ОБЩЕСТВЕННЫЙ ЖЕЛУДОК
ОБЩЕСТВЕННЫЙ ЖЕЛУДОК
Как только оса-нянька раздаст принесенный комочек пищи, она приближается, быстро вибрируя крыльями, к отверстию каждой ячейки для того, чтобы выпить слюну, которая обильно струится изо рта заключенной в ней личинки... Но если секрет не появляется, оса захватывает голову личинки челюстями, тянет ее к себе, а затем внезапно вдавливает обратно в ячейку, опуская туда и свою голову. Эти движения стимулируют ротовые органы личинки и заставляют ее выделить слюнную жидкость. Э. Рубо. Биологические исследования одиночных и общественных ос Африки
Р. Реомюр в шестом томе «Мемуаров...», увидевшем свет в 1742 г., описал удивительное явление, к которому неоднократно обращалась наука в будущем. Наблюдая за обычными во Франции осами-полистами, он заметил, что всякий раз, когда оса накормит личинку, та выделяет изо рта капельку жидкости, которую взрослая оса поглощает как бы в награду. Уже знакомый нам Э. Рубо позднее подробнее исследовал это явление, назвав его экотрофобиозом, что в переводе с греческого прибли-

63. Кормовой обмен (трофаллаксис) между двумя рабочими германской осы (по Г. Монтанье, 1966 г.)
Донор (слева), раздвинув мандибулы, выделяет капельку корма из зобика. Реципиент (справа) поглощает эту жидкость, непрерывно постукивая усиками по голове и челюстям донора.
 64. Личинка обыкновенной осы с необычайно развитыми слюнными железами, выделяющими сахара и пищеварительные ферменты, которые поглощают рабочие (по У. Машвитцу, 1966 г.).
1 — зачаточные антенны; 2 — верхняя губа; 3 — верхние челюсти (мандибулы); 4 — нижние челюсти (максиллы); 5 — нижняя губа; 6 — общее выводное отверстие слюнных желез на нижней губе; 7 — общий выводной проток правой и левой слюнных желез; 8 — спинная ветвь (зачернена для наглядности) левой слюнной железы; 9 — брюшная ветвь левой слюнной железы; 10 — кишечник.
64. Личинка обыкновенной осы с необычайно развитыми слюнными железами, выделяющими сахара и пищеварительные ферменты, которые поглощают рабочие (по У. Машвитцу, 1966 г.).
1 — зачаточные антенны; 2 — верхняя губа; 3 — верхние челюсти (мандибулы); 4 — нижние челюсти (максиллы); 5 — нижняя губа; 6 — общее выводное отверстие слюнных желез на нижней губе; 7 — общий выводной проток правой и левой слюнных желез; 8 — спинная ветвь (зачернена для наглядности) левой слюнной железы; 9 — брюшная ветвь левой слюнной железы; 10 — кишечник.
зительно означает «семейное пищевое сожительство». Он считал, что это своеобразная форма эксплуатации взрослыми осами личинок и что именно эти капельки, получаемые от личинок, являются основной силой, удерживающей самок в гнезде и заставляющей их заботиться о потомстве, т. е. в экотрофобиозе заключена основная причина возникновения эусоциального поведения. В 1918 г. В. Вилер предложил для такого явления более удобный и получивший затем широкое распространение термин трофаллаксис, что означает «обмен пищей». Он отвергал представления Рубо об эксплуатации осами личинок и считал, что это, напротив, взаимовыгодная кооперация между взрослыми особями и потомством. Действительно, ведь осы кормят личинок и дают им гораздо больше пищи, чем получают от них. К началу XX в. ученым уже были известны многочисленные наблюдения взаимного кормления у общественных ос, пчел, муравьев, термитов. Поэтому Вилер рассматривал явление шире, называя трофаллаксисом не только кормление личинками взрослых ос, но и любые формы передачи жидкой пищи между общественными насекомыми. Короче говоря, трофаллаксис — это кормление у насекомых изо рта в рот. Партнером в трофаллаксисе может быть и личинка. Насекомое, отдающее корм, называют донором, а поглощающее его — реципиентом.
Следуя за Рубо, Вилер рассматривал трофаллаксис как силу, организующую насекомых в общество, как основную причину возникновения эусоциальности. Но, как мы увидим ниже, трофаллаксиса нет у многих наиболее примитивных общественных насекомых — стеногастрин, галиктов, аллодапин, шмелей, ряда низших муравьев. Значит, эусоциальность вполне может сформироваться и без трофаллаксиса.
Среди общественных ос трофаллаксис не наблюдали только у стеногастрин, а все остальные виды, даже такие примитивные полибии, как белоногастер, широко практикуют обмен жидкой пищей между взрослыми и поглощение личиночных выделений. Во времена Вилера не был известен состав жидкости, получаемой осами от личинок, и поэтому ученые были вынуждены ограничиваться лишь домыслами и догадками. Исследованиями в середине 60-х годов установлено, что личинки ос обладают чрезмерно развитыми по сравнению с личинками других насекомых нижнегубными или слюнными железами, которые и выделяют поглощаемый осами секрет. Как оказалось, в нем содержится около 9% сахаров. Выделения, получаемые осами от личинок, нередко составляют больше половины их суточного рациона, дополняя нектар, собираемый на цветках, и другую пищу. Осы частенько специально выпрашивают корм у личинок, прикасаясь усиками к их голове. В ответ на такое прикосновение (его можно имитировать даже кисточкой) личинка выделяет капельку сладкого секрета. Ранним утром осы «заправляются» личиночным секретом и вылетают из гнезда за кормом. Крупная личинка за одно «доение» отдает столько сахара, что его хватает для поддержания жизнедеятельности осы до полудня. Особенно важен этот секрет для ос в плохую погоду, когда на цветках провизии не найти, а на обогрев гнезда энергии нужно много.
Кроме сахаров в личиночном секрете содержится немного ферментов, расщепляющих белки и углеводы. Их всего около 0,5%, но и в таком количестве они действуют достаточно эффективно. Осы смешивают личиночный секрет с пережевываемой добычей, и его ферменты подготавливают пищу к усвоению личинками. Слюнные железы взрослых ос выделяют очень мало протеаз, и этот недостаток восполняют ферменты слюны личинок. Получается в некотором роде разделение функций: личинки поставляют ферменты, а осы совершают механическую работу по пережевыванию добычи и пропитке ее ферментами и дают личинкам в готовом к проглатыванию виде!
Все эти факты вместе подтверждают точку зрения Вилера, увидевшего в трофаллаксисе между осами и личинками выгодную обеим сторонам кооперацию. Не исключено, что выделения личинок несут и еще одну функцию: они позволяют избавляться от излишней влаги в гнезде. Ведь выделение жидких экскретов обычным для насекомых путем (через мальпигиевы сосуды и анальное отверстие) могло бы привести к намоканию и порче бумажных сотов. Примитивные муравьи рода амблиопоне и некоторые другие понерины никогда не обмениваются кормом. У нотомирмеции, мирмеций и большинства понерин трофаллаксис наблюдается редко и протекает недостаточно организованно: позы муравьев при обмене кормом могут быть самыми различными (т. е. не стереотипными, как у высших муравьев), кормовые контакты непродолжительны. У высших же муравьев трофаллаксис - обычнейший элемент поведения. Поэтому уже давно сделан вывод, что степень развития трофаллаксиса является одним из важнейших показателей продвинутости социальной организации вида. Но нет правил без исключения, и эта закономерность иногда нарушается в эволюции: у ряда высших муравьев трофаллаксис отсутствует. Примером могут служить муравьи родов афеногастер, мессор, погономирмекс. Муравьи довольно примитивного рода афеногастер, вероятно, никогда не имели трофаллаксиса. А вот у жнецов он не появился, наверное, потому, что пища этих муравьев — в основном твердые зерна и насекомые. Муравьи-кочевники питаются тоже исключительно твердой пищей. Их личинки и царица никогда не получают корм изо рта рабочих. А между рабочими кормовые обмены наблюдаются крайне редко. Значит, возможно все-таки достижение весьма высокой ступени социальной организации, на которой находятся кочевники, без развития трофаллаксиса.
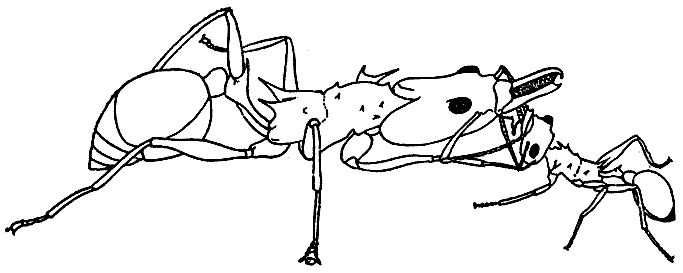
65. Трофаллаксис между рабочими муравья дацетон армигерум (по Э. Вильсону, 1962 г.).
Более крупный рабочий выпрашивает пищу, дотрагиваясь передними ногами до головы маленького, и поглощает появившуюся изо рта партнера жидкость.
Вообще особенности питания муравьев существенно влияют на встречаемость трофаллаксических контактов в их колониях. Так, почти все формицины и долиходерины — заядлые «пастухи»: падь тлей и других сосущих насекомых занимает в их диете важнейшее место. Некоторые виды питаются исключительно жидкой пищей. Потому-то у муравьев этих двух подсемейств трофаллаксис — важнейшая форма поведения. А вот в пищевом рационе муравьев подсемейства мирмицин жидкая пища составляет небольшую долю, даже тлей эти муравьи посещают не очень активно, потому и трофаллаксис мы можем наблюдать в их гнездах не столь часто.
Многие ученые видели, как личинки муравьев выпускают изо рта капельки жидкости, которые слизывают рабочие. Нередко муравьи даже выпрашивают эти капельки, постукивая антеннами по ротовым органам и голове личинки. Царица амблиопон челюстями сдавливает шею личинки, заставляя ее выделить изо рта порцию секрета, которую затем поглощает. К сожалению, до сих пор никто не исследовал состав продуцируемой личинками жидкости. Скорее всего, это продукт слюнных желез. Но пока мы ничего не можем сказать о его роли в жизни муравьев.
Точно так же недостаточно ясным остается пока значение капелек, выделяемых личинками из анального отверстия. Их тоже поглощают рабочие. В большинстве случаев это, конечно, просто экскреты из мальпигиевых сосудов (иногда хорошо видно, как в капельке плавают кристаллы мочевой кислоты). Но не исключено, что в них могут быть и какие-то необходимые муравьям вещества. Не так давно в анальных выделениях личинок фараонова муравья были обнаружены аминокислоты и ферменты протеазы. Это позволяет предполагать, что анальная жидкость личинок может содержать не только вредные продукты обмена веществ.
Среди общественных пчел трофаллаксиса нет у аллодапин и галиктов. У большинства шмелей исследователям не удавалось заметить обмен кормом, лишь у некоторых видов крайне редко наблюдали неупорядоченные трофаллаксисы. Что же касается безжалых и медоносных пчел, то трофаллаксис, является у них важнейшим элементом поведения. У термитов, как уже говорилось во втором разделе, две формы трофаллаксиса: оральная, когда пища выделяется изо рта, и анальная, основанная на выведении корма из заднепроходного отверстия. Анальный трофаллаксис есть только у низших термитов и служит в первую очередь для передачи кишечных симбионтов. Кроме того, этим путем полупереваренная пища из толстой кишки одного термита окончательно усваивается другими членами колонии. При оральном трофаллаксисе рабочие термиты могут выделять корм двух типов. Секрет слюнных желез рабочие дают царской паре, крылатым термитам и личинкам самых младших возрастов; он составляет единственную пищу этих насекомых. Более старшим личинкам, нимфам, солдатам и другим рабочим они отрыгивают содержимое зобика или средней кишки, т. е. частично переваренную пищу. У высших термитов секретом слюнных желез рабочих питаются личинки всех возрастов.

66. Маленький рабочий термита-грибовода макротермес белликозус кормит крупного солдата, выделяя жидкую пищу изо рта (по П. Грассе, 1949 г.).
При трофаллаксисе насекомые принимают стереотипные позы, характерные для семейства, рода и даже вида. Мы еще поговорим об этом, когда речь пойдет о способах общения. Если существует поведенческая иерархия, то корм всегда передается от подчиненного насекомого доминирующему; донора и реципиента легко различить. У пчел при трофаллаксисе донор лишь слегка выдвигает хоботок вперед и вниз и выделяет из него каплю корма, а пчела-реципиент почти полностью вытягивает хоботок вперед, высасывая эту каплю. А вот у муравьев или термитов различить донора и реципиента не всегда просто, если только не было возможности увидеть, кто именно из партнеров отрыгнул каплю корма. Впрочем, у целого ряда видов муравьев решить эту задачу несложно, поскольку донор широко раскрывает челюсти, а реципиент держит их сомкнутыми.
Недавно во Франции была разработана установка, позволяющая измерять радиоактивность отдельных частей тела крупных муравьев в процессе трофаллаксиса. Когда рабочих кампонотуса накормили раствором сахара с радиоактивным золотом, то удалось при обменах кормом между ними по уменьшению радиоактивности брюшка донора и увеличению ее у реципиента измерять скорость трофаллаксиса. На этой же установке было обнаружено и ранее неизвестное явление — реверсии трофаллаксиса. Выяснилось, что нередко в процессе обмена кормом донор и реципиент вдруг меняются ролями и корм начинает поступать в зобик того муравья, который только что его отдавал. Иногда в процессе одного трофаллаксиса роли муравьев могут поменяться даже несколько раз. Пока непонятно, какой смысл заключен в подобных реверсиях.
Основная функция трофаллаксиса — распределение пищи между всеми насекомыми в колонии. Важнейшая особенность системы такого распределения — ее открытость: каждый рабочий, солдат, личинка, крылатая самка или самец могут вступить в обмен кормом с любым членом колонии. Корм обычно отдают по первому требованию и получают при необходимости от других. Поэтому совокупность зобиков всех насекомых в гнезде составляет как бы общественный желудок, в котором корм хранится и перераспределяется путем трофаллаксиса. Впервые представление об общественном желудке ввел в науку О. Форель в 1878 г.
У видов, не обладающих трофаллаксисом, нет и общественного желудка. Некоторое исключение в этом отношении составляют только шмели. В их гнезде есть медовый горшок — временное хранилище резервных запасов меда. Шмели то пополняют запасы, то используют их. Когда одному шмелю скормили немного радиоактивного сиропа, следы радиоактивности вскоре обнаружились у многих особей из этого гнезда, поскольку они, как говорится, «хлебали из одной лохани» — медового горшка. Не все виды общественных насекомых в равной мере способны сохранять в общественном желудке резервные запасы корма. В значительной степени это обусловлено строением пищеварительной системы. У всех насекомых зобик представляет собой расширение передней кишки и связан с ротовым отверстием пищеводом. Следующим отделом кишечника является мышечный желудок, или провентрикулюс. У термитов он снабжен зубчиками, которые дополнительно перетирают проглатываемый корм. У перепончатокрылых единственная функция провентрикулюса — управление поступлением пищи из зобика в собственно желудок. Зобик покрыт изнутри кутикулой, поэтому пища в нем не переваривается и не усваивается. Значит, когда пчела или муравей проглатывает корм, то он попадает как бы в общественный желудок и пока не является «собственностью» данного насекомого. Ведь из зобика корм по первому требованию может быть отдан любому другому. Выходит, что, так сказать, «индивидуальным ртом» муравья, термита или пчелы служит провентрикулюс, ведь насекомое питается, только когда клапаны мышечного желудка открываются и пища поступает в собственно желудок.

67. Пищеварительная система рыжей мирмики (по Ш. Жанэ, 1894 г.).
1 — ротовая полость; 2 — подротовой карман; 3 — глотка; 4 — проток слюнной железы; 5 — слюнная железа; 6 — пищевод; 7 — зобик; 8 — мышечный желудок (провентрикулюс); 9 — железистый желудок; 10 — толстая кишка; 11 — прямая кишка; 12 — выделительные трубочки (мальпигиевы сосуды).
У всех низших муравьев, а также у мирмицин провентрикулюс устроен так, что просвет его закрывается сокращением кольцевой мышцы. Для того чтобы держать пищу в зобике, муравей должен непрерывно напрягать ее. Видимо, по этой причине названные муравьи не могут длительное время хранить в общественном желудке значительные запасы пищи.
У формицин и долиходерин провентрикулюс имеет совершенно иное строение: его хитиновые клапаны постоянно плотно сомкнуты и открываются только при сокращении специальных мышц. Поэтому хранение пищи в зобике не требует особых усилий. Виды муравьев этих двух подсемейств обладают мягким растяжимым брюшком и способны приносить в гнездо и удерживать в зобиках намного больше пищи, чем все прочие муравьи. Длительное хранение в гнезде жидкой пищи у них весьма обычное явление. Стоит дать содержащимся в лаборатории формикам или лазиусам раствор сахара либо меда, как среди них окажется множество рабочих с сильно раздутыми брюшками. Зобики их до отказа заполнены сладким кормом. Эти распухшие муравьи могут в течение многих недель хранить внутри себя общие запасы корма. Именно среди формицин и долиходерин встречаются так называемые медовые муравьи, в гнездах которых можно найти «медовые бочки» — рабочих с полупрозрачным брюшком, раздувшимся от сладкой жидкости до размера крупной вишни. Их называют плерэргатами.
Американские медовые муравьи принадлежат к роду мирмекоцистус и обитают в прериях. В их глубоких почвенных гнездах можно найти до 1500 плерэргатов. Каждый сохраняет около 0,4 г «меда», что составляет до 90% массы его тела. Запасы меда в муравейнике могут достигать 600 г. Муравьи выходят из гнезд по ночам и собирают падь тлей, а затем отдают ее плерэргатам, которые неподвижно висят, уцепившись ножками или челюстями за стенки глубоких камер муравейника. При этом зобик их так раздут, что спинные и брюшные полукольца занимают на брюшке совсем небольшую площадь, остальное — это межсегментарная мембрана. Когда возникает потребность, плерэргаты отрыгивают корм любой попросившей его особи. Так и проходит вся жизнь муравья-бочки, она не балует его разнообразием!
Муравьиный мед не очень сладкий (содержит не более 50% сахаров), но, как и пчелиный, включает в основном простые сахара и оксидазу глюкозы, стерилизующую мед за счет выделения перекиси водорода. Некоторые плерэргаты хранят почти чистую, слегка подслащенную воду. Запас воды в тех засушливых районах, где живут медовые муравьи, тоже очень кстати!
В сухих районах Австралии обитают муравьи кампонотус инфлатус, плерэргаты которых почти столь же огромны, как у американских медовых муравьев, а также виды родов мелофорус и лептомирмекс, обладающие «медовыми бочками» с менее растяжимым брюшком. Муравьи с такими плерэргатами встречаются и в других районах мира: это тапинома в Южной Америке, аноплолепис и плагиолепис в Африке, проформика и пренолепис в Азии. На юге СССР живет несколько видов рода проформика. В гнездах этих муравьев постоянно есть плерэргаты. Брюшко их увеличено не очень сильно, так что плерэргаты свободно ходят по гнезду. Но все же в зобике каждого хранится около 10 мг сладкого корма, а этого достаточно для питания сотни муравьев в течение месяца.
Скорость перемешивания пищи в общественном желудке удивительно велика. Когда шести пчелам скормили 20 мл сиропа с радиоактивным фосфором, то через пять часов 27% рабочих в улье имели радиоактивную метку, а через сутки их было уже 55%. Значит, всего за сутки принесенный шестью фуражирами корм попал в зобики десятков тысяч пчел. Сколько же потребовалось для этого актов трофаллаксиса! В подобном же эксперименте один рабочий бурого лесного муравья набрал полный зобик сиропа с радиоактивным йодом. Через 27 часов все 70 муравьев, обитавших в лабораторном гнезде, были радиоактивными. Многочисленные опыты с весьма сходными результатами неоднократно проводились и с другими видами общественных пчел, ос, муравьев, термитов.
Но в то же время эксперименты обнаружили и некоторые различия. Виды, в колониях которых обмен кормом происходит в среднем реже, отличаются, как и следовало ожидать, меньшей скоростью перемешивания пищи в общественном желудке. Например, у мирмицин принесенный в гнездо корм значительно медленнее распределяется между рабочими, чем у формицин или долиходерин. Различия в скоростях перемешивания корма могут быть связаны и с его природой. Доставленная фуражирами рыжих лесных муравьев падь очень быстро распределяется в основном среди внегнездовых рабочих, а гемолимфу добычи фуражиры передают прежде всего рабочим-нянькам для личинок, и она распространяется среди муравьев в гнезде в 2—3 раза медленнее, чем сладкий корм.
Не все насекомые в колонии совершенно равноценны в системе трофаллаксиса. Царицы обычно только получают корм, но никому не отдают его. Впрочем, у примитивных видов общественных насекомых царицы могут иногда выступать в роли доноров. Такое наблюдается даже у некоторых муравьев, в частности у мирмик. Крылатые самки и самцы, солдаты, личинки я нимфы также чаще являются реципиентами.
Значение трофаллаксиса не ограничивается распределением пищи и перемешиванием ее в общественном желудке, хотя это, несомненно, наиболее важная его функция. Огромна его роль и в регуляции потребления пищи и фуражировочной деятельности насекомых. Спрос в колонии на пищу каждый ее член ощущает, так сказать, на «собственном брюхе». Когда корма не хватает, зобик фуражира пустеет, и это прибавляет ему энергии в поисках добычи.
Трофаллаксис может применяться для передачи информации. Например, при большой потребности в пищи фуражира «атакует» сразу несколько рабочих, выпрашивающих у него корм. Это служит сигналом к усилению фуражировочной деятельности. При мобилизации на пищу пчелы и муравьи нередко используют трофаллаксис для передачи информации о качестве корма.
Еще одна функция трофаллаксиса связана с распространением в колонии феромонов. Установлено, что феромоны царской пары попадают ко всем термитам именно при трофаллаксисе, вместе с пищей. Существует предположение, что таким же путем могут переходить от особи к особи феромоны и у общественных перепончатокрылых. Во всяком случае, трофаллаксис значительно ускоряет, например, передачу феромона пчелиной матки, поскольку способствует более тесным контактам между рабочими, во время которых феромон как бы «перетекает» с одной пчелы на другую.
Голодные личинки ос, выделяя капельки сладкой слюны, привлекают этим рабочих ос. Расчет здесь простой: оса выпьет капельку, обратит внимание на голодную личинку да и покормит ее! В этом случае трофаллаксис обеспечивает функцию привлечения.
И, наконец, трофаллаксис может снижать агрессивность при враждебных столкновениях. Вспомним, как подчиненная оса выделяет капельку корма и этим «усмиряет» доминирующую. Так бывает и у муравьев, но уже в территориальных взаимоотношениях. Когда муравей случайно или целенаправленно попадает во владения соседней колонии и вдруг оказывается окруженным несколькими враждебно настроенными чужаками, он поступает весьма находчиво: отрыгивает капельку корма и предлагает ее врагам. И чем агрессивнее они себя ведут, тем настойчивее прельщает их кормом атакованный муравей. Нередко это спасает храбреца. Враги поглощают пищу и явно «добреют», начинают чиститься, не обращая внимания на нарушителя границы, который успевает улизнуть. Такое поведение наблюдали даже между муравьями разных видов. Настоящий межвидовой трофаллаксис!
Как могло возникнуть в эволюции столь сложное поведение — трофаллаксис? Вероятнее всего, это произошло на основе случайного вначале слизывания капелек пищи, попавших на тело фуражиров. Потом насекомые могли уже облизывать более целенаправленно не только тело, но и ротовые органы, на которых были остатки корма. Отсюда недалеко и до специального отрыгивания корма донором и поглощения его реципиентом. Сначала такой кормовой обмен был неупорядоченным, как у некоторых современных примитивных муравьев. Затем постепенно сформировались стереотипные позы выпрашивания, передачи и приема корма, а это значительно повысило результативность всего процесса. Косвенным подтверждением такого эволюционного пути является тот факт, что трофаллаксис есть только в тех группах общественных насекомых, где практикуется взаимное облизывание. Исключение составляют осы-полисты, у которых взаимное облизывание наблюдается очень редко, но трофаллаксис весьма обычен.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 9. Общественный порядок без любви
Глава 9. Общественный порядок без любви … и в сердце вечный хлад Гёте Противопоставление анонимной стаи и личной связи, которым мы закончили предыдущую главу, означает лишь, что эти два механизма социального поведения в весьма значительной степени исключают друг друга;
Общественный желудок
Общественный желудок Взаимные угощенияСтрогий осенний лес Западной Сибири. На фоне золотых осинок ели кажутся черными, и муравейник — тоже черный среди пожелтевшей травы. Весь день была пасмурная погода, дул холодный ветер, и вот сейчас, когда в серой пелене облаков
Разные типы земледелия дают разный общественный результат
Разные типы земледелия дают разный общественный результат Из перечисленных выше четырех исходных типов земледелия первый и последний не имеют препятствий для индивидуального владения землей. Изолированные от нападений извне группы в таких условиях могли легко
Взаимное кормление, или общественный желудок
Взаимное кормление, или общественный желудок У общественных насекомых — муравьев, пчел, ос, термитов существует обычай кормить друг друга пищевыми отрыжками. Обоюдный обмен отрыжками происходит не только среди взрослых, но даже между взрослыми и личинками. Обмен