/73. Вследствие этого вывод о необычайном сходстве новой находки с гориллами может оказаться преувеличенным. Не так уж сильно BRT-VP-2/73 отстоит от современных людей, чтобы не оказаться похожим на австралопитеков. Представлен лишь один график метрического сопоставления, в котором есть данные об афарском австралопитеке, и то только в дополнительном приложении – там расхождение BRT-VP-2/73 и AL 333–160 действительно велико. В статье упор сделан на описательные признаки. А ведь потенциально материалы для сравнения имеются. С другой стороны, стопа афарских австралопитеков доселе была известна лишь по изолированным и не слишком многочисленным костям, самый полный комплект – AL 333–115 – включает меньше фрагментов, чем BRT-VP-2/73. Нет ли здесь некоторого раздувания сенсации? Может, афарские австралопитеки и не были в строении стопы так уж продвинуты, как до сих пор считалось?
Как обычно, в конце приходится ссылаться на необходимость новых находок, которые якобы прольют свет на все темные места. Пока тайн лишь прибавилось, в соответствии с классическими аллегориями про фонарик в темной комнате и увеличение границ непознанного при расширении области познанного. Горизонты познания безграничны… Хватит ли костей в эфиопской земле?
Строение рук связано с прямохождением косвенно. У высших человекообразных руки приспособлены к хождению по земле с опорой на фаланги согнутых пальцев, что выражается в расширенности и особой мощности средних фаланг и особой скошенности суставной поверхности лучевой кости. Также у разных приматов имеются многочисленные морфологические адаптации к цеплянию за ветви, включая варианты с редукцией большого или других пальцев или с превращением пальцев в единый “крючок”, как, например, у паукообразных обезьян, колобусов или гиббонов. У полностью прямоходящих гоминид руки не приспособлены к хождению по земле или лазанию по деревьям, руки короткие, фаланги пальцев прямые.
Черты приспособления к хождению по земле или лазанию по деревьям в строении рук есть у австралопитеков Orrorin tugenensis, Ardipithecus kadabba, Ardipithecus ramidus, Australopithecus anamensis, Australopithecus afarensis, Australopithecus africanus, Paranthropus robustus и даже Homo habilis. Видимо, только Homo ergaster окончательно избавились от древесного прошлого в своих руках.
Относительно происхождения прямохождения выдвинуто огромное множество гипотез. Про это написана не одна книга и безумное число статей. Двумя важнейшими и ныне чаще цитируемыми являются миоценовое похолодание и социальная гипотеза, но история науки помнит и иные.
Трудовая концепция Ф. Энгельса с позднейшими вариантами, широко известная и вошедшая во все советские учебники, связывает возникновение прямохождения со специализацией руки обезьяны для трудовой деятельности – переноса предметов, детенышей, манипулирования пищей и изготовления орудий. В вульгарном изложении прямохождение возникло для того, чтобы освободить руки для трудовой деятельности. После труд привел к возникновению языка и общества, “сделал из обезьяны человека”. Однако трудовая концепция сталкивается с непреодолимой преградой в виде фактов: по современным данным, прямохождение начало возникать не менее 6 млн лет назад, в полностью завершенном виде сложилось уже 4 млн лет назад, а древнейшие орудия из Гоны в Эфиопии имеют датировку в самом оптимистичном варианте 2,7 млн лет, а из Ломекви – 3,3 млн лет. Скорее уж наоборот, освобождение рук из-за прямохождения способствовало развитию трудовой деятельности, зачатки коей можно наблюдать даже у вполне четвероногих шимпанзе и орангутанов.
Социальные концепции предполагают возникновение прямохождения еще в тропическом дождевом лесу и связывают его не с климатом, а с изменениями поведения приматов. Наибольший вклад тут внес, несомненно, О. Лавджой (например: Lovejoy, 2009). Согласно его разработкам, прямохождение появилось в связи с особой стратегией размножения, из-за удлинившегося детства и ослабления межсамцовой агрессии. Гоминиды в течение очень длительного времени выращивают одного, максимум двух детенышей. Такие животные не могут себе позволить большую детскую смертность. Самка должна постоянно заботиться о своем ненаглядном чаде, она оказывается скована и ограничена в возможностях питания и защиты от хищников. Носить крупного детеныша трудно, надо придерживать его руками, что дает почву для возникновения прямохождения. Отцы, в свою очередь, должны защищать вынужденно малоподвижных самок, бросаясь в хищников разными предметами, а также обеспечивать матерей пищей, а приносить ее, конечно, удобнее опять же в руках. При этом, когда говорится “должны”, не имеется в виду, что кто-то объяснял им права и обязанности гражданина или что они сами догадались и прониклись важностью своего предназначения, просто группы, где самцы не защищали и не кормили самок, банально вымирали. Косвенным последствием такой заботы много позже стало использование орудий труда. Для того чтобы такая система работала, надо, чтобы самец был как можно сильнее привязан к самке, то есть возникает крен в сторону моногамии (хотя и у современных людей, даже при строжайших религиозных ограничениях, моногамия никогда не бывает поголовной). Это достигается со стороны самок – скрытой овуляцией, так что самец не может угадать, на какой стадии созревания находится яйцеклетка (да и самка порой не может); а со стороны самцов – ослаблением конкуренции, что выражается в редукции размеров клыков и ослаблении полового диморфизма. И ведь мы действительно видим уменьшение клыков с самого начала становления прямохождения!
Согласно О. Лавджою, прямохождение возникло еще в тропическом лесу, а в саванны переселились уже двуногие гоминиды. Карликовые шимпанзе, живущие в джунглях Конго, в зачаточной форме демонстрируют те свойства, что предполагаются для предков человека.
У социальной концепции есть два явных минуса. Во-первых, ее крайне трудно подтвердить объективными свидетельствами: фактически единственным обоснованием является оценка полового диморфизма, сделанная в основном по размерам клыков. Во-вторых, тогда как прямохождение связывается с изменениями поведения, остаются неясны причины изменения самого поведения.
Миоценовое похолодание уже сто лет является одним из основных объяснений развития бипедии. Суть концепции в том, что в середине и конце миоцена в результате глобального похолодания климата произошло значительное – в несколько раз – сокращение площадей тропических лесов и увеличение площади саванн. В новых условиях у многочисленных живших тогда в тропических лесах человекообразных обезьян наметились три варианта дальнейшей судьбы. Первый вариант – самый печальный, но которому последовало большинство, – вымирание: в это время исчезло подавляющее большинство гоминоидов. Второй – сохранение в более-менее неизменном виде в оставшихся лесах: потомками этих везунчиков стали современные гиббоны, орангутаны, гориллы и шимпанзе. Впрочем, часть даже тех гоминоидов, что пережили миоценовое похолодание, вымерла уже в более поздние времена, примером чего могут служить гигантопитеки. Третий вариант – переход части гоминоидов к наземному образу жизни в расширяющихся саваннах.
Вообще-то, в саванну можно выйти и на четвереньках, что успешно сделали павианы и мартышки-гусары. Но крупные человекообразные пошли своим путем. Жизнь в саванне диктует свои условия. Для начала, тут есть довольно высокая трава, в которой сложно ориентироваться и искать пищу. Кроме того, в траве могут прятаться хищники, а смотреть поверх нее удобно, вставая на две ноги. Этим часто занимаются самые разные животные – те же мартышки-гусары, суслики, газели-геренуки и другие вкусные и пугливые создания. Если же хищник уже близко, то его можно попытаться устрашить: прямоходящее существо имеет психологическое преимущество перед четвероногим, поскольку смотрит свысока и, таким образом, кажется крупнее и сильнее, хотя может и не иметь реального подтверждения своей мощи. Поднимание уровня взгляда для усиления впечатления на противника является универсальным способом среди всех позвоночных животных, поскольку в первом приближении работает принцип “кто выше, тот больше, а кто больше, тот сильнее”. Связь чувства превосходства с ростом ощущает на себе каждый; впрочем, с разных сторон, в зависимости от того, какой рост имеет он сам. Для приматов в саванне этот аспект мог иметь немаловажное значение.
Кроме того, экспериментально и на математических моделях доказано, что двуногое передвижение на большие расстояния со средней скоростью энергетически более выгодно, чем четвероногое: двигать надо в два раза меньше ног. Правда, это преимущество становится действительно преимуществом только в своем законченном варианте, а переходные от четвероногости к двуногости стадии оказываются энергетически проигрышными.
Важнейший момент – терморегуляция. Лесные приматы были животными дневными, тот же распорядок дня они перенесли и в саванну. Это по-своему выгодно, ведь все хищники активны в сумерках или ночью, когда можно залезть на дерево и спокойно почивать там, пока тебя ищут в траве. Тем более, двуногие приматы очень медленные, они не умеют резво убегать, а днем убегать и не надо. Кроме того, днем гораздо меньше конкурентов, можно не спеша кормиться, не отпихивая локтями посягающих на те же травки-зернышки. Но ведь отсутствие полуденной бурной жизни в саванне тоже сложилось не просто так. Сиеста нужна, чтобы пережить самое жаркое время и не спечься. Днем же надо как-то решать проблему перегрева. В саванне (а дело происходит в Африке близко к экватору) солнышко печет прямо в макушку, так что двуногий примат нагревается гораздо меньше четвероногого, под солнечные лучи попадают только его голова и плечи, а не вся спина. Ту же сложность можно так же эффективно преодолеть с помощью потения. Обезьяны, конечно, потеют и в лесу на деревьях, но без особого фанатизма, а в саванне это становится важным процессом. Но из-за пота слипается шерсть, а суточные перепады температуры в саванне бывают весьма ощутимыми; мокрая шерсть по ночам может привести к переохлаждению и простуде. Поэтому шерсть катастрофически укорачивается; сколь-нибудь существенное оволошение остается только на макушке, где припекает солнце, да на подмышках и в паху – для концентрации запаха. Редукция же шерсти создает новую сложность. В тропическом лесу длинная шерсть выполняет у обезьян две функции: во-первых, защищает от почти ежедневных дождей, а во-вторых, за нее цепляются детеныши. В саванне дожди идут только во влажный сезон, так что первое назначение шерсти теряет смысл. Со вторым сложнее. Хватательный рефлекс прекрасно сохранился и у современных детишек, это прекрасно известно всем родителям, у которых ненаглядные чада регулярно выщипывают клочки волос своими цепкими ручонками. Однако на волосах головы ребенка далеко не унесешь. Логично, что проще всего нянчить беспомощных малюток на руках. К этому добавляются те же факторы, что уже рассмотрены выше в социальной концепции: матери становятся медлительными и уязвимыми, тем более что саванна не лес, спасительные деревья тут растут далеко не на каждом шагу, добежать до них с ребенком на руках непросто. Опять же самцы должны кормить и защищать самок, опять же двуногим существам делать это гораздо проще.
Минусом гипотезы возникновения прямохождения в результате миоценового похолодания является факт, что некоторые из древнейших известных прямоходящих приматов жили в тропических лесах, а из современных приматов живущие в дождевых тропических лесах бонобо очень часто прибегают к прямохождению. Впрочем, палеоэкологические реконструкции – что дышло, какие данные учтешь, то и вышло.
Кстати, о пейзажах…
Одно из новейших исследований на эту тему, видимо, дает самую достоверную и масштабную картину (Cerling et al., 2011b). Изучение 1300 образцов ископаемой почвы из всех основных восточноафриканских местонахождений за последние 6 млн лет на предмет соотношения изотопов углерода показало, что Ardipithecus ramidus жили в весьма открытых местообитаниях, где кроны деревьев закрывали менее 60 %, скорее 20–40 % земли, а то и меньше. Иначе говоря, передвигаться только по ветвям в таком “парке” уже никак не получалось. Правда, ардипитеки держались, видимо, самых лесистых мест, но “самые лесистые” все же были далеко не такими густыми, как им, может быть, хотелось, а островки деревьев окружала и вовсе открытая саванна. Крайне интересно, что после 4 млн лет леса снова стали наступать, так что ключевой вид так называемых грацильных австралопитеков – Australopithecus afarensis – жил в более густых зарослях с покрытием кронами 40–60 %. Около 3 млн лет назад начинается новое сокращение лесов, так что первым представителям нашего рода Homo надо было опять осваиваться в саваннах.
Однако не всех исследователей удовлетворяют доводы “саванной” концепции.
“Гипотеза водной обезьяны” рисует совсем иную картину. Очень подробно ее разрабатывал, например, Я. Линдблад (Линдблад, 1991). Согласно ей, к прямохождению предков человека подтолкнули сложности жизни рядом с водой. Нельзя не признать, что в умеренном варианте “гипотеза водной обезьяны” имеет много сильных моментов. Действительно, находки восточноафриканских австралопитеков почти всегда обнаруживаются в водных отложениях, вероятно, они обитали недалеко от рек и озер и добывали в воде часть своего пропитания (впрочем, это может значить только то, что тут они просто хорошо сохранялись; кости ардипитеков и южноафриканских форм нашли свой покой в сухих местах). Древние гоминиды могли вставать на задние ноги, чтобы переправляться через водные преграды; примеры подобного поведения мы видим среди обезьян-носачей и горилл. В строении человека есть ряд признаков, свидетельствующих о значительной адаптации человека к плаванию и нырянию, в отличие от орангутанов, горилл и шимпанзе: положение волос на теле по направлению от макушки к ногам – по течению воды при нырянии, ориентация ноздрей вниз – для сохранения воздуха в носовой полости, способность задерживать дыхание, редуцированный волосяной покров на теле, неэкономное расходование воды организмом, что крайне нетипично для животных саванны, небольшие перепонки между пальцами. В конце концов, человек просто не боится воды, а многие жить не могут без купания! Это сильные стороны “водной гипотезы”. Проблема в том, что в крайнем варианте ее сторонников заносит чересчур далеко, вплоть до утверждений о происхождении человека от дельфинов или миллионах лет питания икрой, яйцами птиц, моллюсками и рыбой. “Водная гипотеза” – классический пример идеи, “в которой что-то есть”; она многократно подвергалась критике с разных сторон и в целом хуже согласуется с фактами, чем другие концепции, но некоторые ее положения нельзя игнорировать (например: Langdon, 1997).
Подводя итог, очевидно, что прямохождение возникло не по одной какой-то причине, а под воздействием целого комплекса условий и предпосылок.
Интересные параллели и аналогии человеческому прямохождению можно наблюдать среди многих современных обезьян. Лемуры-сифаки и гиббоны, будучи крайними древолазами, по земле передвигаются на двух ногах, ибо их руки уже не годятся для опоры о землю. Сифаки при этом скачут боком приставными шагами или подобно кенгуру. Мартышки-гусары и павианы, живущие в саванне, часто приподнимаются на задние ноги, чтобы сориентироваться или высмотреть опасность. Калимантанские обезьяны-носачи, обитающие в прибрежных мангровых зарослях, переходят затопленные участки на задних ногах, держа передние над водой, хотя группы, живущие в глубине острова, вообще никогда не слезают с деревьев. Похоже ведут себя гориллы в заболоченных джунглях долины Конго в Западной Африке: на сочных лугах, где нет деревьев, разрастаются травы, на них пасутся слоны и карликовые лесные буйволы. Гориллы же, как бы они ни были велики, встав на четвереньки, рискуют погрузиться в эти травы до ушей, а потому они часами ходят на двух ногах.
Интересно, что двуногость неоднократно возникала не только в самых разных группах животных – от динозавров и птиц до кенгуру и тушканчиков, – но и среди приматов. Так, из итальянских угольных шахт в Тоскане известны скелеты своеобразных обезьян Oreopithecus bambolii. Они удивительно хорошо были приспособлены для ходьбы на задних ногах, удобной в уникальном ландшафте – заболоченных кустарниковых зарослях, по которым ореопитеки передвигались, придерживаясь руками за нависавшие над топью ветви.
Итак, прямохождение возникло около 7 млн лет назад, но еще долго отличалось от современного варианта. Походка австралопитеков, вероятно, была несколько семенящей, с маленькой длиной шага. Некоторые австралопитеки и даже Homo habilis использовали и другие виды передвижения – лазание по деревьям, а некоторые, возможно, и хождение с опорой на фаланги пальцев рук. Полностью современным прямохождение стало только около 1,5 млн лет назад.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК