Атавизация
Изучение признаков гибридов позволяет установить одну очень важную закономерность, важную в теоретическом отношении, так как она дает указание на закономерность эволюционной диференциации видов, и важную с точки зрения практической, так как она позволяет до некоторой степени предсказывать свойства гибридов первого поколения.
Эта закономерность состоит в том, что гибриды получают известный атавистический характер, выражающийся прежде всего тем, что у них обыкновенно исчезают или резко ослабляются все более специфические признаки скрещиваемых видов, т. е. признаки, в эволюционном отношении более молодые. Хотя эта закономерность не строгая, тем не менее она достаточно широка дли того, чтобы с нею надо было считаться. Является ли она свойственной всем группам животных или в особой степени некоторым, пока судить трудно, но в некоторых группах (в частности у куриных) она выражена очень резко.
Относящиеся сюда факты следующие. Различные куриные имеют своеобразные и специфические головные украшения. Так, петух Gallus имеет на голове мясистый, более или менее зубчатый гребень и сережки; цесарка имеет вместо мясистого гребня костнокожный шлем, сережки иного строения, широкие ноздри; павлин имеет оперенную голову с оригинальным хохлом из перьев, снабженных маленьким опахалом на конце более или менее голого стержня; индюк имеет голову полуголую, как и цесарка, но над клювом свисающий, мягкий, способный раздуваться отросток и такие же кавернозные шишки на шее.
Все гибриды между названными формами имеют совершенно гладкую голову, более или менее нормально оперенную (см. рис. 70 и фронтиспис). Все перечисленные выше родовые признаки, являющиеся эволюционно молодыми, так как они свойственны только каждому из данных родов и несвойственны массе куриных, у гибридов исчезают. В силу этого голова гибрида с полным правом может быть названа не «промежуточной», а атавистической. Если даже у предков куриных голова и не была именно такой, то во всяком случае несомненно, что она не имела ни гребня Gallus, ни шлема Numida, ни, хохолка Pavo, ни шишек Gallopavo.
Гребень, свойственный Gallus, исчезает (точнее крайне недоразвивается) во всех изученных до сего времени гибридизациях с цесаркой, с павлином, с фазаном, с тетеревом, и у нас поэтому имеются все основания ожидать, что и при всяких других скрещиваниях Gallus он будет также исчезать. Исключением из этого естественно должны быть лишь скрещивания с другими Gallus, у которых гребень также имеется, как и у домашнего петуха. Но здесь уже следует ожидать исчезновения тех специфических отличий в гребнях, которые еще моложе, будучи свойственными лишь отдельным видам, или имеют более специфический характер. Действительно гибриды Gallus bankiva ? G. varius имеют гребень, но, насколько позволяют судить недостаточно точные описания, утрачивают видовые специфические признаки: глубокие вырезки G. bankiva, желтые пятна G. varius и пр.
Из всех куриных цесарка имеет особый и редкий серебристый тон оперения благодаря наличию лишь черного пигмента и отсутствию широко распространенного пигмента желто-коричневого. Так же своеобразен и рисунок цесарки в виде рядов округлых белых пятен. Можно было бы ожидать поэтому исчезновения этих специфических особенностей окраски у гибридов.
Действительно известные до сего времени гибриды цесарки с павлином, индюком и курицей имеют нормально развитый желтокоричневый пигмент и рисунок, в котором ряды округлых пятен заменены подковообразными рыжими полосами. Этот рисунок, подробно изученный Гайером, является характерно «атавистическим», так как он чрезвычайно широко распространен в различных группах куриных птиц и даже у более примитивных видов подсемейства цесарок-
Серебристый и золотистый фазаны оба имеют хохлы, однако совершенно различного строения. У серебристого фазана перья состоят из отдельных пушистых нитей черного цвета (при общем серебристом окрасе оперения), у золотистого — из крышеобразно сложенного опахала из крепкого стержня и рассученных бородок первого порядка золотистого цвета. Хохол гибрида состоит из нормальных перьев серебристой окраски, как преимущественно и все оперение, все специфические особенности строения перьев обоих родов (Gaenneus и Chrysolophus) нацело исчезли.
Подобные же факты можно найти и в других группах животных.
Так например в сем. лошадей зебры обладают очень своеобразной окраской из черных и белых полос. Если наклонность к полосам распространена довольно широко (черные ремни лошадей и ослов, крестообразная полоса на холке ослов, поперечная полосатость ног лошадей и ослов), то белая окраска взамен рыжей является у млекопитающих редким исключением. В соответствии с этим гибриды зебр с другими Equidae нацело теряют белую окраску и сохраняют лишь — иногда по всему телу, иногда местами — темные полосы. Возможно при этом, что древние лошади имели полосы более резко выраженные, чем сейчас, и их нынешняя окраска явилась уже приобретением последних полупустынных эпох. Расположение полос у зеброидов обычно напоминает наиболее примитивный тип зебры Греви (Dolichohippus grevii).
У гибридов як ? зебу исчезают специфические видовые признаки — бахрома по бокам тела яка и горб зебу.
Подобных фактов можно было бы привести еще много для того, чтобы показать, что они превалируют над противоположными. Однако и наличие этих противоположных фактов несомненно. Так например у цесарок несколько первых маховых перьев обычно бывают белыми (как и у павлина). Гибриды цесарки с курами тоже имеют белые первые маховые (Guyer, 1909). Гибриды цесарки с павлином имеют очень удлиненные кроющие хвоста (как у павлина, но может быть и у некоторых других куриных, в том числе у Gallus). Очень эфектно появление резкого, хотя и маленького белого пятна на плече у чрезвычайно далекого гибрида тетерев ? фазан (рис. 73). Совершенно такое же белое пятнышко имеется у тетерева, у глухаря и у межняка.
Блеск оперения вызывается, как известно, не пигментом, а структурными особенностями оперения. Так как эти особенности не одинаковы у различных видов и весьма специфичны, то неудивительно, что даже в том случае, когда оба родителя гибрида имеют яркий блеск, гибрид может оказаться матовым. В качестве такового Гайер приводит гибрида павлин ? курица (Guyer, 1909, 744) и различных других, произошедших от блестящих, но далеких форм, например фазан ? курица.
Несмотря на наличие подобных исключений — на наш взгляд они в самом деле являются более или менее исключениями — мы можем считать атавизацию гибридов действительной закономерностью, определяющей характер признаков гибридов. Какова природа этой закономерности? Действительно ли играют здесь роль «молодость» и «старость». признаков? Толкование этих явлений, как делают некоторые авторы в терминах «молодости» и «старости», граничит с явной мистикой. На самом деле, как мы думаем, вопрос здесь в том, что большинство специфических, вновь возникающих признаков оказывается связанным в большинстве случаев с накоплением рецессивных мутаций. В такой форме эта закономерность, наоборот, совершенно естественно освещается нашими современными генетическими знаниями. Действительно сейчас установлено колоссальным количеством фактов из разных областей животного и растительного мира, что рецессивные мутации возникают во много раз чаще настоящих доминантных. В любой популяции в скрытом виде, «как в губке», оказываются сохраняющимися разнообразные рецессивные мутации до тех пор, пока по условиям жизни вида та или иная из этих мутаций не окажется полезной. Тогда, подхваченная естественным отбором, она может начать быстро накапливаться и войдет в состав характерного генотипа вида. Конечно для естественного отбора, для качества приспособительного признака не имеет значения, обусловливается ли он действием доминантного или рецессивного гена, и поэтому решающим должен явиться вопрос, какая мутация будет чаще подпадать под действие отбора. Поскольку рецессивные мутации значительно. чаще и могут длительно сохраняться в скрытом виде до подходящего момента, постольку они чаще и будут подпадать под действие положительного отбора (некоторые имеющиеся здесь отличия между доминантами и рецессивами, например то, что доминанты могут подпадать под действие отбора непосредственно по их возникновении, а рецессивы — лишь после некоторого их предварительного стохастического размножения[37], — для данного вопроса решающего значения не имеют).

Рис. 73. Гибриды тетерев ? фазан (слева) и тетерев ? глухарь (справа) (ориг. с экз. Ганноверского музеи).
Поэтому для всякого вновь вырабатываемого признака больше шансов оказаться связанным с рецессивными мутациями, чем с доминантными. Вместе с тем понятно и то, что в определенном, различном для каждой группы форм проценте в образование новых признаков должны вовлекаться и доминантные мутации[38]. Тем не. менее роль рецессивных мутаций остается преобладающей, почему и атавизация при гибридизации является действительно закономерностью, отнюдь не устраняемой тем, что она захватывает фенотип не нацело, не на 100 %, а лишь в меру преобладания среди специфических признаков рецессивных над доминантными.
Закономерность атавизации гибридов таким образом является производной от закономерности преобладания в процессе эволюционной дифференциации рецессивных признаков, а эта последняя — производной от преобладания рецессивных мутаций над доминантными. Несомненно, что эта последняя является производной от еще более широких эволюционных закономерностей.
К сожалению специальных исследований, посвященных атавизации гибридов и теоретическому анализу ее, мы имеем очень мало. Помимо Гайера (Guyer, 1909) атавизацию исследовал Битинский-Зальц (1933) у гибридов-бражников, где например нередко возникает на переднем крыле особая линия, считаемая атавистической. Наследование ее прослеживается и в дальнейших поколениях, однако по. неясной схеме. Для объяснения этого наследования автор предлагает ряд соображений в духе Гольдшмидта, гипотетически допуская различные изменения порога раздражения для различных скрещиваний, в результате чего, во-первых, рецессивные гены усиливают свое действие до степени доминантных, в связи с чем, во-вторых, может ослабиться разница между доминантными и рецессивными аллеломорфами и т. и. Подробнее на этих теоретических построениях мы здесь однако останавливаться не будем, так как подкрепляющий их экспериментальный материал еще слишком слаб и к тому же вовсе не исчерпано самое естественное объяснение атавизации комбинаторикой генов по типу явлений атавизации в скрещиваниях пород домашних животных.
Чрезвычайно интересный пример атавизации гибридов мы находим в изученном Крыжановским (неопубликованно) скрещивании леща (Abramis brama) с карасем (Carassius carassius) — представителей подсемейств Abramidinae и Cyprininae. Кровеносная система в области жаберных дуг у гибридов оказывается резко отличной от обоих видов и несвойственной и другим костистым рыбам, в то же время она напоминает кровеносную систему других, более примитивных групп рыб — осетровых.
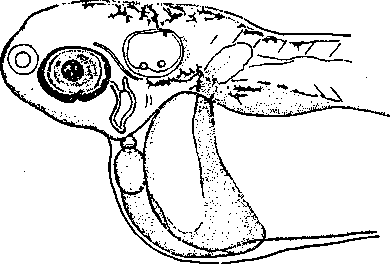
Рис. 74. Атавизация кровеносной системы у гибридов леща с карасем.
Правее и ниже глаза видны два сосуда — мандибулярная и гноидная дуги, соединенные комиссурой. По Крыжановскому, как и три следующие пояснительные схемы).
Рисунок 74 и схемы (рис. 75, 76, 77) поясняют этот поучительный случай. Более примитивный тип развития висцеральных кровеносных сосудов наблюдается у осетровых (Aciponser) (рис. 77). Сперва у них закладывается и функционирует лишь одна мандибулярная дуга аорты: Md(d) + Md(v). Затем появляются дорзальная половина гноидной дуги аорты Hy(d) и комиссура (сот), соединяющая ее с мандибулярной дугой аорты. После этого развивается вентральная половина гноидной дуги аорты Hy(v). У костистых рыб сходная картина развития наблюдается лишь как редкая индивидуальная вариация, отличаясь от типичной для Acipenser картины тем, что закладывающаяся дорзальная половина гноидной дуги аорты всегда более или менее рудиментарна (рис. 76). Обычно же у костистых рыб после мандибулярной дуги аорты появляются, только вентральная половина гноидной дуги аорты Ну(с) и комиссура (сот), соединяющая ее с мандибулярной дугой; дорзальная же половина гноидной дуги аорты совсем не развивается (рис. 75). Несомненно, что отсутствие Hy(d) у костистых рыб есть явление новое. У гибридов Abramis brama ? Carassius carassius всегда появляется Hy(d), нормально функционирующая вместе с вентральной половиной Hy(v), т. е. у них наблюдается более примитивное состояние, приближающее их к ганоидам (Крыжановский С. Г. ex letr.).
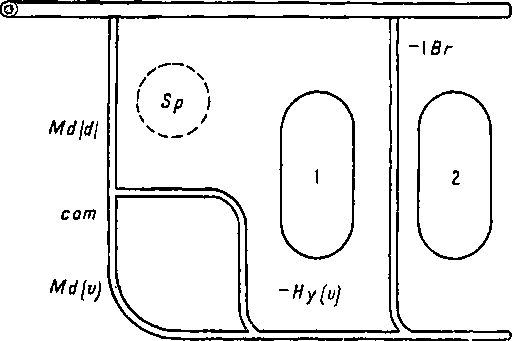
Рис. 75. Схема нормального развития сосудов костистых рыб.
Md (d) — дорзальная часть мандибулярной дуги; Md(u) — вентральная часть; com — комиссура, соединяющая мандибулярную дугу с гноидной, от которой осталась лишь вентральная часть [Hy(v)].
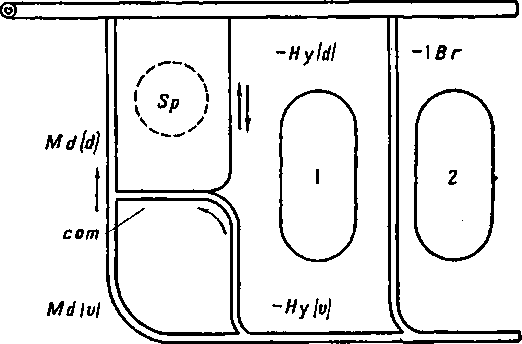
Рис. 76. Схема сосудов, встречающаяся у костистых рыб в качество редкой индивидуальной вариации; сохранилась слабо развитая дорзальная часть гноидной дуги [Hy(d)].
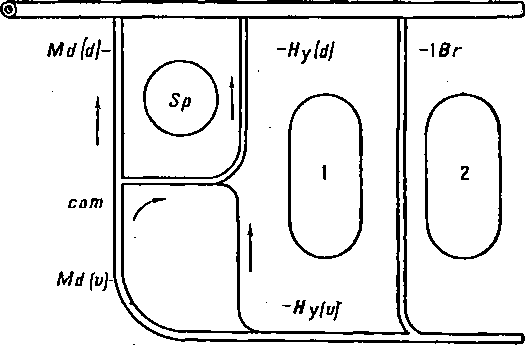
Рис. 77. Схема нормального развития сосудов у ганоидов (Acipenser): дорзальная часть гноидной дуги [Hy(d)] развита лучше вентральной [Hy(v)].
Этот случай является особенно интересным потому, что в свете генетического толкования атавизации он как бы указывает на большую древность расхождения названных подсемейств в ту эпоху, когда указанное примитивное строение должно было быть еще свойственно тем общим предкам, от которых развились и лещ и карась.
Дальнейшие исследования в этом направлении необходимы для окончательного разъяснения явления атавизации гибридов.
Очень интересен в этом же отношении изученный Ноаком (Noack, — 1908) череп гибрида льва и тигрицы, полученного Гагенбеком (Hagenbeck u. Aacken, 1897). Явление атавизации оказалось выраженным очень ясно в сильной редукции хищных зубов (гибрид оказывается менее хищником, чем оба родительские вида) и особенно в редукции мозговой капсулы. «Если бы, — замечает автор, — череп гибрида был найден в ископаемом состоянии…, то он был бы вероятно описан как новый вид и как более низко стоящая прародительская форма, из которой развились как львы, так и тигры и которая благодаря сходству с черепом ягуара являлась связующим звеном между крупными кошками Старого и Нового света» (Noack, 1908, s. 685).
Дальнейшую деталь вносят гибриды бражников Deilephila euphorbiae ? ? ? D. hippophaes. Окраска нижней поверхности крыльев у этих гибридов оказывается несвойственной обоим родительским видам, но зато очень напоминает окраску далеко живущих островных видов D. dahlii (Корсика и Сардиния) и D. tithymali (Канарские о-ва). Мори (Могу, 1901, s. 352) толкует этот случай как атавизацию, считая, что на островах сохранились более древние формы, континентальные же euphorbiae и hippophaes ушли в своей эволюции дальше. Именно в этом смысле следует оценивать и собранный Штандфуссом многочисленный ряд примеров доминирования у гибридов признаков «филогенетически более древних видов» (ср. Standfuss, 1898, Ex. Zool. st., S. 43 и др.).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК