КАСТЫ И РАЗДЕЛЕНИЕ ТРУДА
КАСТЫ И РАЗДЕЛЕНИЕ ТРУДА
Какую удивительную картину представляют нам эти общества насекомых! Ничего подобного не встречаем мы среди высших животных. Бесспорно, у некоторых млекопитающих и птиц замечается стремление к общественности, они соединяются вместе для постройки общего жилища... но принцип разделения труда, специализация отдельных классов на определенных видах работы встречается помимо насекомых только в человеческом обществе, и то на известной ступени цивилизации. Г. У. Бейтс. Натуралист на реке Амазонке
Возникновение эусоциальности знаменуется разделением насекомых в колонии на касты — плодовитую и бесплодную. Первоначально различия между ними затрагивают только функцию размножения. У наиболее примитивных по социальной организации пчел-аллодапин, некоторых галиктов, ос-стеногастрин насекомые обеих каст одинаковы по размерам и строению тела. По мере углубления и совершенствования разделения функций появляются и усиливаются различия в строении тела между насекомыми разных каст.

55. Один из наиболее выраженных случаев кастового диморфизма среди ос-полибий (по Г. Эвансу и М. Вест-Эберхард, 1970 г.).
1 — царица; 2 — рабочий бразильской осы стелополибия флавипеннис.
Плодовитые самки ос-полистов и большинства полибий лишь в среднем крупнее рабочих, и между ними существуют все переходные формы. Среди ос только у некоторых полибий и всех веспин появился полный кастовый диморфизм, когда промежуточных между двумя кастами особей нет. Такая ситуация характерна для высокоорганизованных общественных насекомых.
Среди шмелей мы находим все этапы эволюции кастового диморфизма. Он отсутствует у лесного, малого каменного, мохового, полевого, садового шмелей; царицы этих видов лишь в среднем крупнее рабочих. У городского шмеля промежуточные между кастами особи есть во многих, но не во всех семьях. Полный кастовый диморфизм типичен для каменного, лугового подземного и в наибольшей степени для земляного и обыкновенного шмелей — царица этих видов значительно крупнее рабочих. В общем для шмелей характерна закономерность, отмеченная впервые еще Ф. Слейденом: кастовые различия менее всего выражены у карманщиков, а некарманщиков, как правило, отличает полный диморфизм каст.
У примитивных муравьев и термитов размеры плодовитых и бесплодных особей нередко почти одинаковы, но они хорошо различаются по наличию или отсутствию крыльев, глазков, окраске и другим признакам, т. е. кастовый диморфизм всегда налицо. У продвинутых видов различия между кастами бывают весьма внушительными, что мы уже иллюстрировали.
Возникновение и усиление кастового диморфизма является следствием углубления функциональных различий между кастами. У наиболее примитивных общественных насекомых царица отличается от рабочих в основном тем, что откладывает яйца и препятствует размножению других самок. Царицы галиктов, полистов, шмелей участвуют в кормлении личинок, строительстве, защите гнезда и даже вылетают иногда за кормом. В эволюции происходит постепенная специализация плодовитой касты: функции цариц мало-помалу ограничиваются размножением и выделением феромонов, управляющих поведением рабочих. Такую эволюционную тенденцию можно проследить почти в любой группе общественных насекомых.
У муравьев мирмеций, амблиопон и прочих наиболее примитивных понерин царицы ведут себя в гнезде во многом сходно с рабочими: ухаживают за расплодом, занимаются «туалетом» других особей, обрабатывают принесенную добычу, участвуют в кормовом обмене, причем нередко не только получают, но и отдают пищу, иногда они даже выходят из гнезда в качестве фуражиров. Почти столь же разнообразно может быть поведение цариц у видов тех родов высших муравьев (маника, мирмика, мирмецина, лептоторакс и др.), для которых характерны слабые различия между кастами. У большинства же видов царица совсем не участвует или участвует незначительно в тех делах, которыми заняты рабочие, и уж, конечно, никогда не выходит из гнезда. Но, так сказать, в молодости царицы муравьев обычно умеют и строить, и кормить личинок, и помогать другим особям выбираться из куколок — они делают все это при основании новых гнезд. Затем, когда в гнезде появляется достаточно рабочих, царица «забывает» эти навыки и сосредоточивается на своей основной функции — размножении. Можно сказать, что у таких видов имеет местовозрастная функциональная специализация плодовитой касты. Это же явление характерно для термитов и в меньшей степени для ос и шмелей.
Царицы размножающихся роением медоносных и безжалых пчел необратимо утратили разнообразие поведения и «многогранность» своих предков: они никогда не строят, не ухаживают за расплодом, не кормят и не облизывают других насекомых; функции их ограничены откладкой яиц, питанием и воздействием на рабочих через выделяемые феромоны; один или несколько раз в жизни они участвуют в роении и совершают брачные полеты. Перед нами яркий пример доведенной почти до предела специализации поведения плодовитой касты.
Итак, на основе первоначального разделения функций между двумя кастами в эволюции появляются и углубляются поведенческие и морфологические различия между ними, насекомые разных каст становятся все более непохожими друг на друга. Это приводит к возникновению так называемого социального полиморфизма, т. е. наличию в колонии морфологически четко различающихся особей. У множества общественных насекомых он сводится к кастовому диморфизму. Но нередко полиморфизм приобретает более сложный характер, поскольку морфологически разные формы появляются в пределах одной касты.
Только у немногих видов этот процесс затрагивает плодовитую касту. В колониях термитов иногда могут одновременно встречаться адультоидные, нимфоидные и эргатоидные производители; функции их одинаковы, различно лишь происхождение и строение тела. В гнездах муравьев рода мирмика наряду с нормальными царицами изредка попадаются так называемые микрогины — царицы обычного строения, но примерно такого же размера, как рабочие. В гнезде морщинистой мирмики могут быть либо нормальные царицы, либо микрогины. Эти миниатюрные самки не способны в одиночку основывать новые гнезда, и поэтому колонии с микрогинами размножаются отделением отводков. У рыжей мирмики микрогины могут жить только вместе с нормальными царицами. В последнее время появились доказательства того, что микрогины — результат наследственной аномалии. Довольно загадочны также причины появления эргатоидных, т. е. имеющих облик рабочих, цариц у некоторых видов муравьев. Значительно чаще полиморфизм развивается внутри бесплодной касты, что мы рассмотрим немного ниже.
Разделение труда, или функций, у общественных насекомых получило название «полиэтизм». Различие в поведении цариц и рабочих — это кастовый полиэтизм. Но формы разделения труда между насекомыми бесплодной касты гораздо разнообразнее. Наблюдая за их поведением, мы можем выделить так называемыеполиэтические, или функциональные, группы, хорошо различающиеся по «обязанностям» в гнезде и вне его. Например, среди рабочих муравья-понерины эктатомма руидум исследователи выделили пять функциональных групп: муравьи-няньки, посвящающие значительное время уходу за личинками; няньки, ухаживающие в основном за куколками; малоактивные муравьи, являющиеся резервом рабочей силы; рабочие-сторожа, специализирующиеся на охране гнезда; и рабочие-фуражиры. Поведение каждого муравья (а все они имели индивидуальные метки) было стабильным и четко укладывалось в рамки этих полиэтических групп.
Еще один пример. В сооружении гнезда обитающих в тропических лесах Малайзии муравьев-ткачей (кампонотусы гомбаки и тексенс) участвуют три группы строителей. Рабочие первой группы приносят личинок и, удерживая челюстями поперек туловища, постукивают их усиками, а личинки, быстро двигая головой и передней частью тела, выплетают из шелковых нитей стенку павильона — так называют это гнездо. Муравьи второй группы притаскивают песчинки, мелкие кусочки листьев, древесины и плотно прижимают их к стенке; эти частицы тоже вплетаются в гнездо, что повышает его прочность. На каждого муравья с прядущей личинкой приходится в среднем два приносящих строительный материал. Почти в пять раз больше муравьев составляет третью группу. Они челюстями и передними ногами тщательно утрамбовывают стенку и вплетаемые в нее крупицы. В результате шелковая поверхность становится очень гладкой и крепкой.
Разделение труда у общественных насекомых может проявляться в двух формах. Первая — это расчленение задач, когда весь «трудовой процесс» разделен на ряд операций, которые выполняют последовательно разные насекомые, но рабочие при этом равноценны и в то или иное время могут быть заняты любой операцией. Например, весь процесс сооружения ячейки и снабжения ее провизией разделен у пчел-галиктов на ряд этапов. При этом одни пчелы строят ячейку, другие приносят пыльцу и нектар, третьи оформляют пыльцевой хлебец. Однако каждая пчела регулярно меняет род своей деятельности: она то вылетает за кормом, то становится «землекопом», то «булочником», изготавливающим хлебцы. Получается, что разделение труда существует, но любая пчела остается мастером на все руки. Такая форма организации работ у общественных насекомых, очень распространена.
Если проследить за поведением отдельных медоносных пчел в улье, то можно убедиться, что в течение дня каждая из них чистит ячейки, кормит личинок, строит, запечатывает ячейки крышечками, принимает нектар и перерабатывает его в мед, утрамбовывает запасы пыльцы в ячейках и т. д. Все эти занятия сменяют друг друга без особого порядка круглые сутки. У многих видов ос-веспин рабочие еще более «многогранны». В продолжение одного дня они могут заниматься совершенно разными делами — от кормления личинок и царицы до фуражировки и строительства.
Вторая форма разделения труда — функциональная специализация, когда группы насекомых выполняют лишь вполне определенный набор операций. Функциональная специализация, конечно, невозможна без расчленения задач, но, как мы только что видели, последнее само по себе не предполагает обязательной функциональной специализации насекомых. Выполнение рабочим определенных функций может иметь различную продолжительность — от нескольких дней до многих недель и месяцев — и даже бывает пожизненным. Как же происходит подобная специализация? Известен ряд факторов, могущих служить основой разделения функций. В соответствии с ними выделяют три типа полиэтизма рабочих особей.
Морфологический, или размерный, полиэтизм может существовать, когда рабочие различаются по строению или размерам тела. У общественных пчел и ос такая изменчивость обычно невелика. Поэтому и размерный полиэтизм играет у них очень скромную роль. У ос-веспин рабочие покрупнее в среднем чаще заняты фуражировкой, а осы помельче больше времени проводят в гнезде. Сходна картина у шмелей. Но у многих их видов рабочие изменчивее по размерам, чем у ос, поэтому и размерный полиэтизм выражен у них отчетливее. Мелкие рабочие шмели обычно гораздо реже покидают гнездо, чем их более крупные собратья. А с увеличением числа рабочих в гнезде шмели-крошки полностью посвящают себя внутригнездовым заботам.
Рабочие многих видов муравьев обладают незначительной размерной изменчивостью, в таких случаях морфологический полиэтизм практически не выражен. Но все же гораздо больше муравьев, у которых рабочие сильно варьируют по размерам тела. И тем не менее было бы ошибкой думать, что у всех таких видов существует морфологический полиэтизм. Например, у бегунков рабочие могут различаться по размерам почти в два раза, но выполняют фактически одинаковые функции. Однако такая ситуация достаточно редко встречается среди муравьев, и у большинства видов размерный полиэтизм есть. Но характер его может быть очень неодинаковым в зависимости от вида.
У рыжих лесных муравьев внутригнездовые муравьи (няньки и др.) в среднем немного крупнее работающих вне гнезда. Это происходит потому, что рабочие покрупнее имеют тенденцию начинать деятельность вне гнезда в более старшем возрасте, чем муравьи помельче. Значит, размер муравья влияет на возрастную смену функций, о которой мы еще будем говорить.
Среди муравьев, работающих вне гнезда, в среднем самые мелкие являются сборщиками пади тлей, более крупные — это охотники, и самые крупные приносят в гнездо строительный материал. Подчеркнем, что эти три полиэтические группы отличаются только по средним размерам, включая и маленьких, и крупных рабочих, но в разном количестве. Так что по величине тела конкретного муравья невозможно узнать его функцию. Размер муравья определяет лишь вероятность выполнения им той или иной роли, и функциональная специализация находится под контролем еще и других факторов.
Довольно велика изменчивость размеров рабочих у кампонотусов. У европейского муравья-древоточца самые мелкие в среднем особи — это няньки; рабочие промежуточных размеров — охотники, доставляющие добычу в гнездо, а самые крупные муравьи преобладают среди защитников гнезда при возникновении опасности. У других видов этого рода картина в общем сходная, но самые крупные рабочие не всегда являются активными защитниками гнезда: у некоторых видов (кампонотусы пенсильванский, модок) они в основном заняты доставкой в гнездо пади, собираемой мелкими муравьями.
В гнездах муравьев, сохраняющих запасы сладкой пищи в зобиках, эту функцию выполняют всегда наиболее крупные рабочие. Так бывает и у видов с еще незначительно специализированными «хранителями меда» (проформики, некоторые формики и катаглифисы), и у настоящих медовых муравьев. Когда из гнезда медового муравья удалили все «медовые бочки», то вскоре начали появляться новые. При этом оказалось, что в них превращались наиболее крупные из оставшихся в гнезде муравьев.
Особенно развит морфологический полиэтизм у тех муравьев, рабочие которых не только достаточно изменчивы по размерам, но и значительно различаются по строению тела в зависимости от величины. Так бывает в случае неравномерного роста, когда при увеличении размеров тела его пропорции меняются. По существу, рост большинства организмов, в том числе и человека (вспомните, насколько различны пропорции тела у новорожденного малыша и взрослого человека), не равномерен. Таков же рост подавляющего числа насекомых, но только он происходит на стадии личинки, и вышедшее из куколки насекомое обладает уже окончательными пропорциями и строением тела. У многих муравьев рост гораздо более неравномерный, чем у основной массы необщественных насекомых. Это приводит к тому, что муравьи разных размеров настолько существенно отличаются друг от друга, что их стоило бы отнести к разным видам, если бы не было известно, что они живут в одном гнезде. В таких случаях в пределах касты рабочих выделяют подкасты.
Чрезвычайно изменчивы по размерам, рабочие листорезов атта. Ширина головы самых мелких из них всего около 0,65мм, а наиболее крупных — почти 4 мм. При этом чем крупнее муравей, тем мощнее по сравнению с размерами тела его голова, массивнее челюсти, длиннее их зубчики. Самые крупные рабочие имеют громадную сердцевидную голову, основную часть объема которой занимают мощные мышцы, приводящие в движение внушительные челюсти. Такое непропорциональное увеличение головы у рабочих-исполинов как раз и вызвано необходимостью максимально усилить их челюсти. Однако частотное распределение рабочих атта по размерам непрерывное; это означает, что мы можем найти муравьев любых размеров — от самых мелких до наиболее крупных, а пропорции и строение их тела меняются при увеличении размеров постепенно. Больше всего в гнездах рабочих средних размеров, с шириной головы от 1 до 2 мм. Такое частотное распределение называется одновершинным.
Наблюдения за поведением листорезов позволили выделить среди рабочих четыре функциональные группы. Самые мелкие муравьи являются няньками или ухаживают за грибными садами, рабочие покрупнее занимаются разнообразными делами внутри гнезда, еще более крупные становятся землекопами или фуражирами, срезающими листья, ну, а самые крупноголовые муравьи — это солдаты, защитники гнезда и фуражировочных дорог. Между подкастами не существует резких переходов, и рабочие промежуточных размеров могут относиться либо к той, либо к другой полиэтической группе. Размер тела влияет на выбор муравьями рода деятельности и внутри подкаст. Например, средние размеры муравьев, срезающих и приносящих в гнездо листья, больше, чем у тех, что разрезают их на куски в гнезде, а последние в среднем крупнее особей, занятых вторичной обработкой — пережевыванием частичек листьев; немного мельче муравьи, укладывающие эти частички в грибные сады, еще мельче те, что высевают грибные гифы, и самые маленькие в среднем рабочие ухаживают за прорастающей грибницей.
У всеядного огненного муравья, по наблюдениям Э. Вильсона, самые мелкие рабочие в основном являются няньками, а рабочие среднего размера и особенно самые крупные — строителями и фуражирами. Мелкие рабочие могут выполнять практически любые функции, и их поведенческий репертуар, как выражаются специалисты, шире, чем у средних и крупных муравьев, которые более специализированы. У другого вида этого же рода — соленопсис гемината, специализированного на питании преимущественно семенами, полиморфизм рабочих выражен значительно ярче: крупные муравьи обладают большой головой с мощными лишенными зубчиков челюстями, а промежуточных между крупными и средними рабочих мало: частотное распределение их по размерам становится двувершинным, оставаясь при этом непрерывным. Поведенческий репертуар крупноголовых рабочих очень беден; они заняты исключительно пережевыванием семян, доставляемых в гнездо другими муравьями. Рабочие среднего размера тоже могут пережевывать семена, а мелкие муравьи никогда этого не делают. Таким образом, мы видим, что по мере усиления полиморфизма рабочих углубляется функциональная специализация подкаст. У муравьев-портных мы наблюдаем уже практически полный диморфизм рабочих: промежуточных по величине тела муравьев в семье очень немного. Функции двух подкаст у этих муравьев почти не перекрываются. Крупные рабочие добывают львиную долю корма и выполняют самую трудоемкую операцию при строительстве гнезда — притягивают друг к другу листья; они никогда не ухаживают за личинками и царицей. Мелкие рабочие — это прежде всего няньки, лишь единицы выходят из гнезда за пищей; при строительстве они приносят личинок и сшивают с их помощью листья, стянутые крупными муравьями.

56. Полиморфизм муравья феидоле инстабилис, имеющего рабочих всех размеров (непрерывное частотное распределение) (по В. Вилеру, 1910 г.)
1 — крупный рабочий (солдат); 2, 3 — средние рабочие; 4 — мелкий рабочий; 5 — царица; 6 — самец.
Гораздо реже, чем виды с вариабельными по размерам и полиморфными рабочими при непрерывном одно- или двух- и даже многовершинном частотном их распределении, встречаются среди муравьев виды с прерывистым распределением, т. е. с полным диморфизмом рабочих. Они есть не более чем в 15 родах муравьев. Наиболее известный пример диморфизма — распространенный по всей Земле, тропический по происхождению род феидоле. У всех феидоле солдаты с массивной головой и мощными челюстями, промежуточные между рабочими и солдатами по размерам и строению тела муравьи сохраняются лишь у некоторых видов, а для прочих характерен полный диморфизм. Солдаты обитающего в Европе и Азии феидоле паллидула и многих других видов выходят из гнезда за кормом вместе с рабочими. Они расчленяют и транспортируют крупную добычу, перемалывают собранные зерна, а также являются основными защитниками гнезда. У видов, которые зернами не питаются, например у обитающего на юге Приморского края феидоле фервида, солдаты помимо защиты гнезда выполняют еще одну функцию — сохраняют в зобиках жидкий корм. Подобная же специализация характерна для солдат родов олигомирмекс и эребомирма, относящихся, как и феидоле, к мирмицинам.
Чрезвычайно интересны солдаты пробкоголового муравья. Передняя часть их головы почти совершенно плоская и имеет очень твердую кутикулу. Муравьи эти поселяются в древесине, и основная задача их солдат — закрывать головами выходы из гнезда. Голова солдата играет роль живой пробки: сам он сидит внутри, а плоская часть головы вместе с челюстями находится вровень с краями отверстия, как щит. Попробуй сунься — не пройдешь! Но когда к входу подбегает рабочий муравей (у него в отличие от солдата нормальная форма головы) из этого же муравейника, он прикасается усиками к голове солдата, и тот в ответ на «пароль» отступает в глубь гнезда, пропуская своего собрата. Подобные же солдаты-блокировщики входов есть и у тропических мирмицин родов паракриптоцерус и закриптоцерус. Солдаты-пробки ничем другим не занимаются и никогда не покидают гнезда. Это пример чрезвычайно узкой специализации поведения. Кроме упомянутых выше полный диморфизм рабочих известен в родах мономориум, соленопсис, феидологетон, проформика, псевдолазиус.
Очень разнообразны солдаты термитов. У всех видов они выполняют единственную функцию — защищают гнездо, поэтому их поведенческий репертуар крайне беден. При возникновении опасности солдаты устремляются к поврежденным участкам гнезда и занимают круговую оборону или же просто высовывают головы из разломов и отверстий наружу. Солдат приподнимается на передних ножках и поводит из стороны в сторону головой с раскрытыми челюстями, иногда он покачивается на ножках вперед-назад. Как только солдата касается тело врага, он

57. Защита гнезда солдатами европейского пробкоголового муравья (по Дж. Жабо-Патай, 1928 г.).
1 — рабочий муравей прикасается усиками к голове солдата, закрывающего изнутри вход в гнездо, чтобы тот пропустил его; 2 — голова солдата с уплощенной передней частью; 3 — вид снаружи входа, «запечатанного» головой солдата; 4 — несколько солдат, закрывающих головами широкий вход в гнездо.
впивается в него мертвой хваткой. На что-либо более «интеллектуальное» эти ограниченные создания не способны. Фактически солдаты в колонии термитов действуют так же, как стрекательные клетки гидры или медузы, — срабатывают в момент прикосновения к ним тела врага, вот и все! Но даже в своем единственном занятии — защите гнезда — солдаты эффективны только при встрече с достаточно крупными врагами, в первую очередь позвоночными животными. Мелкие же муравьи, например, легко проникают в разрушенное гнездо, минуя солдат, которые их не замечают. В борьбу с такими врагами вступают рабочие термиты.
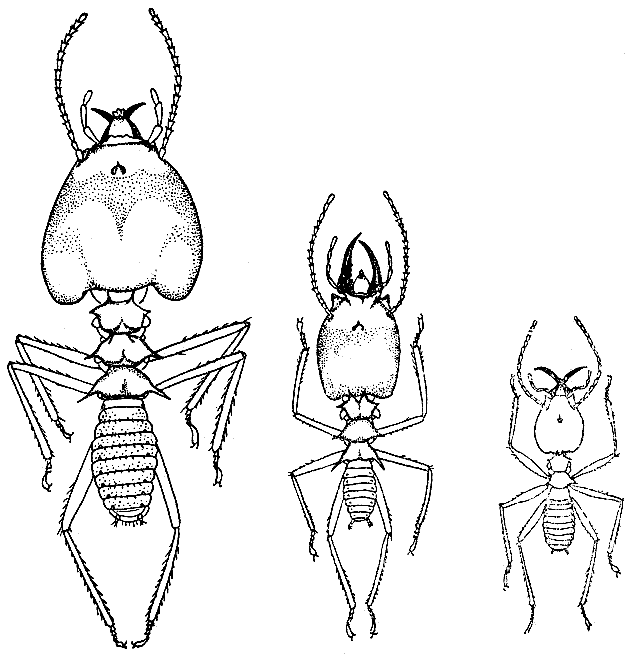
58. Полиморфизм солдат у гриборазводящего термита акантотермес акантоторакс (по П. Грассе, 1949 г.).
Между тремя размерными группами солдат переходных форм
Солдаты большинства термитов имеют челюсти с режущими зубчиками по краю. Еще эффективнее в борьбе с врагами-позвоночными солдаты с колющими серповидными челюстями, лишенными зубцов (роды акантотермес, макротермес, синтермес, термес). Эти челюсти вонзаются в тело врага так глубоко, что концы их смыкаются и укус очень болезнен. Кроме того, избавиться от солдата можно, только разорвав его на части и вынув обе челюсти по очереди! Недаром местное население в тропиках издавна использовало солдат с серповидными челюстями для сшивания ран. К сближенным краям раны подносят солдата, и он впивается в них челюстями. Затем брюшко солдата отрывают, а голова его остается на ране, как хирургическая скрепка. Такие же серповидные челюсти у солдат американских муравьев-кочевников (лабидус, эцитон) и многих муравьев-рабовладельцев.
 59. Голова термита перикапритермес аппелланс с щелкающими челюстями (по П. Грассе, 1949 г.)
59. Голова термита перикапритермес аппелланс с щелкающими челюстями (по П. Грассе, 1949 г.)
Удивительны солдаты со щелкающими челюстями. Мандибулы их асимметричны и налегают друг на друга. Солдат может щелкать ими наподобие того, как мы щелкаем большим и средним пальцами руки. При этом одна из челюстей с силой опускается вниз, и если солдат находится на теле врага, тот получает очень чувствительный удар. Такие солдаты легко проламывают хитиновый панцирь крупных насекомых и причиняют боль позвоночным животным. А кроме того, ударяя челюстями по земле, солдаты отпрыгивают на несколько сантиметров назад.
Среди суходревесных термитов у многих видов рода криптотермес есть солдаты-блокировщики. Своей крупной, почти цилиндрической головой с уплощенной передней частью и очень маленькими челюстями они, как пробкой, закрывают ходы и галереи, не допуская посторонних в поврежденное гнездо. Удивительно сходный с некоторыми муравьями, но самостоятельно возникший в эволюции способ защиты! Наконец солдаты целого ряда термитов борются с врагами настоящим химическим оружием. О том, как это происходит, мы расскажем в пятом разделе.
Рабочие и солдаты множества видов термитов могут быть разного размера, что связано с неодинаковым числом пройденных ими возрастов или с полом. В таких случаях функции термитов разного размера обычно существенно различаются. У термитов-грибоводов фуражируют преимущественно крупные рабочие (самцы), а мелкие рабочие (самки) трудятся внутри гнезда. У носачей рода назутитермес повреждения гнезда ремонтируют только крупные рабочие (самки), а в фуражировке участвуют обе категории рабочих. Многочисленные в гнезде носачей мелкие солдаты (самцы) являются основными защитниками, а присутствующие в значительно меньшем количестве крупные солдаты (самки) играют роль «сигнальщиков». При возникновении опасности они устремляются в глубь гнезда и мобилизуют мелких солдат на оборону, но сами в сражении с врагами не участвуют.
Вторым способом разделения труда является возрастной полиэтизм — смена функций во времени, связанная с возрастом особи. У наиболее примитивных общественных насекомых такой полиэтизм обыкновенно развит слабо или совсем не выражен. Молодые пчелы-галикты первые дни своей жизни нередко посвящают охране входа в гнездо, а затем занимаются строительством и фуражировкой. У ос-полистов и шмелей возрастной полиэтизм практически отсутствует. Среди ос-веспин он выражен в различной степени в зависимости от вида. Рабочие обыкновенной и германской ос, восточного шершня и других видов приступают к трудовой деятельности со строительства, через несколько дней к нему добавляется кормление личинок, а затем осы начинают вылетать из гнезда за кормом и строительным материалом. Таким образом, с возрастом у осы поведенческий репертуар расширяется.
Все высокоорганизованные общественные насекомые обладают хорошо выраженным возрастным полиэтизмом. Впервые он был описан у медоносной пчелы Ч. Батлером еще в 1609г.: «Молодые пчелы, как наиболее сильные и здоровые, несут наибольшее бремя, поскольку они не только работают вне улья, но и денно и нощно приглядывают за всем в доме и охраняют его. При необходимости они рискуют своими жизнями при защите гнезда, они изгоняют трутней и сражаются с другими пчелами и осами, набрасываются со своими жалами на всякого, кто потревожит их, они выносят своих мертвецов наружу, чтобы похоронить их, и выполняют все другие обязанности. Но работа старых пчел — это лишь сбор пищи, который они никогда не прекращают до тех пор, пока крылья еще могут носить их, и когда они перестают работать, они также перестают и есть, вот какими врагами праздности они являются»[9]. В дальнейшем многие ученые наблюдали возрастные изменения в поведении пчел, но детально это явление было исследовано в середине нашего века немецким энтомологом М. Линдауэром.
Первые 1—2 дня своей жизни молодая пчела бездеятельна. У нее начинают быстро развиваться кормовые и восковые железы. Отправным пунктом ее трудовой карьеры является чистка ячеек, а с 3—4-го дня жизни она активно включается во все события, происходящие в улье, и выполняет самые различные функции без особого порядка. Кормит личинок пчела примерно до 15-го дня своей жизни, в это время у нее как раз интенсивно работают кормовые железы. Выделять воск пчела начинает с 4— 5-го дня, после 15-го восковые железы постепенно дегенерируют и пчела перестает строить. Вылетают из улья пчелы уже на 4— 5-й день, но пока совершают лишь ориентировочные облеты, знакомятся со своим домом снаружи. Позже они выносят из гнезда трупы пчел и другой мусор. Такие вылеты составляют незначительные эпизоды в жизни ульевой пчелы, загруженной прежде всего делами внутри гнезда. Но вот в возрасте 15—20 дней пчела вдруг бросает почти все свои прежние занятия и становится сборщицей. Отныне и до конца своей недолгой жизни она будет приносить корм для семьи. Лишь немногие пчелы этого возраста продолжают вентилировать улей или стоять на страже возле летка.
Возрастная смена функций у безжалых пчел осуществляется примерно так же. Но продолжительность жизни их рабочих почти в два раза больше, чем у медоносной пчелы. Поэтому и переход пчелы в сборщицы приходится на более поздний возраст. У некоторых видов безжалых пчел (род гипотригона) окраска брюшка с возрастом темнеет, что позволяет хорошо различать приблизительно пять возрастных групп.
Изменение поведения муравьев с возрастом впервые наблюдал еще О. Форель в конце прошлого столетия. В середине нашего века возрастной полиэтизм был подробно изучен на примере рыжих лесных муравьев немецким исследователем Д. Отто и видов рода мирмика англичанином Дж. Уэйром, а затем еще и на других муравьях. Оказалось, что у примитивных амблиопон этот способ разделения труда отсутствует: молодые муравьи сразу же начинают заниматься всеми делами, в том числе и добывать корм. Но у других понерин и мирмеций возрастной полиэтизм уже существует.
Общая схема возрастной смены функций у всех видов муравьев примерно одинакова и сходна с таковой у пчел. Значительную часть жизни муравьи проводят в гнезде. Вначале они являются няньками — ухаживают за расплодом и царицей. Именно в это время у них хорошо функционируют кормовые железы и яичники, в которых развиваются кормовые яйца. Рабочие постарше — так называемые гнездовые муравьи — занимаются различными делами внутри муравейника, кроме тех, что выполняют няньки, или же пассивно ожидают «приглашения» на какую-нибудь работу от других рабочих. Наконец самые старшие муравьи переходят к деятельности вне гнезда и становятся охотниками, строителями, сборщиками пади. Это совершается у рыжих лесных муравьев в возрасте 1,5—2 месяца, но срок может сильно сдвигаться в ту или иную сторону в зависимости от ряда обстоятельств, которые мы рассмотрим чуть позже. Остаток своей жизни муравьи-«ветераны» проводят на куполе гнезда в роли так называемых «наблюдателей», первыми замечающих приближение опасности. Впервые наблюдателей обнаружил советский энтомолог П. И. Мариковский.
С возрастом у муравьев происходит ряд изменений: хитиновые покровы тела постепенно все более темнеют, зубчики на челюстях, стираясь, становятся тупыми; меняются и некоторые черты «характера». Молодые муравьи-няньки очень пугливы и совсем не агрессивны. Когда возникает опасность, они начинают уносить куколок и личинок и сами скрываются. Гнездовые рабочие не такие робкие и не прячутся, они уносят расплод, но редко проявляют заметную враждебность. Самые старшие муравьи-фуражиры очень агрессивны и первыми бросаются на врага.
Возрастной полиэтизм развит и у термитов. Он довольно хорошо исследован москвичом К. С. Шатовым на примере большого закаспийского термита. Рабочие этого вида имеют восемь возрастов, различающихся по размерам тела. Самые молодые (первый — третий возрасты) являются няньками, ухаживают за яйцами и личинками. В свиту царской пары входят рабочие второго-третьего возрастов. Каста грумов, облизывающих других термитов, состоит из особей второго — четвертого возрастов. Более старшие термиты становятся фуражирами или водоносами. Водоносы — это рабочие в основном четвертых — шестых возрастов; они опускаются за водой в глубокие вертикальные ходы и отдают ее термитам в гнезде и фуражирам, которым она нужна для строительства фуражировочных тоннелей и лепки. Самые старшие рабочие (седьмой-восьмой возрасты) трудятся в роли жнецов; именно они срезают сухую траву, веточки кустарников и разгрызают их на кусочки. А более молодые фуражиры — рабочие четвертого — шестого возрастов — служат носильщиками, доставляя срезанные жнецами кусочки в гнездо. Возрастной полиэтизм высших термитов пока недостаточно исследован. Но то, что мы уже знаем, вполне соответствует картине, нарисованной для большого закаспийского термита.
Третий тип полиэтизма пока наименее изучен. Его можно назвать полиэтизмом предпочтений. Во многих случаях мы встречаемся с функциональной специализацией среди одновозрастных и одинаковых по размерам и строению тела насекомых. Когда экспериментаторы создали искусственную семью черного садового муравья, состоявшую из царицы и нескольких десятков вышедших в одно время из куколок рабочих, то оказалось, что в ней существует отчетливое разделение функций. Одни муравьи были няньками, другие приносили корм, третьи занимались строительством — и все они были практически одинаковыми по величине. Можно предполагать, что каждый муравей выбрал «профессию» в значительной степени соответственно своим склонностям. Когда муравьев заставляли переносить куколок из освещенной в затемненную часть гнезда, то некоторые проявляли кипучую деятельность и перетаскивали до 90% всех куколок; несколько муравьев таскали куколок гораздо менее охотно, еще несколько — делали это изредка, а остальные вообще не участвовали в работе. Сходную картину обнаружили, когда муравьям нужно было рыть песок. Но при выполнении этой задачи самыми трудолюбивыми проявили себя совсем не те муравьи, что старательно таскали куколок. В третьем эксперименте, когда муравьям была предложена живая добыча, лидерами стали опять другие рабочие.
Какие внутренние свойства насекомого обусловливают предпочтение им той или иной работы? Пока мы этого не знаем. Но постепенно в науке накапливается все больше фактов о том, что насекомые даже одной семьи не являются одинаковыми по психофизиологическим свойствам, что каждое из них, как и мы с вами, обладает собственным «характером» и даже имеет определенные «вкусы». Одни агрессивны и смелы, склонны обследовать незнакомые предметы, другие, напротив, робки и миролюбивы, в непривычной обстановке пугаются и прячутся. Мы знаем уже, что эти качества изменяются с возрастом. Но даже у одновозрастных насекомых они могут различаться. Исходя из представлений современной генетики и этологии (науки о поведении животных), можно полагать, что «характер» насекомого в значительной мере определяется его наследственностью, является врожденным, но на него обязательно влияют и внешние обстоятельства. В какой степени? На этот вопрос наука пока не. имеет ответа. Мы еще вернемся к нему в конце книги.
В специальных опытах выяснилось, что охотники рыжих лесных муравьев агрессивны, инициативны, проявляют внимание к незнакомым предметам. А сборщики пади, напротив, очень пугливы, не агрессивны, не склонны к инициативе. Может быть, именно эти свойства «характера» и помогли им выбрать «профессию»? Вероятно, что это так. Но пока мы не знаем, в какой мере может меняться «характер» под влиянием избранной «профессии». Не исключено, что постоянно выполняемая работа модифицирует некоторые психофизиологические качества насекомого.
У большинства достаточно высокоорганизованных общественных насекомых среди фуражиров хорошо заметны так называемые активные фуражиры, или разведчики. Это самые деятельные и инициативные рабочие, проявляющие устойчивый интерес ко всему новому и необычному. Они постоянно заняты обследованием местности, поиском источников пищи. Обнаружив корм, разведчики мобилизуют на его использование пассивных фуражиров, которые сами никогда предприимчивыми не бывают, провизию не ищут и при отсутствии работы праздно сидят в гнезде. Активных фуражиров в гнезде всегда значительно меньше, чем пассивных. Такая организация фуражировки, как правило, наиболее выгодна. Она позволяет обходиться при поисках корма малым числом опытных разведчиков, но когда пища найдена, максимально быстро привлекать для ее использования необходимое количество пассивных фуражиров. Какие насекомые становятся разведчиками, а какие предпочитают быть пассивными фуражирами? Скорее всего, это в значительной мере определяется их «характером».
Мы видим, что система разделения труда у высокоорганизованных общественных насекомых чрезвычайно сложна. Выбор рабочим конкретной функции находится под влиянием множества факторов. С одной стороны, это величина и особенности строения его тела — морфологические и размерные различия рабочих у многих видов являются основой полиэтизма. С другой стороны, изменения поведения насекомых в течение их жизни могут нередко играть еще большую роль, и возрастные функциональные группы существуют почти у всех видов. Оба эти фактора тесно взаимодействуют. Даже у медоносной пчелы, рабочие которой очень мало варьируют по размерам, пчелы помельче становятся сборщицами в среднем на неделю позже, чем крупные. Гораздо сильнее влияют размеры тела на характер возрастной смены функций у многих муравьев. Крупные и средние рабочие муравьев-древоточцев очень недолго бывают в «няньках», вскоре переходя к деятельности фуражиров. Мелкие же муравьи, напротив, значительно дольше выполняют обязанности нянек, часто оставаясь в этой «должности» навсегда.
Возрастная смена функций находится под влиянием и других индивидуальных свойств насекомых. Так, совершенно одинаковые по величине рабочие муравья тапинома эрратикум существенно различаются по характеру возрастного полизтизма. Одни очень быстро «делают карьеру», проходя путь от няньки до фуражира, причем некоторым даже удается миновать какой-то этап, например вообще не быть нянькой. Другие же муравьи «продвигаются по службе» медленно, надолго, даже на всю жизнь задерживаются в промежуточных «должностях». Вероятно, и «карьеристы», и «неудачники» заведомо обладают чертами «характера», в значительной степени определяющими их жизненный путь. Но пока мы об этом можем только догадываться.
Сильнейшее воздействие на разделение функций оказывают и социальные факторы, прежде всего потребности колонии в том или ином виде труда. Если не хватает корма, фуражиры становятся более деятельными и ряды их пополняют новые рабочие. Появилась потребность расширить гнездо или отремонтировать разрушенную его часть — и множество новичков «приобретает профессии» строителей. Недостаточная численность любой функциональной группы обязательно пополняется «новобранцами», специализирующимися на выполнении данной функции. Как правило, в гнезде всегда в избытке резервные контингента рабочих, ожидающих применения своим силам. У огненного муравья, в частности, они составляют около трети рабочих. Именно эти резервисты становятся, когда нужно, фуражирами, строителями, няньками.
При крайней необходимости возможен даже переход насекомых из одной полиэтической группы в другую. Если из улья удалить всех фуражиров, то многие пчелы, еще не достигшие двухнедельного возраста, преждевременно станут сборщицами и снабжение семьи кормом наладится. Ну, а если изъять всех молодых пчел? Оказывается, даже в таком, вряд ли возможном в природе случае семья справляется с задачей: часть пчел-сборщиц бросает свое занятие и принимается кормить личинок, строить соты и делать мед. И что самое удивительное: у этих пчел-старушек вновь развиваются восковые и кормовые железы, они как бы физиологически омолаживаются, чтобы успешно выполнять новые функции. Аналогичные эксперименты с таким же результатом неоднократно проделывали и на муравьях. Значит, социальные потребности могут не только ускорить или замедлить возрастную смену функций у насекомых, но и повернуть ее вспять!
Оказалось даже, что при размерном полиэтизме возможна компенсация функций. Когда из экспериментального гнезда листорезов атта цефалотес Э. Вильсон изъял почти всех рабочих среднего размерного класса (ширина головы от 1,8 до 2,2 мм), которые в обычных условиях срезают львиную долю доставляемых в гнездо листьев, производительность труда колонии муравьев ничуть не упала. Теперь листья стали резать более крупные и более мелкие муравьи, раньше этого не делавшие. Кроме того, оставшиеся немногочисленные рабочие изъятого размерного класса начали работать примерно в пять раз интенсивнее, как бы стараясь восполнить отсутствие своих «товарищей» по подкасте.
Многие специалисты полагали, что при полном диморфизме бесплодной касты солдаты, обладающие столь бедным поведенческим репертуаром, просто не способны делать что-либо еще. Но не так давно экспериментами Вильсона было доказано: если из гнезда феидоле убрать почти всех рабочих, то солдаты вдруг «вспоминают» навыки, необходимые для выкармливания личинок и заботы о царице, и справляются с этим не хуже рабочих. Вот вам и ограниченные солдафоны! Все умеют, оказывается, но в нормальных условиях заняты только своим делом и не вмешиваются в чужие. Такие же способности были обнаружены и у солдат эребомирмы.
Как же осуществляется точная регуляция распределения рабочей силы и разделения труда? Как получается, что в гнезде общественных насекомых все необходимые работы выполняются своевременно и не бывает авралов или нехватки рабочих рук, кроме вызванных внешними факторами? Такой эффективностью организации труда далеко не всегда могут похвастать и люди с их высоким интеллектом и научно-техническим прогрессом. Современная наука может ответить на эти вопросы лишь приблизительно. Очень уж это сложная область!
Отдельные насекомые способны иногда на себе ощущать потребности всей колонии. Например, недостаток пищи каждый фуражир может благодаря кормовому обмену почувствовать по собственному желудку и приложить больше усилий к поиску новых источников провизии. Он же может в этом случае мобилизовать на добывание корма фуражиров из числа резервистов. Значит, способность к передаче информации и проведению мобилизации является важным фактором, позволяющим регулировать разделение труда. Мы еще поговорим специально о механизмах мобилизации.
Но не меньшее, а пожалуй, и гораздо большее значение имеет другой регулирующий механизм, основанный на индивидуальных взаимоотношениях насекомых в группах. Вспомним систему иерархии у рыжих лесных муравьев, описанную немного раньше. Самые активные высокоранговые фуражиры препятствуют выходу других муравьев на «арену». Но если этих, занимающих вершину иерархии рабочих убрать, то их тут же заменят муравьи более младших рангов. Вот элементарный механизм, регулирующий разделение труда. Он, вероятно, достаточно универсален.
В гнезде полистов обычно бывает несколько очень деятельных фуражиров, а другие осы вылетают за кормом редко и больше времени проводят бесцельно. Но если хотя бы один деятельный фуражир исчезнет, его тут же заменяет такая «резервная оса», активность которой быстро возрастает. Складывается впечатление, что интенсивно работающие фуражиры тем самым как бы подавляют функционирование малоактивных ос. Но деталей этого механизма мы пока не знаем.
Вернемся к экспериментальной семье черного садового муравья. Если убрать из нее наиболее активных носильщиков, то те, кто раньше работал с прохладцей, начинают трудиться на совесть, заменяя прежних лидеров. Уберем и их — появятся новые. И так до тех пор, пока в гнезде остаются муравьи. Вероятно, и здесь самые активные носильщики своей деятельностью подавляют стремление других участвовать в той же работе. В экспериментах с листорезами атта цефалотес было замечено, что рабочие среднего размерного класса буквально физически оттесняют от листьев муравьев других размеров, которые тоже выходят на «арену», но не допускаются до разрезания листьев. Когда рабочих среднего размера изъяли, это открыло «оперативный простор» для деятельности мелких и крупных муравьев.
Таким образом, разделение труда регулируется в значительной степени на основе иерархии поведенческих ролей, складывающейся между рабочими в группе. По отношению к каждой функции существует своя иерархия, что и позволяет эффективно распределять усилия всех насекомых. Конечно, здесь еще гораздо больше неясного, чем в других механизмах регуляции поведения, и это лишь убедительная гипотеза. К данной теме мы еще вернемся.
Как мы убедились, у примитивных эусоциальных насекомых разделение труда находится в зачаточном состоянии и обычно основано на иерархии доминирования-подчинения. В процессе эволюции с увеличением количества насекомых в колонии осуществляется все большая кастовая специализация поведения особей, складываются разнообразные формы полиэтизма и механизмы его регуляции. Такой процесс можно наблюдать, например, и при развитии молодой семьи черного садового или рыжих лесных муравьев. Когда в ней всего 20—30 рабочих, разделение функций между ними очень подвижно, а в крупной семье уже жестко фиксировано.
Естественный отбор требует, чтобы разделение функций в колонии обеспечивало эффективный сбор пищи, рост численности рабочих и выращивание в конечном итоге максимально возможного количества плодовитых самок и самцов. Для этого необходимо поддерживать определенные соотношения между разными полиэтическими группами и обеспечивать наибольшую производительность труда насекомых каждой из них. Как это достигается? Решением подобных вопросов занимается специальная наука — эргономика. В новой для нее области — социобиологии насекомых — она делает лишь первые шаги, но уже весьма успешные. Кое-что в связи с ее предметом мы только что обсудили, к другим вопросам вернемся позже. А пока приведем еще один пример, иллюстрирующий проблемы эргономики.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Разделение труда в клетке
Разделение труда в клетке Какова же роль отдельных клеточных образований, с которыми мы только что познакомились? Этот вопрос встал перед исследователями; вполне естественно, что его задаст и читатель, узнавший об их открытии.О защитной функции клеточных стенок мы уже
Неравенство, стратификация, касты
Неравенство, стратификация, касты Трудно остаться равнодушным при виде необычайно пестрой и многокрасочной картины разнообразия архаических обществ, лишь малой толики которых мне удалось коснуться на предыдущих страницах, Однако, сколь бы уникальны ни были быт, нравы и
1. ТЕОРЕТИЧЕСКИЕ ОСНОВАНИЯ ПРИМЕНЕНИЯ ПСИХОФИЗИОЛОГИИ ДЛЯ РЕШЕНИЯ ПРАКТИЧЕСКИХ ЗАДАЧ В ПСИХОЛОГИИ ТРУДА
1. ТЕОРЕТИЧЕСКИЕ ОСНОВАНИЯ ПРИМЕНЕНИЯ ПСИХОФИЗИОЛОГИИ ДЛЯ РЕШЕНИЯ ПРАКТИЧЕСКИХ ЗАДАЧ В ПСИХОЛОГИИ ТРУДА Пониманию необходимости и перспективности изучения психофизиологических процессов в профессиональной деятельности способствуют представления, рассматривающие
Семья и разделение труда
Семья и разделение труда Муравьиный конвейерСуровая обстановка жизни в сыпучих песках пустыни выработали у песчаного бегунка — этого крошечного создания с могучим сердцем и неисчерпаемой энергией три черты: 1 — умение ориентироваться на голых песках, на которых не
Орудия животных и орудия труда человека
Орудия животных и орудия труда человека Не вдаваясь в ход развития самой трудовой деятельности, отметим лишь еще несколько существенных моментов в дополнение к тому, что уже говорилось об орудийной деятельности обезьян.Прежде всего, важно подчеркнуть, что орудием, как
РАЗДЕЛЕНИЕ РЕСУРСОВ
РАЗДЕЛЕНИЕ РЕСУРСОВ Представьте себе, что два вида вьюрков, питающихся семенами, живут в одном и том же месте. Семян мало, и виды конкурируют между собой. Но, несмотря на конкуренцию, виды могут сосуществовать при условии, что между ними наблюдается разделение ресурсов, то
Гендерное разделение труда
Гендерное разделение труда В антропологической литературе середины XX века широкую известность получила теория «естественной взаимодополнительности полов», выдвинутая американскими социологами Т. Парсонсом и Р. Бейлзом. В рамках этой теории дифференциация мужских и