СПОСОБЫ ОБЩЕНИЯ
СПОСОБЫ ОБЩЕНИЯ
Так муравьи, столкнувшись где-нибудь. Потрутся рыльцами, чтобы дознаться, Быть может, про добычу и про путь... А. Данте. Божественная комедия
В прошлом мало у кого возникали сомнения в том, умеют ли муравьи или пчелы разговаривать друг с другом. Это казалось очевидным. Как же иначе они могли бы действовать столь разумно и согласованно? Споры шли лишь о том, общаются ли насекомые с помощью звуков или же их язык основан на иных физических явлениях. Сэр Дж. Лаббок в 70-х годах прошлого столетия затратил массу усилий, чтобы выяснить, слышат ли пчелы, осы и муравьи. «Я снова и снова, — писал он, — испытывал их самыми громкими и резкими шумами, какие мог только производить, употребляя копеечную дудку, собачий свисток, скрипку, равно как и издавая самые пронзительные и ошеломляющие звуки, на какие только способен мой голос; но все это не производило на них никакого влияния. В то же время я тщательно избегал выводить из этого, что они глухи, хотя, кажется, что их род слуха сильно отличается от нашего».[10]
Теперь-то мы знаем, что муравьи и термиты совершенно глухи к обычным звукам, являющимся колебаниями воздуха; у них нет специальных органов для восприятия этих звуковых волн. Но зато все насекомые отлично чувствуют вибрацию твердого субстрата, на котором сидят; ее воспринимают так называемые подколенные органы, расположенные на голенях всех шести ножек. Поэтому насекомые «слышат» ногами. Известно, что звуковые волны, распространяющиеся в воздушной среде, в основном отражаются от твердых предметов. Вот почему оказались тщетными все усилия Лаббока!
В 70-е годы нашего столетия Е. К. Еськов выяснил, что пчелы и осы в отличие от муравьев и термитов способны воспринимать некоторые звуки из воздушной среды специальными волосками (так называемыми фонорецепторами) на голове. Однако чтобы общаться с помощью звуков, надо уметь не только слышать, но и издавать их.
Огромное большинство насекомых, в том числе и общественных, не имеет специальных органов, производящих звуки. Пчелы и осы используют для этой цели крыловую мускулатуру. При быстрых сокращениях мышц хитиновые части наружного скелета груди вибрируют, издавая жужжащие звуки. «Голосовой диапазон» пчел и ос довольно широк, они могут жужжать на разный лад — низко или высоко, отрывисто или продолжительно. Изучение спектров издаваемых ими звуков позволило установить, что осы и особенно медоносные пчелы используют в общении довольно много звуковых сигналов, хорошо различающихся по физическим параметрам. Ниже мы рассмотрим несколько примеров звуковой коммуникации у этих насекомых.
Некоторые муравьи и термиты способны издавать звуки трением друг о друга твердых частей наружного скелета. Звуковые аппараты, базирующиеся на этом принципе, называются фрикционными. У большинства муравьев подсемейства мирмицин на верхнем заднем крае второго членика стебелька есть направленный назад и вниз заостренный кантик (или «коготок»), а на верхней стороне основания брюшка расположено ребристое поле — участок кутикулы с мелкими поперечными морщинками. Когда брюшко движется относительно стебелька вверх или вниз, «коготок» царапает по этим морщинкам и раздается очень слабое стрекотание, или, как выражаются энтомологи, стридуляция. Ее можно расслышать, прижав ухо к лабораторному формикарию с возбужденными мирмиками. Стридуляционный аппарат мирмеций и понерин устроен примерно так же, но стрекочут эти муравьи гораздо громче мирмик, их звуковые сигналы хорошо слышны с расстояния в несколько метров. Нотомирмециястридулирует значительно тише, фрикционный аппарат находится у нее на нижней стороне брюшка. Некоторые термиты могут издавать скрипящие звуки трением головы о переднегрудь.
Но все же множество муравьев и термитов не имеет специальных органов, производящих звуки. Поэтому они просто ударяют о субстрат челюстями, головой или брюшком. Такой стук, распространяющийся в виде колебаний субстрата, воспринимается другими насекомыми на порядочном расстоянии.
П. Гюбер был первым среди ученых, обратившим внимание на роль усиков в общении между муравьями. «Я часто видел, — писал он, — как они пользовались усиками на поле битвы с целью указать на приближение опасности и отличить себя от неприятеля; среди муравейника, чтобы возвестить о появлении солнца, имеющего такое благоприятное влияние на развитие личинок; во время своих путешествий и передвижений, чтобы указывать друг другу дороги; отправляясь на добычу корма; для определения времени начала похода и т. д.».[11]
В 1899 г. известный немецкий ученый, монах ордена иезуитов Э. Васманн, посвятивший многие годы исследованиям муравьев и термитов, объявил, что ему удалось обнаружить у них язык. Словами этого языка, по его мнению, являются прикосновения усиков-антенн одного насекомого к телу, голове и антеннам другого. Васманн описал целый ряд подобных сигналов-прикосновений, имеющих якобы вполне определенный смысл, и утверждал, что муравьи и термиты способны передавать очень сложные сообщения, эквивалентные фразам нашего языка. Концепция «антеннального языка», или «антеннального кода», Васманна получила широкое распространение. Есть у нее сторонники и в наше время. Насколько близка к истине эта точка зрения?
Сейчас прикосновения одного насекомого к другому антеннами, ножками и иными частями тела, передающие какую-то информацию, принято называть тактильными (осязательными) сигналами. Ученые, пытающиеся их изучать, сталкиваются с огромными трудностями. Движения насекомых очень быстры, и человеку порой невозможно даже «схватить» их глазом, а тем более точно описать. Для этого используют скоростную киносъемку, которой не было во времена Васманна. Но мало подробно описать тактильные сигналы, надо выяснить их значение, если таковое имеется. А как это сделать? Допустим, мы видим, что после того или иного сигнала насекомое, которому он адресован, совершает определенное действие и так происходит в большинстве случаев. Можно ли на этом основании утверждать, что данный тактильный сигнал передает информацию о необходимости именно такого действия? Конечно, нет! Мы же не исключили возможности одновременной сигнализации с помощью запахов, звуков или каким-то иным путем. Например, Васманн интерпретировал очень быстрые постукивания антеннами как сигнал тревоги. Но в его время еще не было известно, что возбужденный муравей может выделить химическое вещество со специфическим запахом — феромон тревоги, который и сигнализирует об опасности прежде всего.
Как исследовать независимо роль химических и тактильных сигналов в этой ситуации? Ведь тактильные сигналы тоже могут нести определенную информацию, дополняющую химические сигналы. Но как ее выявить? Создать модель муравья, воспроизводящую характерные прикосновения антенн, но лишенную феромонов? Это могло бы очень приблизить нас к истине. Но пока никому не удалось подобное тончайшее моделирование — слишком уж велики технические трудности. Правда, кое-что с моделями и муляжами уже получилось, но на более крупных насекомых — пчелах и осах. И об этом мы еще поговорим.
Все эти объективные трудности явились причиной того, что тактильная коммуникация у общественных насекомых изучена пока хуже всех остальных форм общения. Васманн и его последователи интерпретировали наблюдаемые ими тактильные сигналы чисто умозрительно, приписывая им то значение, которое, как они полагали, наиболее соответствует ситуации. Понятно, что подобные построения сторонники строгих и объективных методов экспериментальной науки встречали с недоверием. До сих пор концепция антеннального языка в понимании Васманна и его единомышленников не получила сколько-нибудь серьезного подтверждения. Из этого, конечно, не следует, что тактильные сигналы не играют никакой роли в процессах коммуникации у общественных насекомых. Напротив, их значение в общении необычайно велико.
Но прежде чем приводить доказательства этого утверждения, отметим, что язык насекомых можно изучать, не только анализируя используемые при общении сигналы. Можно попытаться выяснить, что вообще способны «сказать» друг другу насекомые? В состоянии ли они передать настолько сложную информацию, которой можно обмениваться лишь при наличии языка? Ответить на этот вопрос проще, чем стараться выяснить смысл отдельных сигналов. Какие же результаты достигнуты на этом направлении? Анализируя имеющиеся в науке данные, следует признать, что до настоящего времени не получены строгие доказательства существования у общественных насекомых языка, хотя бы в некоторой степени подобного человеческому. Конечно, у общественных насекомых есть довольно сложные формы передачи информации, но основаны они на совершенно иных принципах, чем у людей.
Итак, тактильные сигналы вряд ли могут передавать сложные сообщения. И далеко не всегда прикосновения усиками вообще выполняют сигнальную функцию. Насекомые прикасаются друг к другу в первую очередь с целью ориентации, чтобы узнать по запаху, кто находится перед ними — друг или враг, рабочий, солдат, царица или личинка. Усики насекомое гораздо чаще использует именно для получения информации, а не для передачи. В каких же случаях доказано сигнальное значение прикосновений?
Прежде всего без тактильной коммуникации невозможно взаимодействие насекомых при трофаллаксисе. Понаблюдаем за тем, как рабочие рыжей мирмики кормят личинок. Муравей-нянька заранее не знает, какие личинки, во множестве лежащие в камере гнезда, готовы принять от него корм. Он может определить это только путем общения с каждой личинкой. Рабочий подходит к личинке и слегка постукивает кончиками усиков по ее голове и ротовым органам. Если личинка голодна, она отвечает шевелением челюстей или движениями головы вверх-вниз. Уловив усиками движения личинки, муравей тут же отрыгивает ей в рот капельку пищи. Но личинка, которую недавно накормили, не реагирует на прикосновения усиков рабочего. Тогда он начинает ее облизывать. Быстро движется язычок муравья, тщательно обрабатывая голову и ротовые органы личинки. Время от времени рабочий прерывает облизывание, постукивает усиками по голове личинки и опять принимается ее облизывать. В конце концов ему удается «растормошить» личинку и она подает муравью сигнал, «соглашаясь» принять корм. Нередко можно наблюдать и обратную ситуацию: личинка настолько голодна, что реагирует сильными движениями головы и челюстей на случайные касания проходящих мимо муравьев и даже на сотрясение гнезда. Муравьи ощущают эти движения личинки, когда ощупывают усиками ее тело и вознаграждают ее капелькой корма.
Таким образом, кормление личинки муравьем невозможно без довольно сложного общения между ними, основанного на обмене тактильными сигналами. Подобную же картину можно видеть и у пчел-аллодапин. Ш. Сакагами наблюдал, как личинки видов рода экзонеура движением челюстей и головы сигнализируют о том, что им нужен корм; в ответ на это пчелы захватывают их челюстями и отрыгивают им в рот прозрачную жидкость.
Трофаллаксису между взрослыми насекомыми также предшествует обмен тактильными сигналами. Для всех видов общественных насекомых характерны специфические стереотипные позы выпрашивания и предложения пищи, которые, вероятно, тоже имеют сигнальное значение. Медоносная пчела выпрашивает корм у другой особи, всовывая хоботок между ее челюстями и одновременно постукивая усиками по ее голове и ротовым органам. Поза предложения пищи выражается в небольшом выдвижении хоботка вперед-вниз и отрыгивании капли корма. Отделенная от тела голова вполне заменяет живую пчелу — другие рабочие будут предлагать ей корм и пытаться выпрашивать у нее. Голова без усиков гораздо менее эффективна. Но если усики имитировать проволочками, то голова вновь станет объектом предложения и выпрашивания корма. Некоторую роль в этом поведении играет и запах: небольшой шарик из любого материала, лишенный даже подобия антенн, но обладающий запахом пчелиной головы, иногда побуждает окружающих пчел выпрашивать или предлагать корм.
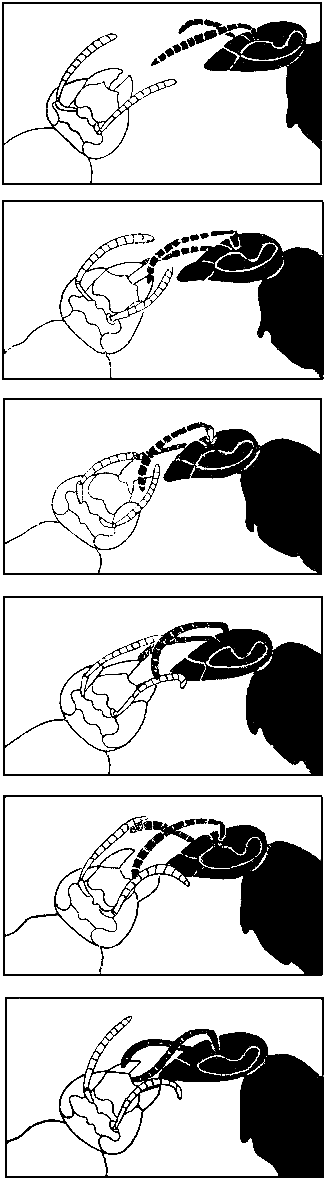 70. Тактильные сигналы при трофаллаксисе у германской осы (по Г. Монтанье, 1966 г.).
Сверху вниз: оса, выпрашивающая корм, т. е. реципиент (справа), подходит к донору и прикасается кончиками усиков к его мандибулам; в ответ донор приближает свои усики к усикам реципиента и приоткрывает челюсти; реципиент начинает слегка постукивать усиками по ротовым органам донора, а затем, немного приблизившись, и по передней части его головы; в этот момент донор, еще шире раскрыв челюсти, отрыгивает капельку пищи, которую реципиент постепенно поглощает, продолжая постукивать усиками по голове и мандибулам донора.
70. Тактильные сигналы при трофаллаксисе у германской осы (по Г. Монтанье, 1966 г.).
Сверху вниз: оса, выпрашивающая корм, т. е. реципиент (справа), подходит к донору и прикасается кончиками усиков к его мандибулам; в ответ донор приближает свои усики к усикам реципиента и приоткрывает челюсти; реципиент начинает слегка постукивать усиками по ротовым органам донора, а затем, немного приблизившись, и по передней части его головы; в этот момент донор, еще шире раскрыв челюсти, отрыгивает капельку пищи, которую реципиент постепенно поглощает, продолжая постукивать усиками по голове и мандибулам донора.
Тактильная коммуникация при трофаллаксисе у германской осы значительно сложнее. Оса выпрашивает корм у предполагаемого донора, постукивая усиками по его голове и челюстям. Во время трофаллаксиса реципиент продолжает постукивание — в противном случае донор прервет передачу корма. Поэтому оса без антенн или даже лишенная их кончиков остается голодной. Отделенной от туловища голове осы не предлагают пищу. Но если антенны заменить проволочками, совершающими от 20 до 100 колебаний в секунду, то такая модель головы реципиента побуждает окружающих ос предлагать ей корм. Однако уже через несколько секунд оса-донор прерывает контакт и уходит, поскольку модель не в состоянии правильно воспроизвести прикосновения антенн к голове донора. Интересно, что самцы ос не могут выпрашивать пищу у рабочих, видимо, потому, что кончики их усиков, загнутые крючком, совершенно непригодны для подачи соответствующих тактильных сигналов.
Лишенные антенн муравьи обычно тоже неспособны попросить пищу у собратьев по гнезду. Выпрашивающий корм рабочий черного муравья-древоточца поглаживает усиками голову и ротовые органы предполагаемого донора и продолжает делать это в течение всего трофаллаксиса. Усики его движутся в противофазе: правый усик прикасается к левой стороне головы донора и скользит по ней до правой мандибулы, затем левый усик проделывает тот же путь, но справа налево. Донор вначале чаще всего вытягивает антенны над головой реципиента, а затем во время передачи корма двигает ими довольно беспорядочно, но не прикасается к реципиенту. Сходным образом ведут себя при тро-фаллаксисе и муравьи других видов: реципиент постукивает антеннами по голове и челюстям донора, а тот отводит антенны в стороны.
У многих видов предлагающий пищу муравей принимает характерную позу. Рыжая мирмика, например, широко раскрывает челюсти и замирает, удерживая на язычке отрыгнутую капельку корма. Сходным образом ведут себя и другие муравьи. Нередко при этом поза муравья изменяется в зависимости от того, сколь сильно он «желает» отдать корм. Рабочий красногрудого муравья-древоточца при настойчивом предложении корма приоткрывает челюсти, поворачивает голову примерно на 90° и приближает ее к голове предполагаемого реципиента, одновременно поглаживая его усиками. Если муравей, которому предлагают корм, не спешит его взять, донор сильно наклоняется на бок, поворачивает голову еще на 90° и подставляет ее под челюсти партнера. Обычно столь «настоятельная просьба» приводит к желаемому результату.
Насекомое, которому адресована сигнальная поза, воспринимает ее зрением и осязанием, т. е. прикасаясь антеннами к партнеру. Поэтому такие позы относят ктактильно-зрительным сигналам. Но если дело происходит в гнезде, где света нет, то тактильное восприятие становится основным. До сих пор мы не знаем, насколько зрительное восприятие важно в тех или иных случаях. Количество известных сигнальных поз также невелико. Когда муравьи некоторых видов рода лептоторакс переселяются в новое гнездо, многие рабочие принимают перед носильщиками позу «предложения»: муравей поворачивается задним концом тела и немного подгибает брюшко, выставляя членики стебелька, за который его и берет носильщик.
Среди тактильно-зрительных сигналов гораздо более распространены не позы, а сигнальные движения, совершаемые всем телом или его частью и нередко сопровождаемые прикосновениями усиками, ножками или другими органами к окружающим насекомым. Такие сигналы являются комплексными, их воспринимают тактильно, по сотрясению субстрата, а при наличии света — зрением. Когда муравей-носильщик рыжей мирмики намеревается транспортировать рабочего в новое гнездо, он совершает в его сторону несколько коротких бросков, прикасаясь каждый раз к приглашаемому муравью антеннами и даже слегка толкая его челюстями. Рабочий обычно реагирует на это, подгибая ноги и антенны, а затем, когда носильщик захватывает его за голову, окончательно принимает стереотипную позу переносимого муравья. Разнообразные сигнальные движения широко используют разные виды общественных насекомых для мобилизации, предупреждения об опасности и в других случаях. Очень сложными сигнальными движениями являются так называемые танцы медоносных пчел. Обо всем этом мы еще поговорим специально.
Существуют и чисто зрительные сигналы, воспринимаемые на значительном расстоянии. Но роль их исследована плохо. Пока известен лишь один достоверный случай использования зрительной коммуникации при мобилизации на пищу у муравьев. Пчелы-сторожа, дежурящие у входа в улей, хорошо отличают по полету и характеру движений пчел-сборщиц от других насекомых, а также от пчел-воровок, пытающихся проникнуть в улей за медом. Воровки обычно движутся несколько иначе, чем сборщицы, как-то неуверенно, что сразу же замечают сторожа. Но в данном случае вряд ли можно говорить о сигнализации в истинном смысле слова.

71. Железы внешней секреции (по Э. Вильсону, 1971 г.).
А — медоносная пчела; Б — муравей иридомирмекс хумилиса. 1 — мандибулы; 2 — мандибулярная железа; 3 — гипофарингеальная (максиллярная) железа; 4 — глоточная (фарингеальная) железа; 5 — головной отдел нижнегубной (лабиальной) железы; 6 — грудной отдел лабиальной железы (у муравья — вся железа); 7 — гипостомальная (защечная) железа: 8 — метаплевральная железа; 9 — железа Насонова; 10 — жало; 11 — железа Кожевникова; 12 — железа Дюфура; 13 — резервуар ядовитой железы; 14 — ядовитая железа; 15 — восковые железы; 16 — железы в задней кишке; 17, 18 — анальная железа и ее резервуар; 19 — железа Павана.
Многие ученые считали, что характерные позы тревоги и угрозы, принимаемые муравьями или медоносными пчелами, окружающие насекомые воспринимают зрением и реагируют на них, как на сигнал опасности. Но ведь одновременно насекомое выделяет и феромон тревоги, являющийся сигналом опасности. Сигнальная функция феромонов тревоги очень хорошо известна, а сигнальная роль поз тревоги и угрозы не доказана экспериментально. Так что вопрос о чисто зрительном восприятии сигнальных поз остается открытым.
Чрезвычайно широко распространена у общественных насекомых химическая коммуникация с помощью сигнальных веществ — феромонов, выделяемых во внешнюю среду специальными железами. Таких желез у общественных насекомых известно очень много.
Сигнализация посредством феромонов исследована, пожалуй, лучше всех остальных форм общения в связи с относительной несложностью и достоверностью используемых методов. Например, довольно просто при должной сноровке выделить из тела насекомого ту или иную железу и приготовить ее экстракт в эфире, хлороформе, гексане или ином органическом растворителе. Такой экстракт можно затем нанести на любой предмет или испарить в определенном объеме воздуха и выяснить реакцию насекомых на содержащийся в нем феромон. На помощь исследователям химической сигнализации пришли и тончайшие методы современной физической химии — газовая хроматография и массовая спектрометрия. Они позволяют не только определить химическую природу феромонов, но и разделять сложные смеси веществ, чтобы изучить роль их компонентов.
Феромоны принято классифицировать на две группы. К первой относятся феромоны-релизеры. Они воздействуют на насекомое через органы обоняния, вызывая немедленный ответ центральной нервной системы и изменения в поведении насекомого. Большинство известных феромонов принадлежит именно к этой группе.
Составляющие вторую группу феромоны-праймеры не вызывают немедленной реакции. В некоторых случаях насекомые воспринимают их органами обоняния, в других праймеры попадают в организм с пищей и воздействуют на эндокринную систему. Это обусловливает физиологические изменения в организме, что может отражаться и на поведении насекомых. Таким образом, феромоны-праймеры вызывают отсроченные реакции. К этой группе относятся, например, феромоны царской пары термитов, регулирующие процессы развития насекомых разных каст, царские феромоны общественных перепончатокрылых, подавляющие плодовитость рабочих и препятствующие появлению новых плодовитых самок. Интересно, что феромоны цариц служат одновременно и релизерами, поскольку вызывают помимо отсроченных эффектов и непосредственные поведенческие реакции — стимулируют фуражировочную и строительную активность рабочих, влияют на их поведение по отношению к личинкам и т. д.
По характеру действия на поведение насекомых феромоны чрезвычайно разнообразны. Они участвуют практически во всех процессах общения и передают всевозможные сигналы. В дальнейшем мы познакомимся с основными классами феромонов. Но несомненно, что первичной функцией феромонов в эволюции было привлечение. Недаром у необщественных насекомых фактически единственными являются так называемые половые феромоны, способствующие встречам самцов и самок. Феромоны, обеспечивающие привлечение, получили название аттрактантов.
Привлечение — это, несомненно, фундаментальная форма коммуникации у общественных насекомых, поскольку социальный образ жизни немыслим без стремления находиться вместе, создавать скопления. Поместим небольшое количество муравьев или медоносных пчел в какой-нибудь пустой закрытый сосуд. Сначала встревоженные насекомые беспорядочно рассеиваются в нем, блуждая туда-сюда. Но постепенно они успокаиваются и начинают собираться в тесные кучки. С течением времени они, скорее всего, образуют одно скопление на дне сосуда. Какие стимулы вызывают подобное поведение? Чаще всего это именно феромоны-аттрактанты. Привлечение участвует в качестве компонента и в других, более сложных системах коммуникации — мобилизации, сигнализации об опасности.
На поверхности тела общественных насекомых всегда есть феромоны-аттрактанты. Достаточно подробные данные о их химической природе и выделяющих их железах получены пока лишь для немногих видов. Так, в задней кишке желтошеего термита при переваривании древесины образуется несложное органическое вещество З-гексен-1-ол. Оно распространяется по всей поверхности тела и в значительной степени обеспечивает взаимную привлекательность термитов. Наибольшей привлекательностью обладают личинки, наименьшей — солдаты. Если термитов кормить чистой целлюлозой или лишить кишечных симбионтов, то привлекающий запах постепенно исчезает. Вероятно, аттрактант синтезируется в результате деятельности жгутиконосцев из каких-то компонентов древесины, отсутствующих в чистой целлюлозе.
У большинства муравьев специальный аттрактант выделяет железа Дюфура. Феромон железы Дюфура муравьи используют для привлечения других рабочих при мобилизации и в некоторых иных ситуациях.
Между пятым и шестым сегментами на верхней стороне брюшка медоносных пчел расположена железа Насонова, выделяющая аттрактант с очень приятным запахом. Он представляет собой смесь, основными компонентами которой являются гераниол, гераниевая кислота и цитраль. Пчелы применяют этот феромон в самых различных ситуациях. Стоящие у входа в улей пчелы-вентилировщицы, как правило, выпячивают железу Насонова. Запах ее феромона насыщает струю воздуха, вытекающую из летка, и помогает отыскивать улей возвращающимся из полетов рабочим. При роении пчелы концентрируются вокруг матки, привлекаемые запахом ее феромонов. Но этот запах они способны воспринимать только на близком расстоянии. Вот почему пчелы, окружающие матку, выделяют аттрактант из железы Насонова, побуждающий всех остальных пчел собраться вокруг матки плотным роем. Феромон железы Насонова пчелы используют и для мобилизации.
На нижней стороне лапок ос-веспин и медоносных пчел обнаружены небольшие железистые образования — так называемые железы Арнхардта. Природа выделяемого ими феромона неизвестна, но установлено, что поверхность, по которой передвигались пчелы и осы, вызывает пристальное внимание других особей того же вида. Поэтому вход в гнездо пчелы и осы отыскивают по знакомому запаху. Когда несколько пчел побывает на кормушке с не имеющим запаха сахарным сиропом, то она становится привлекательной для других пчел, поскольку их предшественники оставили на ней пахучие следы своих лапок.
Аттрактанты содержатся в так называемой линочной жидкости, выделяющейся между сбрасываемой шкуркой и телом термитов. Именно эти вещества привлекают грумов и заставляют их облизывать линяющих насекомых.
В некоторых случаях эффект привлечения могут обеспечивать не специальные феромоны, а иные вещества. Например, для многих муравьев очень притягателен углекислый газ. Если пустить в лабораторное гнездо рыжей мирмики слабую струйку углекислого газа, то муравьи сразу же устремятся к точке, откуда она исходит. В экспериментах Э. Вильсона с огненным муравьем было установлено, что рабочие этого вида отлично умеют ориентироваться по градиенту концентрации углекислого газа, двигаясь обычно в направлении ее увеличения. Это позволяет им присоединяться к группам муравьев, от которых во все стороны распространяется выделяющийся при дыхании углекислый газ. Если несколько муравьев засыпать тонким слоем песка, то другие рабочие начинают выкапывать пленников. Установлено, что землекопы ориентируются по запаху углекислого газа, просачивающегося сквозь песок от зарытых муравьев. Запах углекислого газа чувствуют медоносные пчелы и, вероятно, многие другие насекомые.
Феромоны, выделяемые царицей и царем, прежде всего обеспечивают репродуктивный контроль, о чем уже говорилось. Но одновременно они являются аттрактантами для рабочих. Синтетическая кетодеценовая кислота — феромон царицы медоносных пчел — позволяет создать муляж, заменяющий матку. Если смочить раствором этого вещества любой небольшой предмет в улье, то пчелы образуют вокруг него свиту и будут вести себя совершенно так же, как с настоящей царицей. Легко удается этот опыт и с синтезированным химиками феромоном царицы восточного шершня. Можно получить активные экстракты царских феромонов муравьев, термитов и безжалых пчел. Достаточно положить физогастричную царицу черного садового муравья, например, в пузырек с эфиром, чтобы через несколько дней проделать эффектный опыт: любой пористый предмет (сердцевина веточки бузины, губка и т. п.), смоченный этим экстрактом, будет долго «заставлять» муравьев обращаться с ним, как с царицей. Если царицу муравья-кочевника рода эцитон подержать некоторое время на кусочке дерева или картона, то он станет крайне привлекательным для рабочих муравьев. Такой опыт удается и с другими муравьями, в частности с кампонотусами. Вот насколько сильным аттрактантом является феромон царицы!
Царские феромоны способны очень долго сохраняться. В гнезде огненного муравья труп царицы не утратил привлекательности для рабочих девять недель. А рабочие муравья-портного продолжали «ухаживать» за высохшим трупом царицы полгода. Но рекорд здесь принадлежит, видимо, медоносной пчеле: сухая матка, свыше десяти лет находившаяся на булавке в коллекционном ящике, оказалась все еще в некоторой степени привлекательной для пчел!
У царицы муравьев-портных на верхней стороне брюшка Э. Вильсон и Б. Хеллдоблер обнаружили железы, открывающиеся отверстиями на небольших выростах перепонки между сегментами. Вот эти выросты и привлекают особенно рабочих, облизывающих их и все тело царицы. У огненного муравья царица, видимо, выделяет аттрактант из ядовитой железы. На верхней стороне брюшка находятся продуцирующие аттрактант железы у царицы безжалой пчелы паратригона субнуда. Царица очень часто вибрирует крыльями и потирает ножками брюшко, этим она способствует распространению по телу феромона.
Неодинаковое количество выделяемого аттрактанта может лежать в основе предпочтения рабочими одной из нескольких цариц. Эксперименты с огненным муравьем позволили установить, что царицы значительно различаются по привлекательности, а рабочие чаще всего предпочитают царицу, выделяющую больше феромонов, нередко убивая остальных. Как показывают некоторые данные, количество выделяемого царицей аттрактанта напрямую связано с ее плодовитостью. Поэтому выбор рабочими самой привлекательной царицы вполне оправдан. Различия в привлекательности цариц определяют очень интересный процесс смены царицы, в деталях прослеженный у вида паратригона субнуда. У этой безжалой пчелы вышедшие из куколок плодовитые самки прячутся в районе горшков с провизией и даже занимаются там разнообразной строительной деятельностью. Постепенно молодая «претендентка на престол» становится все более привлекательной для пчел. Она выделяет изо рта капельки секрета мандибулярных желез, которые поглощают рабочие. Пчелы поедают и экскременты царицы. Вероятно, в мандибулярном секрете и экскрементах тоже содержатся феромоны, но это пока не исследовано. Брюшко молодой царицы увеличивается, и вокруг нее появляется обширная свита пчел, которые облизывают царицу, особенно кончик ее брюшка, и вступают с ней в трофаллаксис — в процессе его царица издает характерный запах, хорошо ощутимый даже человеком. В это время молодая царица либо покидает гнездо и с роем пчел переселяется в новое, либо пытается сместить старую царицу.
В последнем случае молодая царица в сопровождении свиты приближается к старой и, возбужденно вибрируя крыльями, наносит на ее брюшко капельку мандибулярного секрета, а иногда еще и капельку из анального отверстия. Старая царица начинает отчищаться, ее облизывают и окружающие пчелы. Если старая царица еще плодовита, она, видимо, выделяет достаточно феромонов и сохраняет привлекательность для рабочих. В этом случае пчелы не позволяют молодой царице «обижать» старую, прогоняют и даже убивают ее, если она не может покинуть гнездо с роем. Но если феромоны у молодой царицы оказываются «сильнее», чем у старой, то она еще и еще раз пачкает старую царицу своим секретом. В конце концов это вызывает у пчел враждебность к старой царице, они нападают на нее и раздирают верхнюю сторону брюшка, где находятся кожные железы. Молодая царица становится единственной в гнезде. «Дворцовый переворот» завершился!
Есть у общественных насекомых и половые феромоны-аттрактанты. Трутни находят в воздухе летящую матку по сохраняющемуся запаху ее кетодеценовой кислоты, которая, таким образом, выполняет функции и полового аттрактанта. Самки и самцы термитов оставляют за собой пахучие следы с помощью той же железы, что выделяет следовой феромон у рабочих термитов (подробнее см. в подразделе «Мобилизация»). Самцы шмелей производят половой аттрактант в мандибулярных железах. Крылатые самки некоторых муравьев, в особенности эргатоидные царицы ряда видов (харпагоксенусы и др.), привлекают самцов, принимая призывную позу с поднятым кверху брюшком и выделяя аттрактант. Самцы муравьев-древоточцев, вылетая из гнезда, выделяют какой-то феромон, побуждающий самок присоединиться к их полету.
В очень редких случаях общественные насекомые используют для привлечения себе подобных не феромоны, а звуковые сигналы. Еще Э. Рубо установил, что «вибрация крыльев осы-няньки служит сигналом для личинки, которая для того чтобы получить корм, высовывает голову из ячейки... Достаточно бывает издать громкий свист или другой резкий звук возле гнезда белоногастера — и головы всех личинок показываются из отверстий».[12] Голодные личинки ос-веспин, двигая головой, скребут челюстями по стенкам ячеек, издавая характерный «шуршащий» звук. Личинка подает сигнал каждые несколько секунд. Осы воспринимают возникающую при этом вибрацию сота, быстро находят голодных личинок и кормят их. Интересно, что если таких личинок в гнезде несколько, то они скребут челюстями синхронно. Быть может, призывные сигналы в «хоровом исполнении» быстрее доходят до рабочих ос?
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Потребность общения
Потребность общения Два миллиона языковВ старинной немецкой балладе о крысолове из Гамельна рассказывается, как местный бродячий волшебник, чтобы спасти город от расплодившихся крыс,«На дивной дудке марш сыграл И прямо в Везер крыс согнал».Здесь, в волнах реки Везер
Способы расселения.
Способы расселения. Этот вопрос уже разбирался сэром Ч. Лайеллем и другими авторами. Я, со своей стороны, могу представить здесь очень краткий обзор наиболее важных фактов. Перемены в климате должны были оказать сильнейшее влияние на миграцию. Регион, в настоящее время
Жажда общения
Жажда общения Нужен ли оратору слушатель? Вспомните, как мы, разговаривая по телефону, сопровождаем нашу речь мимикой и жестами. Наблюдая со стороны за погруженным в свои мысли человеком, по его мимике можно проследить за ними. Мимика и жесты – часть (и немаловажная)
Проблемы общения
Проблемы общения В концепции Л.В. Крушинского о рассудочной деятельности интересно представление о двух его механизмах, особенно о вербальном. Автор допускает существование вербального мышления у животных, в частности у человекообразных обезьян, и возможность
Способы вызывания гипноза
Способы вызывания гипноза Обращаясь к вопросу о вызывании гипноза, необходимо иметь в виду, что для этого с пользою могут служить различные способы, из которых одни могут считаться физиологическими, так как они действуют непосредственно на те или другие из
Азбука общения живых организмов
Азбука общения живых организмов Многообразен мир животных, и каждый существующий вид в своей жизни пользуется довольно сложной и своеобразной сигнализацией. Среди различных способов передачи информации в животном мире значительная роль отводится запахам. Животные и
СПОСОБЫ ДРЕССИРОВКИ
СПОСОБЫ ДРЕССИРОВКИ Под способом понимают такое воздействие на животное, которое приводит к воспроизведению (появлению) нужных дрессировщику действий или сохранению необходимых поз.Хотя, по словам К. Прайор, «способов дрессировки существует столько, сколько
Способы размножения
Способы размножения Размножение — важнейший биологический процесс, обеспечивающий поддержание и увеличение численности вида, возможность его расселения и, в конечном итоге, успех борьбы за существование. В животном мире существует целый ряд способов размножения,
Разные способы защиты
Разные способы защиты Способы защиты насекомых от врагов бесконечны в своем многообразии. Тот, кто часто бывает на природе, мот заметить, что как только какое-либо животное становится многочисленным — будь то ящерица, птица, зверь, насекомое, оно сразу же делается менее
Пренатальное развитие сенсорных способностей и элементов общения
Пренатальное развитие сенсорных способностей и элементов общения Влияние сенсорной стимуляции на двигательную активность эмбриона Выше приводились примеры рефлекторных движений зародышей, производимых преимущественно в ответ на тактильные раздражения.
Раннее формирование общения
Раннее формирование общения Раннее формирование общения у птиц Ознакомившись в общих чертах с основными компонентами раннего постнатального развития поведения и их взаимоотношениями, рассмотрим теперь специально, как формируется общение между детенышами и
Раннее формирование общения у птиц
Раннее формирование общения у птиц Ознакомившись в общих чертах с основными компонентами раннего постнатального развития поведения и их взаимоотношениями, рассмотрим теперь специально, как формируется общение между детенышами и родителями и другими особями. Правда,
Раннее формирование общения млекопитающих
Раннее формирование общения млекопитающих У млекопитающих взаимное запечатление индивидуальных отличительных признаков родителей и детенышей и установление контактов между ними происходят в разные сроки после появления детеныша на свет — в зависимости от степени
Формирование общения у низших позвоночных
Формирование общения у низших позвоночных Отмеченные закономерности не имеют конечно, значения всеобщих правил для всех позвоночных, а тем более беспозвоночных. Даже среди птиц и млекопитающих обнаруживаются многочисленные отклонения и исключения, обусловленные
Формирование общения в играх животных
Формирование общения в играх животных Совместные игры Групповое поведение у высших животных также формируется в большой степени в процессе игры. Эту роль выполняют совместные игры: под ними следует понимать такие игры, при которых имеют место согласованные действия