Теория главной копии
Теория главной копии
По мнению ведущего молекулярного биолога Харриса Бернштейна (Harris Bernstein) из Аризонского университета, половой процесс возник для починки генов. Первым намеком на это стало исследование особых мутантных дрозофил, не способных репарировать (исправлять) свои гены: оказалось, что и рекомбинировать (перемешивать) их они тоже не могут. Рекомбинация — перемешивание генов дедушки и бабушки в сперматозоиде или яйцеклетке родителя — важнейший этап полового процесса. Сломайте генетическую репарацию — разрушится и половое размножение.
Бернштейн заметил, что клетка в последнем и при репарации генов использует одни и те же молекулярные механизмы. Но он не смог убедить генетиков и экологов, что связь между починкой генов и половым процессом не ограничивается заимствованием вторым инструментария первого. Генетики согласны с тем, что молекулярные механизмы полового процесса сформировались на основе механизмов генетической репарации, но не считают, что его современная задача — починка генов. В конце концов, человеческие ноги происходят от рыбьих плавников, но сегодня они используются, чтобы ходить, а не плавать{35}.
На секунду переключимся на молекулы. ДНК — материал, из которого сделаны гены — длинная тонкая молекула. Она несет информацию, записанную простым алфавитом из четырех химических «оснований» — как хитрая морзянка, в которой есть два типа точек и два типа тире. Назовем эти основания «буквами»: А, Ц, Г и Т. Красота ДНК в том, что она состоит из двух цепей, и каждая буква одной цепи комплементарна соответствующей букве другой. Эти буквы располагаются друг напротив друга: А слипается с Т и наоборот, а Ц — с Г и наоборот (см. рисунок). Это значит, что есть простой способ копирования ДНК путем движения вдоль нити молекулы и сшивания второй нити по ее образцу — из букв, комплементарных буквам в первой из них. Последовательность ААГГТЦ на комплементарной нити превращается в ТТЦААГ: скопируйте ее еще раз — и получите оригинал. Каждый ген обычно состоит из нити ДНК и ее комплементарной копии, плотно сплетенных в знаменитую двойную спираль. Специальные белки репарации двигаются, находят поломки и чинят их, сверяясь с комплементарной нитью. ДНК постоянно повреждается солнечным светом и химическими соединениями. Если бы не белки репарации, она стала бы бессмысленной абракадаброй в мгновение ока.
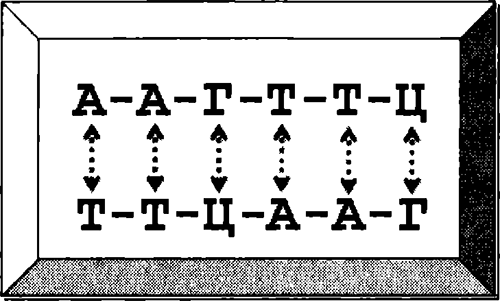
Схематическое изображение двойной цепи ДНК. Пунктирные стрелки соединяют пары комплементарных нуклеотидов.
Но что произойдет, если обе нити повреждены в одном и том же месте? Это происходит довольно часто — например, когда две цепи случайно химически спаиваются (это похоже на каплю клея на застегнутой молнии). В этом случае белки репарации не знают, какие буквы ставить на поврежденное место. Им нужен образец. Его предоставляет половой процесс, приносящий в организм копию того же гена из другого организма (скрещивание) или из другой хромосомы того же организма (рекомбинация). Теперь починка может идти по новому, неповрежденному образцу.
Конечно, и он может быть поврежден в том же самом месте, но это маловероятно. Продавец, расставляя ценники, убеждается, что ничего не перепутал, просто расставляя их еще раз. Он полагает, что вряд ли допустит одну и ту же ошибку дважды.
Репарационная теория возникновения полового размножения подтверждается и некоторыми косвенными фактами. Например, особь лучше переносит повреждающее ультрафиолетовое облучение, если у нее есть рекомбинация и если в ее клетках каждая хромосома существует в двух экземплярах. А вот когда в результате мутации какая-нибудь линия (группа близкородственных особей) теряет способность к рекомбинации, ее представители оказываются восприимчивыми к повреждению ДНК ультрафиолетом особенно сильно. Более того, теория Бернштейна объясняет некоторые моменты, которые не по зубам ее противникам: например, почему при образовании яйцеклетки в процессе мейоза клетка удваивает число хромосом, но потом избавляется от трех четвертей из них. Согласно репарационной теории, это вызвано необходимостью выявления ошибок и уничтожения их{36} [19].
Тем не менее эта теория неадекватна задаче, для решения которой она возникла — задаче объяснения полового размножения. Оправдывая существование рекомбинации, она не объясняет, зачем понадобилось скрещивание (вспомним, что мы разложили половое размножение на независимые этапы рекомбинации и скрещивания). Если половой процесс необходим для получения неповрежденных копий гена, то было бы разумнее пытаться получать последние от родственников — тогда инбридинг оказался бы страшно полезной штукой. Бернштейн говорит, что скрещивание — это способ «спрятать» мутации за нормальными вариантами генов. Но тогда просто переформулируем вопрос: почему инбридинг вреден, хотя, согласно Бернштейну, он должен быть полезен и, будь это так, мог бы считаться хорошей причиной для возникновения скрещивания.
Более того, гипотеза о рекомбинации как о способе починки генов — это, по сути, предположение о необходимости содержания запасных генных копий. Но ведь есть другой способ починки генов, гораздо более простой, чем случайный обмен между хромосомами. Он называется диплоидностью{37}. Яйцеклетка или сперматозоид гаплоидны — у них есть только по одной копии каждого гена. Бактерия или низшее растение — скажем, мох — тоже гаплоидны. Но большинство представителей флоры и почти вся фауна — диплоидны. То есть имеют по две копии каждого гена — по одной от каждого родителя. Некоторые живые организмы — особенно растения, получившиеся путем естественной гибридизации или в процессе искусственной селекции — полиплоидны. Гибридная пшеница, к примеру, в основном гексаплоидна: у нее по шесть копий каждого гена. У ямса женские растения октоплоидны или гексаплоидны, а мужские — тетраплоидны; это делает ямс стерильным. Ряд видов радужной форели, домашние куры и один примкнувший к ним попугай{38} триплоидны. У экологов даже сложилось впечатление, что полиплоидия у растений решает некоторые проблемы, возникающие в отсутствие полового размножения. А в некоторых отношениях может служить ему и альтернативой — допустим, на большой высоте или в высоких широтах.
Но мы рано заговорили об экологах. Не будем бежать впереди паровоза: пока все еще рассуждаем о генетической репарации. Если бы диплоидные организмы по мере роста тела «смотрели сквозь пальцы» на небольшое количество рекомбинации между хромосомами, то при каждом простом клеточном делении возникала бы возможность починки генов. Однако этого не происходит. Диплоиды рекомбинируют их только в момент специального, окончательного деления, называющегося мейозом и ведущего к формированию яйцеклетки или сперматозоида. Вот как Бернштейн объясняет однократность починки: во время обыкновенного клеточного деления существует другой, более экономичный способ репарации повреждений генов, который позволяет выживать самым приспособленным клеткам. На этом этапе починка не нужна, поскольку неповрежденных клеток скоро станет больше, чем поврежденных[20]. Проверка на ошибки{39} необходима только при образовании зародышевых клеток, встречающихся с миром один на один.
Вынесем вердикт репарационной теории: НЕ ДОКАЗАНО[21]. Молекулярный инструментарий полового процесса, похоже, действительно развился из механизмов генетической репарации — и рекомбинация в некоторой степени способствует починке генов. Но в этом ли смысл полового размножения? Вероятно, нет.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Обратная транскрипция — создание ДНК-копии по матрице РНК
Обратная транскрипция — создание ДНК-копии по матрице РНК За десять лет после открытия структуры ДНК и расшифровки генетического кода сформулированная Джеймсом Уотсоном в 1952 г. гипотеза об однонаправленном переносе генетической информации (от нуклеиновых кислот к
Что это за плазматические клетки, вырабатывающие антитела, и можно ли плазматическую клетку считать самой главной клеткой иммунной системы?
Что это за плазматические клетки, вырабатывающие антитела, и можно ли плазматическую клетку считать самой главной клеткой иммунной системы? — Что это за плазматические клетки, вырабатывающие антитела? О них уже знали во времена Мечникова или это более позднее
7 Расовая теория
7 Расовая теория Из всех способов уклонения от разбора характера социальных и моральных воздействий на разум человека самый распространенный — объяснение разнообразия поведения и характера человека наследственными природными признаками. Джон Стюарт Милл Совершенно
ТЕОРИЯ ФАНТОМНЫХ ОЩУЩЕНИЙ
ТЕОРИЯ ФАНТОМНЫХ ОЩУЩЕНИЙ Что все это означает? Ответ прежде всего зависит от мировоззрения конкретного человека. Некоторые воспринимают нематериальное тело как проявление души. Обычно оно оживляет физическое тело, но способно и действовать самостоятельно. Фантомно
8.1. Неодарвинистская теория эволюции
8.1. Неодарвинистская теория эволюции Фактически очень мало известно, если вообще когда-нибудь может быть известно, о деталях эволюции в прошлом. Также эволюцию нелегко наблюдать в настоящем. Даже на шкале времени в миллионы лет начало новых видов является редким, а родов,
ТЕОРИЯ, ЭВОЛЮЦИЯ И ТЕОРИЯ ЭВОЛЮЦИИ
ТЕОРИЯ, ЭВОЛЮЦИЯ И ТЕОРИЯ ЭВОЛЮЦИИ Термины эволюционируют помимо и вопреки нашей воле. Поэтому даже самое лучшее определение не может считаться окончательным. В прошлом теории (с оттенком пренебрежения) противопоставляли точному знанию, почерпнутому из Библии или, на
Теория моральных осцилляций
Теория моральных осцилляций Есть и другие причины, по которым стремление к половой морали — то ближе, то дальше от половой сдержанности — может самоподдерживаться. Если мужчины и женщины действительно созданы так, чтобы приспосабливать свои половые стратегии к
Теория игр и реципрокный альтруизм
Теория игр и реципрокный альтруизм Если бы случилось осуждать Дарвина за то, что он не создал и не развил теорию реципрокного альтруизма, то одним из доводов в его защиту был бы тот факт, что он — выходец из интеллектуально «неразвитой» культуры. В викторианской Англии не
3.2. Теория адекватного питания
3.2. Теория адекватного питания Кризис теории сбалансированного питания и открытие некоторые важных, ранее неизвестных механизмов (лизосомного и мембранного типов пищеварения, различных типов транспорта, общих эффектов кишечной гормональной системы); результаты
ОСТРОВНАЯБИОГЕОГРАФИЧЕСКАЯ ТЕОРИЯ
ОСТРОВНАЯБИОГЕОГРАФИЧЕСКАЯ ТЕОРИЯ Хотите узнать, как размер острова влияет на количество видов, обитающих на нем? Это легко сделать: поделите его пополам и наблюдайте за последующими переменами. Хотите узнать, как скорость иммиграции связана с расстоянием до материка?
27. Хромосомная теория наследственности
27. Хромосомная теория наследственности Вспомните!Что такое хромосомы?Какую функцию они выполняют в клетке и в организме в целом?Какие события происходят в профазе I мейотического деления?В середине XIX в., когда Г. Мендель проводил свои эксперименты и формулировал
2.2. Античная теория питания
2.2. Античная теория питания Античная теория связана с именами Аристотеля и Галена. Согласно этой теории, питание организма происходит за счет крови, которая непрерывно образуется из пищевых веществ в результате сложного процесса неизвестной природы, сходного с
Химическая теория
Химическая теория И тут на научном горизонте появилась химическая теория. Организм — это фабрика ядов. Таков был её главный тезис. «Мы отравлены с головы до ног продуктами наших собственных органов. Это — общий результат обмена веществ…» — писал в книге «Сон» И. И.
Теория Пенелопы
Теория Пенелопы У племени чероки существует обычай, пишет Леви-Брюль в своей книге «Первобытное мышление», по которому человек, видевший во сне, что он был укушен змеей, должен быть подвергнут тому же лечению, которое применяется при настоящем укусе змеи. В прошлом веке
Официальная теория
Официальная теория Личность человека, его сознание имеют своей природой химические процессы, то есть химические реакции и перемещение химических элементов в клетках головного мозга, называемых нейронами.Хорошо. Адепты официальной теории, вы можете предоставить для
Теория сексуальных стратегий
Теория сексуальных стратегий Начиная с 1972 г. зоологи стали широко и с большим успехом привлекать теорию полового отбора для объяснения феномена сексуальности у животных. Однако до конца 70-х годов XX века ее даже не пытались применять к анализу феномена человеческой