Необычная сохранность
Ископаемые отпечатки — это почти единственные свидетельства существования мягкотелых представителей биоты или мягких частей организмов. К счастью, только «почти». Сохранение беспозвоночных в некоторых редких местонахождениях окаменелостей заходит гораздо дальше обычного сохранения экзоскелета или раковины: мельчайшие детали целого тела, включая глаза и придатки, сохраняются в ископаемом состоянии на различных стадиях развития. Их обнаружение и изучение сложны, поскольку обычно они хрупкие и мелкие. Но благодаря новым методам получения изображений и компьютерным программам трёхмерной реконструкции открывается много новой информации. Местонахождения имеются по всему миру и среди них формация Крато из меловых отложений Бразилии, Золенхофен из юры Баварии, райниевые черты из девона Шотландии, и «орстенские» местонахождения по всему миру, лучше всего известные из Швеции и охватывающие период от нижнего кембрия до нижнего ордовика.
Открытия и описания онтогенетических стадий столь большого количества таксонов быстро наполняют литературу. В дальнейшем накопление этой информации, синтез и анализ этих открытий, несомненно, окажут огромное влияние на наше понимание самых ранних стадий развития нескольких групп членистоногих. Некоторые международные исследовательские группы, такие, как группы Марка Саттона и Дерека Бриггса, используют сложное программное обеспечение для получения трёхмерных реконструкций в целях изучения крохотных и хрупких окаменелостей, заключающих в себе богатую морфологическую информацию.
Пример, который я упоминаю здесь, касается изучения раннего представителя ракообразных из среднего кембрия Швеции, Henningsmoenicaris scutula.
Йоахим Хауг и его коллеги идентифицировали десять онтогенетических стадий этого животного и создали, как они любят это называть, «четырёхмерную модель», где четвёртым «измерением» является время. Эти стадии документируют время и способ добавления сегментов и их придатков. Первая стадия имеет четыре сегмента, несущих придатки; это количество увеличивается до семи, после чего происходит дифференциация отделов тела.
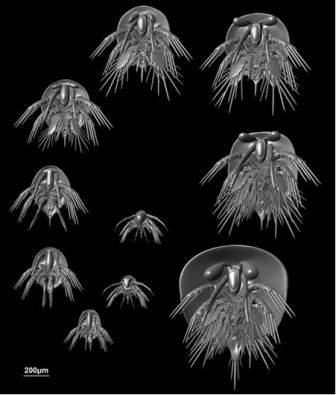
Рисунок 51. 4-D модель Henningsmoenicaris scutula, примитивного ракообразного из кембрия Швеции. Рисунок любезно предоставил Йоахим Хауг (Нью-Хейвен).
Хауг и его коллеги документировали развитие сложных глаз, впервые распознаваемых на пятой стадии из десяти имеющихся в наличии. Каждая из последующих стадий документирует изменение от сидячих глаз к стебельчатым. Прекрасная морфология даёт нам не только эстетически приятное описание изменений формы, происходивших около 520 миллионов лет назад, но также данные, важные для понимания ранних этапов разделения ракообразных и других групп членистоногих в процессе эволюции.
Какие способы развития характеризовали происхождение крупных групп членистоногих, в том числе трилобитов? Являются ли общие системы индивидуального развития признаком эволюционного родства, или они эволюционировали независимо в различных группах? Хауг и его коллеги заметили, что способ слияния последних туловищных сегментов у Henningsmoenicaris в ходе онтогенеза также присутствует у трилобитов: новые сегменты появляются и присоединяются к прилегающему к ним спереди сегменту, в противоположность картине, имеющей место у более поздних ракообразных. Эти и другие аспекты относительного времени возникновения онтогенетических событий можно нанести на эволюционные древа, чтобы отследить, насколько консервативным или пластичным было индивидуальное развитие на самых ранних стадиях эволюции многоклеточных животных.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК