2.2. ЗРЕНИЕ
Обмен сильного зрения в сумерках на низкую остроту зрения при ярком свете позволяет собакам занимать экологическую нишу, недоступную нам.
P. E. Miller, С. J. Murphy (1995)
Зрение — это чувство, формируемое в процессе воздействия света на зрительный аппарат (глаз). Оно заключается в восприятии света фоторецепторами с преобразованием его в электрические импульсы, передаче их по нервным волокнам в головной мозг, где посредством анализа и синтеза пришедших сигналов формируется образная копия визуального объекта на определенном эмоциональном фоне. Свет обладает двумя физическими свойствами: интенсивностью и частотой. Они обусловливают его соответствующие психологические свойства: степень освещенности (яркость) и цвет. Благодаря зрительному восприятию за счет воздействия света объекты обладают следующими характеристиками:
— расположение в пространстве;
— размер, форма и состав;
— подвижность или неподвижность.
Материальную основу зрения составляет зрительный анализатор, состоящий из трех частей:
— воспринимающей — оптические, фоторецепторные, нервные и другие элементы глаза;
— проводящей — нервные волокна, нервы и нервные ганглии;
— центральной — четверохолмие (средний мозг), наружное коленчатое тело в таламусе (промежуточный мозг), лимбическая система, зрительная зона коры головного мозга (КГМ).
Для лучшего понимания специфики зрения собаки[1] за основу возьмем зрение человека, как наиболее изученное, а также волка (ближайшего дикого родственника собаки) и кошки как ведущей сходный с собакой образ жизни в связи с человеком.
Способность животных определять (распознавать) различные визуальные характеристики объектов обусловливается свойствами зрения, связанными прежде всего с особенностями строения глаза. Глаз — орган, воспринимающий световые раздражения. Он имеет оптическую (преломляющую), воспринимающую и проводящую системы. Первую составляют роговица, хрусталик и стекловидное тело, вторую — фоторецепторы и элементы глазного дна, третью — нейроны (наиболее связаны с фоторецепторами — биполярные и ганглиозные) и нервные волокна, выходящие из глаза в виде зрительного нерва. Вторая и третья системы входят в состав сетчатки — светочувствительной оболочки.
Пространство между роговицей и радужкой со зрачком — передняя камера. Пространство от роговицы до узловой точки хрусталика и от нее до сетчатки — передний и задний отделы: через узловую точку в середине хрусталика свет проходит по оптической оси без преломления, не искажаясь.
Глаз человека имеет эллипсоидную форму, слегка вытянут в переднезаднем направлении. Роговица выпуклая с преломляющей силой 43 диоптрии (Д), с большой разницей толщины между центром и периферией, с неглубокой передней камерой (2,7…3,0 мм), с более длинным задним отделом (17 мм), чем передний (6,5…7,0 мм). Узловая точка хрусталика далеко отставлена от сетчатки. В сочетании с небольшим зрачком это обусловливает попадание неяркого света на большую площадь в центре сетчатки, где он улавливается сразу большим количеством фоторецепторов, с малым рассеиванием по периферии. Последнее минимизирует расплывчатость изображения (аберрацию), повышая его четкость. Хрусталик относительно плоский, с небольшой преломляющей силой (19,8 Д), но с большой способностью изменять кривизну. Это позволяет глазу одинаково хорошо воспринимать разноудаленные объекты.
Глаз у собаки имеет форму, близкую к шару. Роговица с незначительной разницей толщины в центре и на периферии имеет изменчивые величины преломления (от 36 до 45 Д, в среднем 40,2 Д) и кривизны (от почти сферической до почти плоской, например у добермана и колли). Глубокая передняя камера (в среднем 4,2 мм), почти равные и небольшие размеры переднего и заднего отделов (7…10 мм) при большом диаметре зрачка (до 6…7 мм) способствуют максимальному проникновению света в глаз [12, 44, 94], что обеспечивает видимость при низкой освещенности. Придвинутость узловой точки к сетчатке и малый объем стекловидного тела (преломляющей среды) способствуют созданию на ней уменьшенного и сильно освещенного изображения. Но при этом свет из маленького центрального участка сетчатки, где он улавливается небольшим числом фоторецепторов, расплывается по всей ее поверхности, снижая четкость изображения.
У собак хрусталик большой, выпуклый (переднезадняя ось 4,0…7,8 мм). Как и роговица, он обладает большой преломляющей силой: 40,2 (35…45) и 32 (25…54) Д соответственно [8, 44]. Это сочетается с его слабой способностью изменять кривизну (слабая аккомодация — не более 2…3 Д) и малым объемом стекловидного тела. В результате свет фокусируется, как правило, несколько дальше сетчатки. Поэтому большинство собак слегка дальнозорки (гипер-метропия: +0,5 Д), т. е. они могут различать мелкие предметы или их детали на расстоянии не ближе 50…33 см; все предметы, расположенные ближе, кажутся расплывчатыми, в кругах рассеивания. В результате усиливается нечеткость изображения на сетчатке. По данным P. Kaufman (1992), у девятилетнего ребенка аккомодация повышается до 14 Д, что позволяет сократить критическое расстояние ясного видения до 7 см.
Исследование 240 служебных собак подтвердило в среднем их небольшую дальнозоркость (+0,25 Д). Однако за этим близким к норме (эмметропии) показателем скрывались значительные индивидуальные колебания в виде сильной близорукости (миопии) и явной тенденции ее развития с возрастом. По другим наблюдениям, близорукими оказались 53 % немецких овчарок (-0,5 Д и больше) и 64 % ротвейлеров. В то же время из-за строгого отбора среди собак-поводырей близорукость обнаружили лишь у 15 % немецких овчарок (С. J. Murphy et al., 1992). О развитии с возрастом близорукости свидетельствует и В. П. Давыденко (2000): «Собака в 8 мес узнает человека с расстояния 100 м лишь по походке, в 2 года это расстояние сокращается до 40 м, а к 8 годам она различает только близко расположенные предметы, становясь почти слепой. Все это необходимо учитывать при предъявлении собаке каких-то зрительных сигналов, объектов, особенно состоящих из мелких деталей, выбирая расстояние до нее не ближе и не дальше оптимального».
Еще одним нарушением световой фокусировки является астигматизм. Это когда различные участки оптической системы глаза не могут фокусировать свет одинаково, т. е. в одной точке. В результате изображение получается расплывчатое (аберрация), например в результате региональных нарушений кривизны роговицы или хрусталика. Астигматизм наблюдали у небольшого числа собак (С. J. Murphy et al., 1992): из 240 собак обнаружили астигматизм (-0,5…+3,0 Д) лишь у 10 из них. Е. П. Копенкин и др. (1998) сообщили о физиологическом астигматизме (-1,75…+2,0 Д) у большинства обследованных (22 собак). Это указывает на необходимость обследования служебных собак, выполняющих задачи, основанные на зрении, особенно связанные с жизнью людей, с целью выявления нарушений аккомодации (близорукость, дальнозоркость, астигматизм).
Итак, оптическая система глаза определяет особенности зрения собаки: способность ясно видеть при слабом освещении в сочетании с некоторой нечеткостью изображения при ярком свете, различные нарушения аккомодации. Еще в большей степени специфика зрения обусловливается свойствами сетчатки. Детально исследовали ее морфологию у собак давно: М. Schultze (1866); S. Ramon и V. Cajal (1894), и даже сравнивали I. Z?rn (1902) по этому признаку собак, охотящихся «по зрению и чутью». Оказалось, что сетчатка собак, схожая по структуре с таковой у других хищников, разделена горизонтально на две части: верхнюю — светлую, с сильной отражающей способностью и нижнюю — темную, наоборот, поглощающую свет.
Верхняя половина сетчатки покрыта отражающей структурой — тапетум (от гр. tapes — покрывало) (tapetum lucidum), а нижняя покрыта темным пигментом (tapetum nigrum). Верхняя половина сетчатки предназначена для восприятия слабого света, нижняя — для восприятия сильного света, который поглощается темным пигментом фусцином, предохраняя сетчатку от «засветки». Прошедший через фоторецепторы до отражающей мембраны свет возвращается, повторно проходя через них. Это позволяет собаке видеть не только при слабой освещенности, но и при разных ее уровнях. Правда, при этом отраженный свет, особенно сильный, рассеивается внутри глаза, снижая четкость изображения.
Работа обеих тапетальных частей сетчатки заключается в следующем. Начнем с верхней половины сетчатки. В физиологическом обиходе тапетумом принято называть только отражающую мембрану. Она имеет клеточную структуру [110], богата Zn и цистеином, содержит кристаллы гуанин-нуклеотидов в чистом виде [32] или в виде солей Са [44], окрашена в разные цвета от оранжевого через светло-желтый до ярко зеленого «яблочного». По мнению С. J. Murphy (1993), цвет тапетума скорее обусловлен взаимодействием света с его физической структурой, чем собственным спектральным составом, т. е. цветом его пигментов. Копенкин Е. П. и др. (1998) предполагают, что цвет тапетума от бледно-зеленого до оранжевого зависит от цвета радужной оболочки: у светлоглазых собак тапетум более светлый, у темноглазых — более темный. Также замечено, что у светломордых собак глаза, если их осветить фонариком, отражают слабый красноватый свет, а у темномордых — сильный зеленоватый свет [8]. Однако у представителей породы немецкая овчарка, ризеншнауцер и ротвейлер не отмечена прямая связь между цветом отсвета и окрасом морды (И. В. Егоренкова, 1999). Зато замечено, что переход отраженного из глаз света от красноватого к зеленому соответствует переходу темперамента от спокойного к возбудимому. Не воспринятый сетчаткой отраженный свет «выходит» из глаза в виде характерного свечения, отсвета [2, 12]. Причем он выходит точно в исходную точку вне глаза собаки, находящуюся в непосредственной угловой близости от глаза наблюдателя. В других направлениях этот свет глазом не отражается [8].
Цвет отсвета колеблется не только в зависимости от породы собак, но и внутри каждой из них [118]. Так, у грейхаундов и терьеров преобладает желто-зеленое отражение, у ирландских сеттеров добавляется оранжевое, которое у некоторых переходит в ярко оранжево-красное. В ряде пород, таких, как пойнтер, цвет отсвета сочетался с определенным волосяным окрасом, например, светло-оранжево-коричневый — с темно-коричневым окрасом. У одного трехцветного бульдога отражение света от тапетума было светло-голубо-серым с фиолетовым на периферии. Цвет тапетума желтый у евразиер и голубоватый у сибирской хаски, комбинированный желтый с зеленым краем у лабрадора и спаниеля, зеленый с голубым краем у цвергшнауцера, желто-оранжевый у чау-чау [110].
Существует мнение, что тапетум лучше развит у собак, преимущественно пользующихся зрением, и хуже — у мелких пород типа йоркширских терьеров и чихуа-хуа. Однако оно не всегда совпадает с данными по конкретным породам. Например, тапетум оказался плохо развит (занимал очень малую часть тапетального дна) у лхасо апсо, сибирской хаски и шелти (обе последние пользуются преимущественно зрением), хорошо развит у ретривера, сильно развит (занимает почти всю площадь тапетального дна) у австралийской черной трехцветной овчарки, лабрадора и пойнтера (две последние пользуются преимущественно чутьем).
Нижняя половина сетчатки, или нетапетальное дно глаза, насыщенного черно-коричневого цвета [118]. Однако этот цвет очень изменчив в зависимости от присутствующего пигмента и цвета радужной оболочки: темно-коричневый, серый или черный у евразиеров, сибирской хаски, но светло-коричневый, красно-коричневый и более красный у собак шоколадного или печеночного окраса со светло-коричневой или желтой радужной оболочкой, например у печеночных пойнтеров и английских спрингер-спаниелей, коричневого той-пуделя и шоколадного лабрадора. Слабопигментированное нетапетальное дно может иметь тигроидную окраску, например у шелти [110].
Казалось бы, такая ясная и упорядоченная морфологическая структура сетчатки у собак, по данным тех же исследователей, в действительности имеет отклонения. Отмечено [118], что тапетум на периферии и даже в центре не сплошной, а с вкраплениями темных участков нетапетального дна с ясными или размытыми краями. Возможны и другие варианты [110]: переход между тапетальной и нетапетальной зонами может быть постепенным (чаще у собак длинношерстных пород), а может быть резким, ясно очерченным (чаще у собак короткошерстных пород типа уиппет). Существуют и различные промежуточные формы. В некоторых случаях тапетум отсутствовал, например у шелти и австралийских овчарок — черной трехцветной и голубой мрамор (это у пастушьих собак, работающих по зрению!). Считается, что отсутствие или слабое развитие тапетума присуще «мраморным» породам: колли, шелти, бордер-колли, кардиган велш корги и арлекин. Известны случаи отсутствия не только тапетума, но и пигмента в нетапетальной зоне. В результате дно глаза красное (сосудистая сеть) — «рубиновый глаз», в частности у австралийской черной трехцветной овчарки и трехцветного бигля с бледно-желтой радужной оболочкой. Однако описанные виды морфологической структуры сетчатки нельзя считать специфически присущими собакам какой-то конкретной породы: могут быть самые разнообразные вариации. Например, у одних австралийских черных трехцветных овчарок отсутствовали тапетум и пигмент, у других отсутствовал только тапетум, а у третьих тапетум настолько сильно развит, что занимал практически всю площадь дна глаза.
Невольно возникает вопрос: имеют ли значение для реального зрения собак описанные отклонения от классической, морфологически ясной двухэтажной структуры сетчатки (сверху — светлая, внизу — темная) да и сама эта классическая структура? Ни B. Parry, ни G. H. R. R. Janssens не связали такие, казалось бы, естественные явления. Более того, по данным последнего, в «зрячих» породах по сравнению с «чутьистыми» тапетум оказался слабо развит или отсутствовал. Правда, в конце концов, он связал это с окрасом — мраморностью. P. E. Miller и С. J. Murphy (1995) приводят результаты электрофизиологических исследований М. S. Burns и др. (1988): оказывается, у биглей с наследственной дегенерацией тапетума светочувствительность мало отличалась от здоровых собак. У собак с отсутствием тапетума не обнаружены клинические изменения способности видеть в сумерках. Точно так же у животных с отсутствием пигмента в нетапетальной зоне, а потому с повышенным рассеиванием света внутри глаза клинически не проявилось значительное ослабление остроты зрения при ярком свете. Правда, по данным L. F. Rubin (1989), сильную светобоязнь наблюдали у австралийской овчарки в основном из-за внутриглазного рассеивания света в связи с полным альбинизмом (отсутствием светопоглощающих пигментов в глазу). Таким образом, по результатам исследований роль тапетума в зрении однозначно не определена.
Следующей структурой сетчатки, определяющей специфику и эффективность зрения собаки, служат фоторецепторы: колбочки и палочки. Они обладают разными свойствами, а потому по-своему формируют особенности зрения. У колбочек более выражено индивидуальное представительство в мозгу, чем у палочек. Это связано с тем, что отдельные нейроны сетчатки соединены с меньшим количеством колбочек, чем палочек. В результате каждое нервное волокно несет информацию в мозг от меньшего числа колбочек в сравнении с палочками [2, 87, 93], а значит, о более мелких деталях рассматриваемого объекта. Поэтому в палочковом поле уровень пространственной суммации выше, чем в колбочковом. Например, в сетчатке собак 1 колбочка или не менее 8…10 палочек соединяются с 1 биполярной клеткой, которых в 15…20 раз больше, чем ганглиозных. Кроме того, колбочковая активность после раздражения достигает «своей» ганглиозной клетки быстрее, чем палочковая [93], что тоже благоприятствует более детальному рассмотрению объекта. В то же время колбочки обладают низкой светочувствительностью в связи с медленным распадом их зрительного пигмента — йодопсина — на свету. Поэтому они могут работать только при повышенной освещенности. Отсюда понятно, что именно колбочки при высокой концентрации и достаточной освещенности обеспечивают остроту зрения — способность различать мелкие предметы или мелкие детали [2].
Палочки, наоборот, имеют высокую светочувствительность в связи с быстрым распадом на свету их зрительного пигмента — родопсина. Поэтому они могут длительно работать только при слабой освещенности. Но поскольку по одному нервному волокну поступают в мозг сигналы от многих палочек, то они не обусловливают остроту зрения. В результате предметы в сумерках воспринимаются расплывчатыми, хотя и могут быть видны далеко.
У человека сетчатка делится на две неравные части: маленькую центральную — центральную ямку, желтое пятно (fovea) и всю остальную, покрытую темным пигментом фусцином. Его назначение — поглощать свет, прошедший через фоторецепторы. Центральная ямка находится на пути прямого хода света, т. е. через узловую точку хрусталика, без преломления. В ней находятся только колбочки с очень высокой плотностью — 147 000 на 1 мм2 (Е. П. Копенкин и др., 1998). Причем не только 1 колбочка соединена с 1 биполярной клеткой, но и 1 биполярная клетка соединена с 1 ганглиозной клеткой, т. е. отношение колбочки и ганглиозной клетки составляет 1:1. Таким образом, у приматов центральная ямка — место наилучшего видения. Однако уже в 10 мм в сторону это отношение увеличивается до 16: 1. Кроме того, по мере отхода от центра сетчатки каждая колбочка окружается сначала одним кольцом палочек, потом все большим с одновременным уменьшением плотности колбочек. Так что у края сетчатки сосредоточены только палочки. В результате снижается острота зрения, но повышается способность видеть в сумерках. Одновременно уменьшается и плотность нейронов от центра к периферии сетчатки. Тем не менее общее их число велико, чему соответствует большое количество волокон в зрительном нерве — 1,2 млн.
У собак, волков и кошек центральная ямка отсутствует, а фоторецепторы рассредоточены в пределах овальной полосы. Она расположена поперек сетчатки перед тапетумом. Такая позиция фоторецепторов обусловливает возможность видеть в сумерках, но снижает остроту зрения за счет рассеивания отраженного от тапетума света. В овальной полосе, более короткой к виску, чем к носу, морфологически максимально благоприятные условия для контакта света с фоторецепторами: малое количество сосудов крупнее капилляров, а нервные волокна проходят от рецепторов к нейронам искривленно, ближе к ее краям.
Эффективность зрения собак определяется распределением фоторецепторов в сетчатке. Этот вопрос изучается давно, но мнения по нему неоднозначные. Впервые I. Z?rn (1902) сообщил о нахождении в центре сетчатки собак, охотящихся по зрению (немецкие беговые, русские псовые и т. д.), в отличие от охотящихся по чутью (некоторые терьеры), беспалочковой ячейки с повышенной плотностью колбочек. Однако G. L. Walls (1942) опроверг это с точки зрения целесообразности: животные с тапетумом не имеют такой ячейки, поскольку ее достоинства аннулируются рассеиванием отраженного тапетумом света. H. B. Parry подтвердил такое мнение в результате сравнения сетчатки у грейхаундов, охотящихся по зрению, и у сеттеров и ретриверов, охотящихся по чутью. Он не обнаружил центральную беспалочковую ячейку с высокой плотностью колбочек. Через некоторое время появилось сообщение о наличии на сетчатке центрального круглого пятна. Поскольку оно находится на оптической оси, то является местом наилучшего видения [2, 32]. Правда, авторы не указали, какие и сколько там фоторецепторов, хотя это обусловливает эффективность зрения. Через много лет после первого исследования фоторецепторов в сетчатке собак, охотящихся по зрению и по чутью (I. Z?rn, 1902), S. A. Koch и L. F. Rubin (1972) изучили распределение колбочек в сетчатке у 10 аляскинских маламутов, которые, будучи ездовыми собаками, ориентируются, используя зрение. Подсчет велся на участках сетчатки диаметром 50 мкм, содержавших не менее 10 колбочек: центральной, промежуточной (по другую сторону от головки глазного нерва, слепого пятна) и периферической (у носового края) зонах. В этих участках среднее число колбочек составляло 5,09 ± 1,47; 4,62 ± 1,38 и 3,86 ± 1,30 соответственно (р<0,01). Тем не менее считается, что разница между ними очень мала (сформирована близкими минимумам и максимумами по зонам: 1…10, 1…8 и 1…8 соответственно), что означает широкое распространение колбочек по всей сетчатке. Доказательством служит сохранение световоспринимающей функции при расширенном зрачке у таких же маламутов с различными поражениями центральной части сетчатки, включая атрофию обоих видов фоторецепторов.
Обнаруженное у собак большее число колбочек в центре сетчатки по сравнению с периферией не противоречит мнениям об отсутствии в ней беспалочковой центральной зоны, «населенной» колбочками, и об овальной полосе как ложе фоторецепторов и нейронов. Исходя из определяющей роли колбочек для остроты зрения и снижения зрительной функции в периферической части сетчатки, вполне естественно уменьшение их числа с удалением от центра. В то же время само количество колбочек в той или иной зоне сетчатки не указывает на связь с ними эффективности зрения. Значительно более информативны их относительное количество, соотношение с палочками и ганглиозными клетками, а также связь с мозгом. По данным G. Schubert (1950), у собак отсутствует центральная зона, как у человека, в которой каждая отдельная колбочка имеет собственную центральную связь. P. E. Miller и С. J. Murphy (1995) предполагают, что в центре сетчатки у собаки их менее 10 %, т. е. большая часть фоторецепторов представлена палочками, а отношение колбочек и ганглиозных клеток примерно такое же, как у кошки: 4:1 в центре (значит, связь колбочек даже здесь с мозгом не индивидуальна) и 20:1 на периферии. Полагают, что собаки по этому признаку ближе к кошкам, чем к приматам. В центральной зоне (25°) находятся преимущественно палочки; во всей овальной зоне отмечена высокая концентрация их пигмента — родопсина. Все эти данные указывают на невозможность у собак острого зрения. Этому же способствует и малое число ганглиозных клеток в сетчатке, с чем связано малое количество волокон (167 тыс.) в зрительном нерве. Таким образом, у приматов в центральной ямке только колбочки, а в зрительном нерве 1,2 млн волокон. Кстати [44], наличие 12…15 % колбочек во всей овальной полосе собаки еще не указывает на низкую остроту зрения. У человека их во всей сетчатке 5,2 % [2], но у него есть беспалочковая зона, состоящая из колбочек, расположенная в месте наилучшего видения.
Палочки, будучи значительно более многочисленными и чувствительными в сравнении с колбочками, расположены тоже по всей длине овальной полосы, т. е. в месте наилучшего видения. Это обеспечивает не только сумеречное зрение, но и точное определение дистанции и дальность зрения, особенно в связи с различением движущихся и неподвижных объектов. Так, собаки могут распознать движущийся объект с дистанции 810…900 м, но тот же объект в неподвижном состоянии они различают не дальше чем с 585 м. Вот еще несколько примеров. Л. Ф. Уитни (1997), подтверждая мнение, что движущиеся объекты вызывают реакции, которые не вызывает неподвижный объект, отмечал: одна собака могла распознавать движущийся объект за 914 м, а две другие — почти за 823 м. В то же время в его питомнике всегда находились собаки, способные разглядеть незнакомого человека за 137…183 м. Ё. Бергман (1992) наблюдал, что летом на острове таксы без труда отличали летящих за 200…300 м от них ворон от паривших на том же расстоянии сизых чаек. Эта способность выработалась у них благодаря тому, что он регулярно подкармливал чаек, а ворон, напротив, отгонял прочь. По другим данным, граница видимости движущегося предмета находится в пределах 250…300 м. В отдельных опытах хорошо дрессированная собака замечает движущийся предмет за 500…700 м. Очень развитую чувствительность к движению необходимо учитывать при общении с собакой, поскольку она легко замечает малейшие движения рук и глаз, изменения позы человека [89]. О том же свидетельствуют данные Э. И. Шерешевского (1970): на расстоянии 10 м собака замечает малейшие движения рук и глаз охотника, следит за движущимися предметами на расстоянии нескольких сот метров.
Рассмотрим роль ганглиозных клеток в связи с фоторецепцией. Их плотность во всей овальной полосе у волков не только высокая, но и менее изменчивая (12 000…14 000 на 1 мм2), чем у собак (6400…14 400 на 1 мм2) (L. Peichl, 1992). Это объясняется постоянными условиями существования волков (естественный отбор по остроте зрения, способствующей выживанию) и отсутствием селекции (искусственный отбор) по зрению. У собак с выведением самых разнообразных пород этот признак невольно становится все более изменчивым. В результате собаки стали различаться по строению сетчатки. Например, у немецких овчарок овальная полоса ясно выражена, высокая плотность ганглиозных клеток на большей ее части (волкоподобна), а у биглей овальная полоса слабо выражена, низкая плотность ганглиозных клеток (и очень неоднородная по этому признаку). Но даже среди биглей были отмечены различия. У большинства собак одной линии овальная полоса слабее выражена по наличию ганглиозных клеток, а крупные сосуды радиально сходились со всех сторон к центру через всю ее поверхность (звезда). У большинства собак другой линии овальная полоса сильнее выражена по тому же признаку, а крупные сосуды сходились к центру только сверху, пересекая ее лишь в двух направлениях; тем самым уменьшается помеха ходу световых лучей. В целом изучили недостаточное количество собак для выявления разницы по морфологии сетчатки между породами для охоты по зрению и по чутью. Правда, обнаружение у большого числа биглей (охотящихся по чутью) ясно выраженной овальной полосы свидетельствует об отсутствии больших различий между обеими группами собак по данному признаку [115]. Было отмечено, что отсутствует связь между изменениями зрения собак и морфологией тапетальной и нетапетальной зон сетчатки. Похоже, что и различия в морфологии овальной полосы как места наилучшего видения не влияют на зрение. Конечно, подразумевается, что фоторецепторная и оптическая системы глаз подопытных собак работают нормально. Так что эффективность зрения в разных его проявлениях, скорее всего, определяется характером распространения колбочек и палочек в овальной полосе, особенно в ее центральной части, и слабой аккомодацией.
Проанализировав морфофизиологические свойства глаза собаки, рассмотрим, как они реализуются в виде различных проявлений ее зрения. Особенности строения глаз собаки позволяют ей видеть в широком диапазоне освещенности, в том числе очень слабой, в сумерках. Для примера: у кошки минимальный порог световой чувствительности в 6 раз ниже, чем у человека (R. Gunter, 1951). У собак его не измеряли, но считают его несколько выше. Тем не менее еще И. П. Павлов указывал, что у собак способность анализировать низкую интенсивность освещения настолько хорошо развита, что экспериментально трудно определить ее границы. Е. П. Копенкин (1998) и другие исследователи объясняют низкий порог темнового зрения у собак большим числом палочек в сравнении с человеком и их расположением не только на периферии, но и в центре сетчатки, т. е. в месте наилучшего видения. Это значительно расширяет возможности использования собаки в разнообразных световых условиях.
Наличие фоторецепторов по всей овальной полосе, расположенной на горизонтальной оси сетчатки, обусловливает панорамность зрения, т. е. одинаковую его возможность по центру и на периферии. Такая способность усиливается за счет расположения глаз у большинства собак по бокам головы [12]. Поэтому оптические оси глаз расходятся латерально от осевой линии головы в среднем на 20° (Duke-Elader S., 1958) или на 20…50° [20]. В результате общий горизонтальный обзор двумя глазами, т. е. суммарное монокулярное зрение, достигает 240…250° (S. M. Sherman, 1975), или 250…260° [44], или 280° [20], что значительно шире зрения по вертикали. Все это позволяет собаке следить за горизонтально движущимися объектами без быстрого движения за ними глаз [8]; к тому же глазные мышцы у собак слабо развиты. Наверное, по этой причине при пристальном взгляде животного на быстро приближающийся предмет не наблюдается сколько-нибудь определенного схождения осей глаз (конвергенции). Подтверждением служит явление, когда собака, скосив глаза, внимательно разглядывает какой-то предмет у себя под носом, однако это скорее всего можно объяснить неспособностью хрусталика фокусировать изображение близко расположенного предмета в оптимальном месте сетчатки. Тем более при наличии слабой аккомодации у собак, которая действительно обусловливается слабой работой глазных мышц, управляющих формой хрусталика, а не тех, которые формируют конвергенцию.
В противоположность хорошо развитому боковому зрению (монокулярному) зрение перед собой (бинокулярное) у собак неширокое [12], в пределах З0…60° (S. M. Sherman et al., 1975), 60…90° [44] или 80…110° [20]. Это значительно уменьшает глубину воспринимаемого пространства, снижает протяженность точно определяемой дистанции до объекта. В результате собаки легко определяют расстояние только вблизи, что видно по их способности схватить быстро движущийся перед ними объект и преодолеть явные препятствия. По данным G. L. Walls (1963), у человека более узкое монокулярное зрение и более широкое бинокулярное: 180 и 140° соответственно [115].
В общем, угловое распределение зрительной информации у собак следующее (минимум — максимум для одного глаза): горизонтальное (боковое) зрение (относительно оси глаза) внутрь (к носу) — 30…45°, наружу (к уху) — 100…125°; вертикальное (фронтальное) зрение (относительно линии горизонта) вверх — 50…70°, вниз — 20…60° [94].
Однако по разным породам эти данные сильно расходятся (табл. 1). У собак с глубоко посаженными или выпуклыми глазами, расположенными спереди или по бокам головы, разное поле зрения [44].
1. Особенности зрения у собак некоторых пород с крайними морфологическими характеристиками (по Е. П. Копенкину)
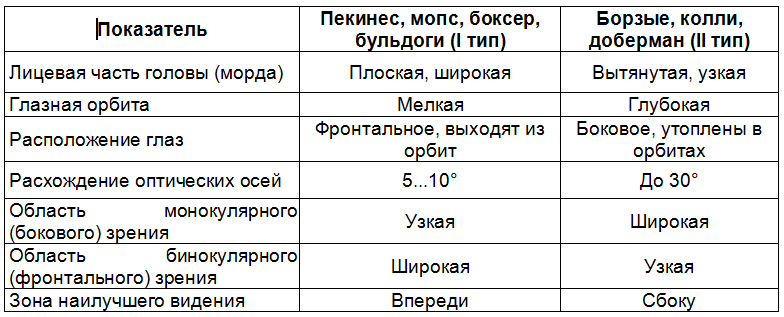
Разумеется, животные других пород с промежуточными морфологическими характеристиками обладают средним зрением. Все это необходимо учитывать при требовании от собак выполнения каких-то действий, связанных с их зрительными способностями. Например, животные I типа превосходят животных II типа по остроте зрения и точности определения дистанции близко перед собой, что связано с более развитой конвергенцией оптических осей, но уступают им по панорамности обзора и улавливанию подвижности объекта на большом расстоянии.
Еще одним показателем зрения является перспектива, которая обусловлена расстоянием от земли до глаз, что зависит от роста собаки. Разумеется, у низкорослых животных такая перспектива будет значительно меньше, чем у высокорослых. Можно сравнить крайние варианты пород по росту: брабансон, брюссельский и бельгийский гриффоны — 18…20 см и немецкий дог — до 80 см, ирландский волкодав — 86 см [82].
Более сложный вариант проявления зрения у собак — это способность к дифференцировке предметов, например геометрических фигур. Специальными исследованиями выявили достаточно высокое развитие у них этой способности. Так, в работе ученика И. П. Павлова М. М. Губергрица (1917) собаки отличали круг от эллипса с соотношением полуосей 8:9 [85]. В работе W. H. Karn et al. (1932) собаки быстро научились различать горизонтальные и вертикальные линии и более медленно — поставленный на основание и перевернутый на вершину треугольники. При этом не имели значения размеры объектов и были они контурными или закрашенными.
Интересную работу провели в одном из московских клубов служебного собаководства. Собаку научили садиться при виде круга, ложиться при показе треугольника вершиной вниз и стоять при показе стоящего на основании квадрата. Все фигуры были закрашены. Правильное выполнение визуальных команд началось после 2…3 предъявлений. И все-таки не обходилось без ошибок: даже после научения правильное выполнение составило в целом 87 %. Затем положение фигур стали менять, но с прежним выполнением визуальных команд: сидеть, лежать и стоять. Повернули квадрат, поставив его на вершину, а треугольники повернули вершинами вниз, влево и вправо. Повернутые треугольники собака узнала сразу, а перевернутый квадрат — со второго предъявления.
Затем те же опыты проводили с большими фигурами, а также предъявляли контурные круги и треугольники разного размера (ранее они были закрашены). Во всех вариантах правильное выполнение составило 100 %, в том числе при различении фигур по размерам. Потом показывали те же фигуры, но заштрихованные. Круг и треугольник собака узнала сразу, а квадрат — со второго предъявления. Далее провели опыт, являющийся смешением всех предыдущих: те же фигуры были разных размеров, положений и качества поверхности. В ответ на такую смесь вариантов число правильных ответов снизилось до 93 %. В следующей группе опытов изменили поведенческую реакцию собаки: научили подавать лапу при показе заштрихованного круга. При показе заштрихованного и не заштрихованного кругов она начала их правильно различать со второго предъявления. В предыдущем аналогичном опыте та же собака начала различать такие же круги, но с посадкой, сразу.
Далее опыт усложнили: на показ круга с вертикальной штриховкой собака должна была подавать правую лапу, на круг с горизонтальной штриховкой — левую. В результате она очень часто ошибалась; даже после научения правильное выполнение составило лишь 57 %. Отсюда можно сделать вывод: собака не различала наклоны штриховки, хотя в предыдущем опыте она успешно различала заштрихованную и незаштрихованную фигуры. Таким образом, собаки не только различают геометрические фигуры, даже очень похожие, но и узнают эти фигуры при изменении их характеристик, т. е. собаки способны отвлекаться от частностей, удерживая во внимании сущность. А это, как известно, составляет основу элементарной рассудочной деятельности, является проявлением мышления. Однако у такой способности оказался предел: отличение треугольников вершиной вверх и вниз; отличение квадратов в разных положениях; и совсем непреодолимое препятствие — различение одной и той же фигуры с разнонаправленной штриховкой. Кроме того, на успех решения задачи влияет способ ответного (сигнального) действия собаки, а также побочные характеристики различаемых объектов.
Все проявления зрения у собак предназначены для восприятия отдельных, хотя и взаимосвязанных, проявлений окружающего мира. Но все они обусловливаются обобщающей и самой сложной функцией зрения — его остротой, т. е. способностью ясно видеть мелкие объекты или их детали. Острота зрения — наименьшее расстояние между предметами, при котором они воспринимаются раздельно [20], или наименьший угол, под которым две точки видны раздельно [47, 50], а также наибольшая частота вспышек, при которой они не сливаются в сплошной свет. По данным электроретинограммы (ЭРГ), у анестезированных собак палочки способны раздельно улавливать вспышки с максимальной частотой 20 Гц. При более сильной освещенности, когда активизируются колбочки, слияние вспышек происходило при частоте 70 Гц. Например, 4 бигля, приученные нажимать педаль при вспышках, раздельно воспринимали их при более высокой максимальной частоте (до 80 Гц) и меньшей освещенности, чем при определении путем снятия ЭРГ [108]. У людей раздельное восприятие палочками максимальной частоты вспышек (70 Гц) примерно такое же, как у собак, а восприятие колбочками немного ниже (50…60 Гц).
Остроту зрения измеряют и другими методами. Один из них — метод Снеллена: он заключается в сравнении стандартного (постоянного) расстояния, с которого испытуемый ясно видит самые мелкие элементы, с расстоянием, с которого субъект с нормальным зрением, принимаемый за эталон, ясно видит то же. Его наиболее часто применяют в опытах с собаками, в частности, используют диаграмму Снеллена, в которой на экране меняют расстояние между темными и светлыми полосами до тех пор, пока испытуемый продолжает их видеть раздельно. В поведенческих опытах острота зрения у собак при сильной освещенности (37 лк) составляла в среднем 0,21. При определении у 3 биглей электропотенциала в коре головного мозга в результате освещения сетчатки острота зрения оказалась равной 0,235…0,308, а по ЭРГ — 0,40…0,43. Потом опыт усложнили. Горизонтальные полосы двигались с одновременным изменением расстояния между ними. Определяли минимальное расстояние, когда начинался нистагм (непроизвольное, ритмичное, пилообразное движение глаз в ответ на зрительные или вестибулярные раздражения). Острота зрения составила 0,2. Далее ЭРГ сравнивали по площадям сетчатки: 15° в центре и 15° тороидально вокруг него. Острота зрения в этих участках составила 0,143 и 0,085 соответственно (R. Orfi et al, 1993). Все эти исследования подтвердили низкую остроту зрения у собак (в среднем 0,267), так как при наличии у них панорамного зрения световоспринимающая способность сетчатки снижается с удалением от центра в связи с уменьшением количества фоторецепторов на периферии.
Японские исследователи определяли остроту зрения у трех собак породы шиба в поведенческих опытах. Их приучили к подкормке при виде обычного круга. Потом показывали круг Ландолта, т. е. с разрывом в каком-то месте окружности (в данном случае внизу), на который собаки не должны реагировать так, как на нормальный круг. При этом меняли расстояние до кругов, их диаметр, ширину окружности и разрыва в ней. За максимальную остроту зрения принимали ситуацию, когда круги правильно различались в 3 сеансах подряд из 21 на уровне не менее 70 % предъявлений (р<0,05). Острота зрения у собак оказалась равной 0,23; 0,24 и 0,33 [125].
Еще более естественные поведенческие опыты описывают P. E. Miller, С. J. Murphy (1995). Они определяли чувствительность сетчатки по угрожающей реакции собаки на движение руки поперек ее поля зрения или способности следовать за движущимся хлопковым мячиком. Так вот, в самых трудных экспериментальных условиях, когда острота зрения падала ниже 0,05, у собак еще сохранялись положительные зрительные реакции. Человека с таким зрением считают слепым. Отсюда следует обоснованный вывод о том, что различение мелких деталей для жизнедеятельности собак (даже рабочих) менее важно, чем для большинства людей. Сравнивали значение для реальной жизни собак способностей различать мелкие детали при ярком свете и видеть в сумерках. Низкая острота зрения у собак предопределена морфофизиологическими характеристиками глаз: малое число колбочек даже в месте наилучшего видения и их низкая светочувствительность и отсутствие связи каждой из них с центром (G. Schubert, 1950); большое число палочек с их значительной пространственной суммацией и потому низкой разрешающей способностью; слабая конвергенция оптических осей. И все это в сочетании со слабостью такого мощного корректирующего фактора, как аккомодация. Все вместе взятое усугубляется расплывчатостью отраженного тапетумом света внутри глаза.
Зрительные представления о любом предмете значительно расширяются и обогащаются, если помимо описанных выше его характеристик воспринимается цвет. Последнее возможно лишь через способность различать цвета, для проявления которой необходимо цветовое зрение (ЦЗ). Из всех чувств собаки наличие его остается наиболее спорным, хотя изучается более 100 лет. Это является следствием того, что несмотря на обычность и однотипность методов исследования ЦЗ (выработка дифференцировочного УР на подкрепляемые и неподкрепляемые цвета), получали неоднозначные результаты. Да и объясняли их не всегда адекватно и объективно. Поэтому до сих пор и нет единого мнения о наличии (или отсутствии) ЦЗ у собак. Одной из причин служит трудность проведения опыта в «чистом» виде. Изучаемые цветные объекты могут обладать совокупностью неуловимых для экспериментатора (но уловимых собакой) признаков: размеры и форма, подвижность, качество поверхности, а если используется свет, то яркость, температура, мигание и т. д., не говоря уже об окружающей обстановке и индивидуальных особенностях животных. Да и сам цвет может иметь разные насыщенность, оттенки. Поэтому выводы по результатам опыта иногда были ошибочными. Вот тому два примера, начиная с первого исследования, которое провел F. Graber (1884). Оно заключалось в том, что девять собак разных пород (половина — фокстерьеры) имели выбор свободного входа в светло-красную и темно-синюю камеры. Лишь две собаки (фокстерьер и пудель) чаще заходили в темно-синюю, чем в светло-красную. Остальные заходили в любую камеру, но когда яркость окраски камер выровняли, то «положительные» собаки одинаково часто заходили как в синюю, так и в красную камеру. Тем не менее собаки по-разному реагируют на объекты различного цвета, хотя совершенно очевидно, что это больше связано с интенсивностью окраски [74]. Еще раньше такое мнение высказали W. Nagel, F. Himstedt (1902): в большинстве случаев, где исследователи находят различение цветов, различная реакция животных может быть приписана именно разнице в степени освещенности, а не разнице в цвете [74].
Другой пример. Некоторые исследователи зажигали перед мордой собаки фонари с красным и синим стеклами. Причем в первом случае давалась подкормка, во втором не давалась. В результате легко вырабатывалась дифференцировка двух цветов. Однако при этом не учли, что красное стекло нагревается сильнее, чем синее, а разницу в температуре собака могла ощущать. Тогда перед фонарями поместили водяной светофильтр, поглощающий исходящую от них теплоту. В результате легко выработавшаяся дифференцировка цветов исчезла [76]. Следовательно, собака ориентировалась не на цвет стекла, а на его температуру. В других опытах подобные ошибки старались не допускать или, по крайней мере, учитывать, но тем не менее результаты исследований отличаются неоднозначностью. Поэтому вопрос о цветовом зрении собак до сих пор остается открытым.
Способность собак различать цвета изучали на основе слюноотделительных и поведенческих рефлексов. В качестве раздражителей применяли свет разных цветов и разноцветные предметы. Так, собаку с фистулой слюнной железы подкармливали одновременно с показом на экране красного квадрата от находящегося сзади фонаря [73, 74]. После 65…70 таких совпадений красный цвет без подкормки становился возбудителем слюноотделения. Затем на экране поочередно показывали квадраты зеленого, синего, фиолетового и белого цветов, но без подкормки. Тем не менее во всех этих случаях выделялось такое же количество слюны, как при показе красного квадрата. Если же после многократного показа фиолетового и синего или фиолетового, синего и зеленого квадратов показывали красный, который раньше совпадал с подкормкой, он тоже не вызывал слюноотделения. Однако при восстановлении возбуждающего действия красного квадрата показанный за ним зеленый квадрат опять стал вызывать слюноотделение. В этих опытах проявились иррадиации возбуждения и торможения (см. гл. 1). С уменьшением интенсивности каждого цвета путем увеличения расстояния экрана от собаки с 1 до 3 м, а также при показе черного квадрата после квадратов любого цвета повышалось количество отделяемой слюны. Следовательно, на слюноотделение действовал не столько цвет, сколько его интенсивность.
Недостоверные результаты были получены G. Nicolai (1907) при изучении на основе слюноотделения способности различать цвета у двух собак. В ответ на показ красной или зеленой чашки при различных степенях освещенности они очень скоро совершенно запутались, поэтому не удалось с несомненностью установить способность различения цветов собаками. На основании результатов обеих работ Л. А. Орбели считал, что собака не может реагировать на разные цвета различно, так как во всех случаях ее реакция определялась всецело изменениями в интенсивности освещенности.
В другом опыте Л. А. Орбели предлагал двум собакам с фистулой слюнной железы геометрические фигуры, различающиеся по направлению движения, форме, внешнему очертанию, качеству поверхности, сочетая или не сочетая их с подкормкой. Во всех случаях четко проявлялась дифференцировочная слюноотделительная реакция. Обобщая результаты этой и предыдущей работы по различению цветов[2], автор заключил, что ориентирование собаки в области световых явлений определяется тремя факторами (колебаниями интенсивности света, даже очень слабыми, движением и формой предметов) и что собака лишена такого могучего ориентировочного средства, как цветовая гамма. Потребовалось много времени, чтобы Л. А. Орбели (1938), став известным физиологом, академиком, пришел к выводу, что нельзя говорить, что собака не различает цветов, на том основании, что не обнаруживается разница в слюнном пищевом рефлексе[3]: «… разница между двумя моментами: способностью анализаторов более или менее тонко дробить внешние явления и способностью животного связывать отдельные детали с эффекторной деятельностью и проявлять различную деятельность под влиянием различных раздражителей. Можно хорошо различать раздражения, но не уметь различно на них реагировать».
Таким образом, было подтверждено наличие цветового зрения у собак, что вполне подтверждается преобладанием положительных результатов по его изучению.
Однако приблизительно в этот же период были опубликованы результаты исследований по выработке у собак поведенческих условных рефлексов на основе цвета: частично результаты были отрицательными, но в большинстве случаев — положительными.
Как пример можно привести две работы, где в качестве раздражителя тоже использовался свет разного цвета, но подкрепления были разные. В первой работе на переднюю лапу собаки наносили электроболевое раздражение при красном свете и не наносили при свете других цветов. После многократных сочетаний она отдергивала лапу только при зажигании красного света [13]. Во второй работе двух собак научили подходить к кормушке при красном свете различной яркости и не подходить при зеленом и синем свете. Было сделано заключение, что у собак несомненно существует способность различать цвета, но в отношении быстроты развития дифференцировки встречаются значительные индивидуальные различия.
В следующем опыте Л. А. Орбели (1913) позволял собаке подходить к корму при свете красного фонаря и не позволял подходить при свете сначала сине-фиолетового, потом зеленого фонаря. Различение красного и сине-фиолетового цветов вырабатывалось очень медленно и трудно, а различение красного и зеленого цветов вырабатывалось еще труднее. Но даже сформировавшееся различение было очень нестойким: нескольких дней перерыва в работе оказалось достаточно, чтобы оно исчезло. Правда, потом различение можно было восстановить и сделать это значительно легче, чем выработать вначале. Интенсивность любого цвета играла определяющую роль в скорости и правильности его выбора при решении о возможности или запрете подхода к кормушке. Исходя из результатов своих работ и противоречивых результатов других исследователей, автор пришел к заключению, что у тех собак, у которых обнаруживалось различение цветов, оно является рудиментарным, поскольку вырабатывается с большим трудом, требует значительной силы света и отличается нестойкостью.
В других исследованиях использовали разноцветные предметы, но применяли их по-разному. Первая серия опытов заключалась в предоставлении собакам возможности заходить в камеру или подходить к объекту, обозначаемому цветом, который сочетался с подкормкой, и не заходить в камеру и не подходить к объекту, обозначаемому цветом, который не сочетался с подкормкой. Во всех случаях расположение цветоносителей постоянно меняли (Е. М. Smith, 1912). Семь собак научили заходить в камеру с красной картонкой. Затем им предоставлялся свободный выбор входа в четыре камеры: на верху одной из них была прикреплена известная картонка красного цвета, на следующих двух — картонки других цветов или серые, а четвертая камера была без картонки. В конце концов животные в большинстве случаев входили в камеру с красной картонкой. Однако такое предпочтение проявилось только у трех собак (фокстерьер, вельштерьер и уиппет) и не проявилось у четырех остальных (шипперке, ирландский терьер и два фокстерьера). Причем это предпочтение легко затормаживалось при изменениях яркости и расположения цветных картонок. Надо отдать должное исследователю: и до, и после мало кто проводил подобные опыты на таком количестве собак. Был сделан вывод, что различение цветов собакой чрезвычайно нестойкое. Еще раньше А. Самойлов и А. Феофилактова (1907) указывали, что собака пользовалась способностью различать цвета (зеленый и серый) после обучения лишь при хорошо знакомых условиях. Если же поставленная ей задача чем-нибудь усложнялась, то она руководствовалась в своем поведении не цветом, а другими признаками предметов.
Аналогичный опыт провела В. К. Шепелева (1954), только «положительным» цветом (сигналом для захода в камеру) был зеленый, а «отрицательным» — красный, фиолетовый, синий и оранжевый. Для достижения правильных заходов (100 %) двум беспородным собакам потребовались 650…700 сочетаний зеленого диска с дисками других цветов. Такая наученность нарушилась заменой зеленого диска темно-зеленым и еще больше — светло-зеленым. Однако путем длительной тренировки восстанавливалась правильность заходов в камеры с новыми дисками. Во всех ситуациях длительность обучения у животных различалась.
По такому же принципу А. А. Бабушкин (2002) научил 10 собак разных пород подходить к чашке с зеленой карточкой (с одновременной командой «Зеленый!») и не подходить к чашкам с красной и синей карточками. Только при этом условии они получали подкормку и только из чашки с зеленой карточкой. Но даже через 8 нед правильность подходов составляла 30…90 %. При замене зеленой карточки светло-зеленой даже у собак с наибольшим исходным числом правильных подходов (77…90 %) увеличилось число ошибок. Путем длительной тренировки так и не удалось восстановить исходное число правильных подходов: в лучшем случае у трех собак из пяти оно достигло 76…87 %.
Подобное исследование провели на двух собаках породы шиба. Они научились подходить к кормушке с красной, синей или желтой карточкой (в этих случаях они получали подкормку) и не подходить к кормушке с серой карточкой [125]. Индивидуальные особенности собак здесь проявились не только в длительности тренировки для совершения правильных действий, но и в разной затрате времени для отличения каждого из трех цветов от серого. Например, для одной из них для отличения красного, зеленого и синего цветов от серого потребовались соответственно 3, 6 и 9 испытаний.
Вторая серия опытов заключалась в том, что собака должна выбрать из массы разноцветных предметов такой, который соответствует цвету, показанному экспериментатором. Впервые такие работы были проведены в начале XX в. W. Nagel, F. Himstedt (1902) научили пуделя брать с пола синюю, оранжевую или желтую карточку в ответ на показ карточек аналогичных цветов. Другой пудель научился выбирать красный шар в массе зеленых и синих, затем — шар любого цвета в соответствии с показанным экспериментатором. Но самое интересное заключалось в том, что необходимый цвет потом ему называли. Несмотря на длительность обучения (10 нед), был сделан вывод: у собаки можно выработать реакцию именно на определенный цвет, независимо от освещенности. К такому же выводу пришли А. Самойлов и A. Феофилактова (1907): продолжительными упражнениями можно заставить собаку различать цвета. В их эксперименте собака отличала зеленый цвет от серого разных степеней освещения. У В. Л. Дурова (1924) во время сеансов экспериментальной дрессировки немецкая овчарка приносила разнообразные предметы разных цветов и разной яркости в соответствии с показанным экспериментатором.
Трех спаниелей [120] научили выбирать красную пластиковую тарелку, из которой производилась подкормка, среди комплекта серых, из которых ее не подкармливали. Потом подкормка давалась из рук человека за выбор красной тарелки среди серых. После выработки прочного условного рефлекса серые тарелки поочередно заменяли голубыми, зелеными и желтыми. Позже они, тоже поочередно, служили «положительными» объектами, т. е. за их выбор среди комплектов других цветов собаки получали награду (из рук). Причем тарелки всех цветов имели разную яркость, но во время одного сеанса яркость тарелок всех цветов была одинаковой. В итоге все собаки безошибочно различали цвета любой яркости. Но выявились индивидуальные особенности: одна из них с трудом выбирала красный цвет в зеленом комплекте, другая — зеленый цвет в желтом комплекте.
И наконец, третья серия опытов заключалась в том, что собака при показе сигнального цвета, который выполнял роль цветовой команды, совершала достаточно сложные движения. Так, B. Л. Дуров (1924) демонстрировал способность немецкой овчарки при показе одного цвета пройти в одном направлении, при показе второго цвета — в другом направлении и т. д. В этой ситуации собака ошибалась чаще, чем при подаче предмета требуемого цвета, но преимущественно выбирала направление правильно. Продолжая свой опыт, А. А. Бабушкин отобрал 3 собак (из 10 исходных) с наилучшим различением цветов и прошедших общий курс дрессировки. Он научил их выполнять знакомые им команды: «Сидеть!», «Стоять!» и «Лежать!» при показе соответственно красной, зеленой и синей карточек. Число правильных выполнений команд составило в среднем 73,8 % (немецкая овчарка), 77,6 % (кокер-спаниель) и 86,7 % (доберман).
На обложке изображено выполнение этих навыков. Причем представлено в виде обычного двухфазного способа выработки условного рефлекса. Сначала подкрепляющий ранее усвоенный сигнал (голосовая команда) подается одновременно с обучающей командой (цветная карточка) или сразу после нее. Затем подкрепляющий сигнал не используется, и собака выполняет навыки только по предъявлении соответствующих им цветных карточек.
Несмотря на, казалось бы, подтверждение наличия цветового зрения у собак, остаются сомнения в их способности различать цвета. Так, С. А. Корытин (1984) считает, что собаки не различают цвета, а реагируют лишь на степень яркости. Наиболее одиозным примером служит высказывание финского зоолога и кинолога Ё. Бергмана (1992) по поводу результатов его соотечественницы A. Rosengren (1969): «Тот факт, что кокер-спаниели могли различать цвета, еще не подтверждает этой способности у других пород». Странность такого заявления заключается в том, что за 80 с лишним лет до работы A. Rosengren ни разу не была замечена породная специфичность цветового зрения. А вот высказывание американского ветеринара и кинолога Л. Ф. Уитни (1997): «Насколько можно судить по полученным учеными данным, цвета собаки не различают. У них не существует разницы между яркостью и собственно цветом. Одна группа ученых заключила, завершив исследования, что собаки не различают цвета, и если вообще реагируют на цветовые стимулы, то весьма слабо». Примерно этого же мнения придерживается его соотечественница Е. X. Вайтли (1997). Интересно, что все перечисленные авторы самостоятельно не изучали этот вопрос. Они использовали единственное обоснование, сформулированное еще Е. М. Smith (1912) в объяснение неоднозначных результатов своей работы. Она предположила, что различение цветов едва ли может играть ориентирующую роль в нормальной жизни собак. С ней согласился Л. А. Орбели (1913). Уже наш современник Ё. Бергман (1992), допуская вероятность восприятия собаками цвета, отметил, что он мало что для них значит и в их жизни не играет большой роли.
Однако работа В. К. Шепелевой (1954) показала, что подобное мнение, по крайней мере, не универсально. Ее дальнейший эксперимент заключался в следующем. С зеленым клапаном, закрывавшим вход в камеру, где две собаки получали подкормку, соединили тампон, смоченный в камфаре. После многократных таких сочетаний (цвет-запах) тампон перенесли на красный клапан, закрывавший вход в камеру, где подкормку не давали. При многократных повторениях собаки ориентировались только на зеленый клапан. То же самое происходило при замене камфары тополевым маслом в сочетании с зеленым клапаном. А дальше создали, казалось бы, совсем простую «обонятельную ситуацию». Тампон с тополевым маслом соединили сначала с одним из пяти красных, потом с одним из пяти зеленых клапанов (обычно входы в камеры закрывали клапаны разных цветов). Собаки проявили хаотическую реакцию. И в первое время число правильных подходов (на запах) составляло 10…30 %. Интересно, что и в этих ситуациях проявились индивидуальные особенности животных. Так что в некоторых ситуациях цвет для собак важнее, чем даже запах, который принято считать главным для них раздражителем. А отмечаемая всеми исследователями слабость цветового зрения объясняется в первую очередь спецификой морфофизиологических характеристик ее глаз. Был сделан вывод: собаки обладают цветовым зрением. Однако условный рефлекс на цветовой тон образуется у них чрезвычайно медленно, еще более затрудняясь при изменении его интенсивности. Но образовавшиеся временные связи на цветовой тон достаточно прочные. Собаки с трудом переключаются на ориентирование при помощи обонятельного или двигательного анализатора.
Обобщая информацию о способности собак различать цвета, уместно привести высказывание В. К. Шепелевой (1971) после успешного изучения цветового зрения у лесного хорька по той же методике, что и на собаках: «Видимо, распространенное среди экспериментаторов, изучающих наличие цветового зрения, мнение о весьма ограниченном числе видов животных, обладающих цветовым зрением, нельзя считать достаточно обоснованным ввиду крайне ограниченного количества соответствующих экспериментальных исследований и из-за ошибок в методических приемах. Невозможно, чтобы среди обширного класса млекопитающих со столь высокоразвитыми органами зрения и тонкой организацией нервной системы цветовое зрение имелось бы только у приматов и эволюция не шла бы по линии выработки такой значимой для существования животных способности, как различение цветов».
Следует заметить, что во всех работах по изучению цветового зрения даже не упоминались морфофизиологические особенности глаза собаки, хотя понятно, что именно они в первую очередь определяют цветовое зрение[4]. А ведь морфология сетчатки изучалась с середины XIX в., фактически одновременно с изучением цветового зрения. И даже в работах, посвященных колбочкам, фактическим цветоприемникам, их описывают в основном как часть сетчатки и менее как элемент глаза, обусловливающий остроту зрения. Поэтому сопоставим морфофизиологические характеристики глаза собаки с особенностями ее цветового зрения, т. е. попытаемся выяснить, почему оно такое, а не иное. Начнем с того, что в месте наилучшего видения (в центре овальной полосы) колбочек очень мало и они соединены по несколько штук с одной ганглиозной клеткой. Кроме того, они перемешаны с палочками (нецветоприемниками), которых даже здесь в 10 раз больше, чем колбочек, а на периферии сетчатки разница еще больше. Все эти факторы при низкой светочувствительности колбочек обусловливают слабое и нечеткое восприятие цветов, затрудняют их различение. Но и это еще не все «оппоненты» цветового зрения.
Панорамность зрения в сочетании с преобладанием палочек, имеющих высокую светочувствительность, позволяет собаке даже при характерной для нее слабой дивергенции оптических осей одинаково хорошо видеть спереди и по сторонам, замечать малейшие изменения обстановки и различия в форме, размерах, удаленности, освещенности, особенно подвижности и других нецветовых признаков объектов. Все это, конечно, отвлекает собаку от цвета и препятствует адекватному ответу на него. Незнание или неучет мешающих факторов в сочетании с непродолжительностью тренировки были причиной некоторых неудачных работ по изучению цветового зрения у собак [8, 115].
Однако у известного дрессировщика В. Л. Дурова собака при общении с ним отдавала предпочтение цвету предмета, а не форме. Например, при показе круглой фарфоровой зеленой сахарницы она приносила зеленые мячи, а при показе круглой лакированной коричнево-красной розетки приносила коричневые мячи. Причем последние были разного тона и лежали все вместе [60].
Приведенные свойства сетчатки глаза, конечно, обусловливают слабость цветового зрения, если колбочки охарактеризованы только с количественной стороны. А есть ли в них что-то, свойственное именно собакам, и какое это имеет значение? Были выявлены два типа колбочек с использованием антител к их наружным сегментам (К. Gropp et al., 1993) и с помощью ЭРГ-мигающей фотометрии (J. Neitz, G. H. Jacobs et al., 1989; G. H. Jacobs, 1993). Одни колбочки максимально чувствительны к свету с длиной волны 429…435 нм (для людей — фиолетовый цвет), другие колбочки — к свету с длиной волны около 555 нм (для людей желто-зеленый цвет). И хотя неизвестно, воспринимают ли собаки эти цвета, как люди, исследователи предположили, что видимый спектр у собак делится на два оттенка — фиолетовый и сине-фиолетовый (430…475 нм), который они, вероятно, видят как синий, и зеленовато-желтый, желтый и красный (500…620 нм), который они, вероятно, видят как желтый. Собаки, наверное, воспринимают узкую часть видимого спектра как бесцветную.
Цвета с двух концов спектра (синий и желтый), вероятно, воспринимаются ими наиболее насыщенными, а цвета промежуточных волн — менее интенсивными, как если бы они были смешаны с белым или серым. Поэтому в противоположность человеку, который обладает трихроматическим зрением и воспринимает волны видимого спектра как сотни отдельных цветов, цветовое зрение собак дихроматично со спектрально нейтральной зоной. На основании обнаруженной спектральной чувствительности колбочек американские исследователи предположили, какие цвета видят собаки.
Исходя из максимальной чувствительности одного из типов колбочек в средней части спектра был сделан вывод о неспособности собак различать свет средних и длинных волн (J. Neitz et al, 1989). Для подтверждения проводили сравнение их с людьми (дальтониками), у которых отсутствуют колбочки, воспринимающие зеленый цвет, и потому не различающих красный и зеленый цвета (красно-зеленая слепота). Во-первых, колбочки, максимально чувствительные к зеленому цвету, воспринимают и красный (620 нм), но в меньшей степени, чем зеленый. Во-вторых, и самое главное, многочисленными работами показана способность собак различать любые цвета, в том числе красный, любой яркости при соблюдении условий, соответствующих особенностям их цветового зрения. Возможно, на основании этих данных Е. X. Вайтли (1997) пишет, что, вероятно, у собак способность видеть цвет главным образом в спектре длинных волн света (красные оттенки) ограничена. Еще менее обоснованным представляется сформированное значительно раньше мнение, что собаки цветные изображения видят в серых тонах: желтый цвет воспринимается ими как светло-серый, оранжевый и синий — более темным, красный, зеленый и фиолетовый — как темно-серый [48]. Или что собаки видят большинство объектов серыми, будто полинявшими [12]. В целом произошла подмена понятия «различение цветов» понятием «видение цветов». Так вот, во всех работах изучали, как собаки различают цвета, а не какими они их видят. Можно выявить спектр поглощения света колбочками собаки, можно показать различение ею цветов, но нельзя сказать, каким каждый цвет видит собака (в нашем восприятии). Очень кратко это объяснил Г. И. Азимов (1958): «Животное смотрит глазами, а видит мозгом».
Помимо особенностей нервной системы, анатомо-физиологические характеристики глаз обусловливают еще одно свойство цветового зрения — индивидуальные особенности. Одной из причин может быть отклонение морфологии сетчатки от ее классической схемы — двухэтажной светло-темной структуры. Светлая, тапетальная, т. е. отражающая, зона может быть незначительной по площади вплоть до ее отсутствия за счет расширения темной, нетапетальной, т. е., наоборот, поглощающей зоны. Может быть и обратная картина, вплоть до отсутствия пигментного слоя (частичный альбинизм). Разумеется, в глазу создаются разные световые условия в связи с отражением, рассеиванием и поглощением света. В результате меняется эффективность работы колбочек как цветоприемников. Да и сами колбочки могут отличаться по набору зрительных пигментов, что неминуемо отражается на разном восприятии цветов у отдельных собак. Индивидуальные особенности цветового зрения могут обусловливаться также расширением панорамности зрения по мере смещения расположения глаз от осевой линии головы на ее боковые поверхности, т. е. расширением монокулярного зрения (крайние варианты: борзые, колли, лайки). Исходя из большей возможности вести боковой обзор, у собак этого типа более вероятно отвлечение на посторонние факторы во время отработки какого-то действия в связи с цветом по сравнению с собаками с противоположным расположением глаз — около осевой линии головы (крайние варианты: боксер, бульдоги, мопс, пекинес).
Таким образом, при общении с собакой следует использовать и цветовое зрение. Вероятно, в связи с цветом объекта у них можно вырабатывать не только простые условные рефлексы, но и более сложные формы поведения — навыки. Правда, при этом необходимо учитывать все особенности цветового зрения. Оно проявляется значительно слабее и менее обеспечивает соответствующую память, чем восприятие других характеристик объекта: размера, формы, удаленности и особенно подвижности. Решающую роль может сыграть и окружающая обстановка. Значит, малейшие изменения, даже незаметные для человека, могут отвлечь животное, помешать сосредоточиться именно на цвете. Следовательно, выработка каких-либо навыков у собаки в связи с цветом объекта требует многократных повторов при соблюдении неизменности других характеристик объекта и окружающей обстановки. Учитывая непрочную цветовую память животного, эти навыки придется периодически закреплять. С другой стороны, цвет объекта, в том числе детали одежды, может запомниться собакой независимо от воли и даже просто внимания общающегося с ней человека. Тогда не исключено, что цвет совершенно неожиданно сыграет возбуждающую или тормозящую роль при выполнении какой-то команды или в свободном поведении. Кроме того, необходимо учитывать возможные индивидуальные особенности собак в восприятии того или иного цвета. Их можно выявить только практическим путем и затем использовать в соответствии с замыслом дрессировщика.
Представления о зрительных характеристиках объектов вытекают из их образных копий, которые формируются в мозгу на основе информации, поступающей из сетчатки. Например, представление о дистанции. Каждый глаз видит один и тот же объект с разных сторон, где возможно наличие разных элементов. Поэтому его изображения на сетчатке обоих глаз будут различаться. Чем более выражены эти различия, тем ближе кажется объект, и наоборот, чем меньше этих различий, тем дальше объект воспринимается. Представление о дистанции формируется также на основе изменения взаимной направленности зрительных осей обоих глаз: их схождение (конвергенция) свидетельствует о близости объекта, их расхождение (дивергенция) свидетельствует о его удаленности. Представление о размерах и форме объекта создается в связи с площадью и формой раздражаемых участков сетчатки и оценкой его удаленности. Представление о движении объекта складывается из смещения его изображения по сетчатке и двигательных ощущений при слежении за ним.
Во всех случаях изображение объекта попадает на одноименные, соответствующие друг другу зоны сетчатки обоих глаз. В результате создаются условия для восприятия объекта как единого, без двоения. Эти условия реализуются в связи с прохождением информации от каждого глаза в виде нервных импульсов не только в свое полушарие, но и в соседнее. Оно осуществляется по двум перекрестным путям: на основании мозга (на уровне его среднего отдела) и между полушариями (мозолистое тело). В результате анализа и синтеза пришедших от обоих глаз сигналов в мозгу создается единый зрительный образ объекта, его единая образная зрительная копия, а значит, и единое представление о нем (бинокулярный эффект). Однако есть такой вид зрения, при котором образ объекта, воспринятого двумя глазами, отличается от образов объектов, воспринятых каждым глазом в отдельности. Это — цветовое зрение. Имеется в виду смешение мозгом благодаря перекрестным путям цветовой информации в виде нервных сигналов, пришедших от обоих глаз, один из которых воспринял один цвет, а второй — другой цвет. В результате оба глаза одновременно видят третий цвет как результат смешения нервных сигналов от двух исходных цветов. Например, одновременное смотрение обоими глазами на красный и желтый цвета формирует ощущение оранжевого цвета, смешение синего и желтого цветов дает зеленый цвет, смешение красного и зеленого цветов дает желтый цвет и т. д. Причем степень точности совпадения объекта и его образной копии обусловливается уровнем в ЦНС, где происходят анализ и синтез зрительной информации. Так, самая точная копия формируется в корковом зрительном центре.
При повреждении указанных перекрестных нервных путей, т. е. при анатомическом и функциональном разъединении полушарий, сигналы от каждого глаза поступают только в свое полушарие, не объединяясь. В результате в мозгу не формируется единый зрительный образ объекта, а только составляется представление о тех его характеристиках, на которые смотрит каждый глаз отдельно, т. е. в одном объекте видятся как бы два объекта (нарушение бинокулярного зрения).
В заключение следует рассмотреть еще одно явление — адаптацию. Будучи связанной с изменением освещенности, она представляет собой универсальный процесс, поскольку с ней связаны восприятия видимой характеристики объекта, любой качественной характеристики света. В целом зрительная адаптация заключается в изменении восприятия света в связи с изменением поступления его к сетчатке. Это осуществляется в результате изменений диаметра зрачка и чувствительности фоторецепторов: с усилением освещенности они уменьшаются, с ослаблением освещенности увеличиваются. Поэтому в первом случае восприятие света падает, во втором — возрастает. Как следствие данных процессов, при переходе из сильно освещенного пространства в слабо освещенное и, наоборот, из слабо освещенного пространства в сильно освещенное наступает временная слепота. Это связано с рассогласованием интенсивности распада зрительного пигмента на свету и его восстановления в темноте. С такими процессами напрямую связаны чувствительность фоторецепторов, их количество в деятельном состоянии и в конечном итоге восприятие света и возможность зрения. Так, при сильной освещенности распадается много зрительного пигмента. Его оставшегося количества достаточно для работы в этих условиях, но недостаточно для работы при слабой освещенности. Однако постепенно количество зрительного пигмента восстанавливается, как и связанные с этим процессы. В результате зрение при слабой освещенности становится возможным, т. е. произошла темновая адаптация.
При дальнейшем переходе в сильно освещенное пространство имеющееся к данному моменту большое количество зрительного пигмента обусловливает слишком высокую для этих условий чувствительность фоторецепторов. Они оказываются в состоянии сильнейшего перевозбуждения, значительно превышающего предел их работоспособности. В результате фоторецепторы становятся недеятельными. Однако постепенно избыточное количество зрительного пигмента разрушается на свету, а его оставшаяся часть достаточна для работы при сильной освещенности. Снизившиеся чувствительность фоторецепторов и их количество в деятельном состоянии стали соответствовать большой интенсивности освещения. В результате зрение при сильной освещенности становится возможным, т. е. произошла световая адаптация.
Необходимо отметить разную функцию отдельных фоторецепторов — колбочек и палочек. Зрительный пигмент колбочек — йодопсин — разрушается на свету и восстанавливается в темноте медленнее, чем зрительный пигмент палочек — родопсин. Это обусловливает меньшую светочувствительность колбочек по сравнению с палочками. Поэтому при сильном освещении больше работают колбочки, а при слабом — палочки. В определенной степени это подтверждается данными Г. Тамара (1976): при темновой адаптации ганглиозная клетка стимулируется как от палочек, так и от колбочек. В светлоадаптированной сетчатке ганглиозная клетка стимулируется только колбочковой импульсацией.
Таким образом, можно предположить, как протекает зрительная адаптация у собак. В связи с быстрой и сильной изменяемостью диаметра зрачка, центрально-продольной концентрацией палочек (в овальной полосе) и наличием светлой и темной половин сетчатки, что позволяет ей работать в широком диапазоне освещенности, адаптация у собак осуществляется быстро и полно. Но поскольку колбочек имеется значительно меньше, чем палочек (15 и 85 % соответственно даже в зоне наибольшей зрительной активности), темновая адаптация, скорее всего, происходит быстрее, чем световая. Это следует учитывать при работе с собакой в условиях резко меняющейся освещенности, принимая во внимание и какие-то ее индивидуальные особенности.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК