Митозы гибридов. Элиминация хромосом
Переходя к вопросу о том, как происходят у гибридов митозы, в которых участвуют-хромосомы различного происхождения, цитоплазма одного вида и центросома, принесенная сперматозоидом другого вида, Приходится отметить, что у гибридов митозы могут итти и совершенно правильно и в высшей степени неправильно. При этом важно подчеркнуть, что правильность или неправильность митозов как будто ни в какой обязательной связи с отдаленностью скрещенных форм не стоит. Достаточно-таких примеров: митоз у гибрида двух морских ежей Strongylocentrotus ? Sphaerechinus идет совершенно, неправильно, а митоз у ультрагибрида Strongylocentrotus ? Antedon (морская. лилия) идет совершенно правильно (Бальцер, 1910). Мало того у реципрокных гибридов Strongylocentrotus ? Sphaerechinus митозы идут совершенно различно. Когда отцом является Sphaerechinus, митозы идут неправильно, а когда отцом является Strongylocentrotus — правильно. Во всяком случае необходимо подчеркнуть важнейший факт, что в случае гибридов Strongylocentrotus ? Antedon, т. е. морской еж ? морская лилия, совершенно правильно могут итти митозы, в которых участвуют хромосомы, принадлежащие животным разных классов, и что неправильные митозы (если говорить пока о митозах соматических) «оставляют не такое частое явление. Нужно впрочем иметь в виду, что вопрос этот изучен у животных еще совершенно не достаточно.
На материале морских ежей, изученном Бальцером (1910), можно хорошо познакомиться с неправильностями гибридных митозов и с проблемой их причин. При опытах гибридизации 4 неаполитанских морских ежей, Echinus, Strongylocentrotus, Sphaerechinus и Arbacia[40], выяснилось, что неправильные митозы возникают вполне закономерно, именно только тогда, когда отцовским видом является Sphaerechinus (рис. 94 и 47), но зато во всех случаях, т. е. во всех трех скрещиваниях Sph? ? ?Str, Sph? ? ?Ech и Sph? ? ?Arb. Во всех остальных скрещиваниях митозы идут нормально или почти нормально.
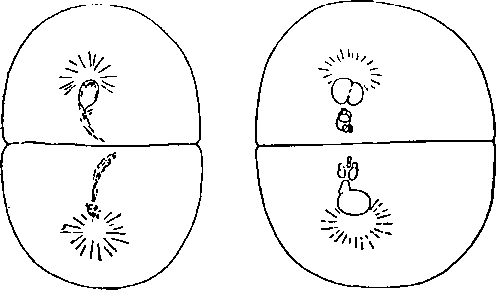
Рис. 94. Элиминации хромосом у Str. ? ? ? Sph. (слева) и Arb.? ? Sph. ? (справа). (По Бальцеру.)
Неправильности митозов выражаются в том, что начиная с метафазы наблюдается отставание некоторых хромосом. В то время как одни хромосомы успели разделиться и разошлись к полюсам, часть хромосом обнаруживает неспособность разделиться и либо они остаются на экваторе, либо, оторвавшись от одной из двух дочерних нитей, отходят с опозданием к одному из полюсов, либо, вовсе оторвавшись от веретена/остаются вне дочерних ядер, сохраняя в некоторых случаях способность к размножению, как было описано в главе VII. Рис. 47 представляет картину таких неправильных митозов. В результате этих неправильностей происходит элиминация хромосом, выпадение их из дальнейшего участия в жизни ядра, выпадение в цитоплазму с последующим разрушением.
Бальцер с доступной тщательностью изучил вопрос о том, 1) сколько хромосом подвергается элиминации и 2) какие именно хромосомы элиминируются.
Гаплоидные числа хромосом ежей следующие:
Strongylocentrotus purpuratus — 1.8
Echinus micrntuberrulatus — 18 или 9 (разные расы)
Sphaerechinus granularis — 20
Arbacia pustulosa — 20
У гибридов Sphaerechinus ? ? ? Strongylocentrotus при оплодотворении число хромосом должно быть 18 + 20 = 38. Однако уже после первого деления их остается гораздо меньше (в среднем из 41 подсчета лишь 21,4) и приблизительно такое же число сохраняется и в дальнейшем: после второго деления — в среднем 21,0 и в разных дальнейших стадиях, до бластулы включительно, насчитано в среднем 21,1. Во всех 14 культурах Бальцера картина получалась одинаковой: принимая во внимание трудности подсчетов, можно считать, что у гибридов Spherechinus ? ? ? Strongylocentrotus остается 21–22 хромосомы и соответственно 17 или 16 элиминируется. Таким образом замечательно, что остающееся число на 3–4 хромосомы больше гаплоидного числа Strongylocentrotus.
У гибридов Spherechinus ? ? ? Echinus, где число хромосом должно было быть также 38, их остается после элиминации тоже 21–22. Наконец у гибридов Spherechinus ? ? ? Arbacia должно было бы быть 20 + 20 = 40 хромосом, а после элиминации остается 22 хромосомы и элиминируется 18.
Какие же хромосомы элиминируются? Так как не все хромосомы могут быть узнаны точно под микроскопом, то этот вопрос может быть решен косвенно, несколькими путями: во-первых, можно проследить судьбу тех хромосом, которые могут быть всегда узнаны (например четыре самые длинные хромосомы Sphaerechinus); во-вторых, можно исследовать вариацию длины хромосом чистых видов и гибридов. Наконец можно сделать некоторые заключения по наблюдениям над многополюсными митозами (тетрасферами). Бальцер показал с высокой убедительностью, что элиминируются только хромосомы Sphaerechinus.
Таблица на стр. 253 показывает распределение хромосом по длине у чистых видов и гибридов.999 258
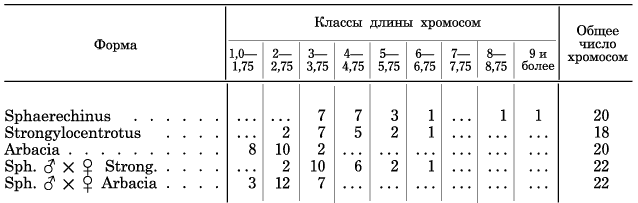
Длина хромосом определена в миллиметрах по рисункам при увеличении в 2270 раз.
Сравнивая распределение хромосом после элиминации у Sph. ? ? ? Str. с распределениями хромосом у родителей, видно, что 1) самые длинные хромосомы Sphaerechinus несомненно элиминировались и что распределение хромосом гибрида почти точно совпадает с распределением хромосом у Strongylocentrotus, 2) только в классах 3–3,75 и 4–4,75 имеется избыток на 3 (10—7) и 1 (6–5), т. е. на 4 хромосомы. Отсюда можно заключить, что у гибрида элиминировались все отцовские хромосомы кроме 4 из числа наиболее коротких. Это отвечает тому, что плютеусы Spherechinus ? ? ? Strongylocentrotus имеют чисто материнский характер с незначительными порой вариациями в отцовскую сторону (нужно помнить, что неизвестно, как долго сохраняются эти 4 отцовские хромосомы и не элиминируются ли они в дальнейшем).
Еще более убедителен гибрид Sphaerechinus ? ? ? Arbacia. Самые длинные его хромосомы не выходят за класс 3–3,75, как и у чистой Arbacia. Следовательно здесь с уверенностью можно говорить, что 13 более крупных хромосом подверглись элиминации и очевидно у гибрида остались лишь 2 самые короткие хромосомы Sphaerechinus.
Причины элиминации сейчас конечно еще совершенно неясны. Здесь возможны различные гипотезы. Исходя из факта, что элиминация в данной форме происходит только в том случае, когда хромосомы Sphaerechinus попадают в яйца других видов, ее можно сопоставить с более медленным темпом, свойственным дроблению Sphaerechinus. Так как элиминация начинается с того, что хромосомы Sphaerechinus отстают в процессе расщепления от хромосом другого вида, то можно истолковать явления так, что хромосомы каждого вида имеют свой собственный темп деления, отвечающий темпу дробления. «Медленные» хромосомы, попадая в компанию с «быстрыми» в «быстрое» же яйцо, естественно отстают, тогда как при реципрокном скрещивании отставания уже не будет, а будет, наоборот, некоторое забегание вперед, которое может не оказаться фатальным.
Такое объяснение встречает однако ряд трудностей. Так, темпы дробления Arbacia и Sphaerechinus повидимому совпадают. Также странно, что часть хромосом оказывается в состоянии перестроиться на новый тип дробления (ср. также благополучные скрещивания трех видов рода Fundulus, различающихся темпами дробления).
Также маловероятно, чтобы здесь какую-либо роль могли играть свойства центросом, например недостаток энергии для растягивания крупных хромосом и т. п. С другой стороны, Бальцер обращает однако особое внимание на свойства самих хромосом и в частности на их величину как на единственный пока признак, доступный более точному изучению. Анализируя только что приведенную таблицу, мы действительно видим, что как в случае гибрида Sphaerechinus ? ? ? Strongylocentrotus, так и Sphaerechinus ? ? ? Arbacia в элиминации хромосом обнаруживается очень простая закономерность — элиминируются все хромосомы, превышающие длину хромосом материнского вида. Для эмбриона, развивающегося из яйца Strongylocentrotus, хромосомы свыше 6,75 — предел длины хромосом самого Strongylocentrotus — оказываются уже чересчур длинными и не могут удержаться в митозах. Для яиц Arbacia, предел длины хромосом которого лежит уже при 3,75, «невыносимыми» оказываются уже хромосомы в 4. С другой стороны, для Strongylocentrotus оказываются «терпимыми» короткие хромосомы морской лилии Antedon несмотря на их чрезвычайную чужеродность. Таким образом можно предполагать существование некоторого нормального отношения, в котором должны находиться величина хромосом и окружающая их плазма.
Эти наблюдения для гибридагогии представляют конечно исключительный интерес, так как они, даже и не давая пока полного понимания явления, во всяком случае открывают возможность построения ряда рабочих гипотез, особенно учитывая возможности изменения длины хромосом рентгеном через транслокации. То, что величина хромосом оказывается действительно важным фактором, определяющим возможность сосуществования в общем правильном митозе хромосом различного происхождения — весьма правдоподобно. В этом отношении особенно поучителен конечно гибрид Sphaerechinus ? ? ? Arbacia, элиминирующий все хромосомы Sphaerechinus кроме 2 самых мелких, соответствующих по размерам хромосомам Arbacia.
Возможно ли сохранение в гибриде отцовских хромосом в неактивном состоянии? Этот вопрос с точки зрения хромосомной теории наследственности парадоксален, но мы считаем возможным отвести его обсуждению место ввиду наличия некоторых недостаточно изученных моментов в кариологии гибридов.
Выше излагался процесс элиминации отцовских хромосом при гетерогенном оплодотворении, как то было показано Купельвизером, Бальцером и другими, что сопровождалось возникновением в результате гибридизации чисто матроклинного потомства.
В некоторых случаях однако (Годлевский, 1906, Моррис, 1914) подобная элиминация не только не была доказана, но, наоборот, собранные наблюдения говорили в пользу ее отсутствия, причем личинки оказывались попрежнему чисто матроклинными.
Годлевский (1909) наблюдал в скрещивании морской еж ? морская лилия (см. выше, стр. 116) хромосомы, да и то не точно, лишь в первом дроблений. Однако при исследовании личинок уже на стадии гаструлы, протекавшей совершенно точно, как у ежей, он обнаружил величину ядер не только не вдвое меньшую, как должно было бы быть у гаплоидов, но как бы промежуточную между величиной диплоида-ежа и диплоида-лилии, имеющую ядра значительно более крупные.
Рис. 95 показывает эти размеры: рис. А показывает величину ядер гаструл у чистых Echinus, рис. С — величину ядер у Antedon, а рис. В — величину ядер гибрида, имеющего впрочем чисто матроклинный фенотип. Подобная же разница обнаружена и у плютеусов — «гибриды» имеют несколько более крупные, но более малочисленные ядра, чем у чистых Echinus. Годлевский на основании этого заключил, что хромосомы Antedon принимают участие в образовании ядер эмбрионов, хотя тем не менее у гибридов этой генерации признаков Antedon не обнаруживается.

Рис. 95. Величина ядер в гаструлах у Echinus (А) Antedon (С) и F1 (В).
Данные Годлевского в полной мере подтвердил Бальцер (1910). Им были оплодотворены спермой той же лилии Antedon яйца ежей Strongylocentrotus и Echinus (18-хромосомные) и прослежены митозы. Хромосомы Antedon (11 или вернее 12) — по крайней мере некоторые — оказались характерной двуплечей формы, не свойственной ежам (см. выше рис. 92), и на митозах ультрагибридов можно было совершенно уверенно наблюдать рядом хромосомы обоих типов. Никаких следов элиминации хромосом, комков хроматина в плазме и прочих неправильностей замечено не было, и на всем протяжении дробления число хромосом оставалось постоянным — 29–30.
Развитие гибридов по сравнению с контролем было сильно задержано.
На стадий бластулы наблюдалась остановка развития. Нередко гаструл не образовывалось вовсе, а бластоцели наполнялись комками клеток. Остановка могла продолжаться дня 2. Полученные плютеусы все имели чисто материнский тип. Величина их ядер оказалась неотличимой от диплоидных ядер чистых Echinus и Strongylocentrotus, Поэтому Бальцер присоединился к мнению Годлевского, что элиминации хромосом здесь не происходит и что в этих гибридных плютеусах хромосомы Antedon пребывают в совершенно латентном, инертном состоянии. Нужно все же иметь в виду, что хромосом типа Antedon в гаструлах никто констатировать не пытался.
Совершенно аналогичный случай, упоминавшийся выше, мы встречаем у рыб, по данным Моррис (1911). При скрещивании рыб Fundulus и Ctenolabrus она проследила хромосомы, обоих видов в ряде делений. Они резко различны по форме — палочкообразные у Fundulus и округленные у Ctenolabrus. Эту свою форму они точно сохраняют во всех последовательных делениях, доказывая, что хромосомы обоих видов (родов) одинаково размножаются и с совпадающими темпами распределяются по бластомерам.
Сама М. Моррис проследила развитие гибридов лишь до 12—24-часовой стадии, после, которой они отмирали, как впрочем вскоре (на 24-часовой стадии), отмирали и эмбрионы, чистых видов. Таким образом установить, по какому типу развивался эмбрион, Моррис не могла. Но Лебу (1912) с лучшей аквариумной техникой удалось воспитывать этих гибридов [41] до месяца и более, так что они развивали сердце, кровеносные сосуды, глаза и пр., хотя и не вылуплялись никогда. Эти гибриды по Лебу были вполне матроклинными, так что Леб считал, что в данном случае имеет место лишь возбуждение яиц к развитию чужой спермой в качестве особого случая искусственного партеногенеза. Некоторые уклонения от нормы, получившиеся среди них, не носили ни в каком смысле гибридного характера и легко могли быть вызваны и у чистых Fundulus легкой прибавкой NaCN.
Если увязать исследования Моррис и Леба в одну цепь, то окажется, что 1) сперматозоид Ctenolabrus действительно оплодотворил яйцо Fundulus, 2) хромосомы Ctenolabrus прослежены ясно во всех делениях вплоть до той стадии, до которой исследование было проведено, и 3) гибриды оказываются матроклинными, откуда вытекает вывод, что хромосомы нормально размножаются, но на организм никакого действия не оказывают, становясь «инертными»[42].
Однако такого вывода пока делать конечно нельзя, т. е. нельзя без дальнейшего считать условия опытов Моррис и Леба одинаковыми. Развитие гибридов у Моррис, начиная с 12 часов, обнаруживало характерные аномалии в виде образования гигантских клеток и гибрид погибал (по погибал и контроль!). У Леба же возможно действительно был партеногенез, почему личинки и жили благополучно до месяца. Тем не менее исследовать этот вопрос более тщательно представляло бы конечно огромный интерес для всей теории действия генов вообще и в гибридах в частности.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК