О Saurichthys и рыбах-мутантах
Я обсуждал главным образом то, как палеонтологические данные могут дополнить наше понимание эволюции истории жизни и механизмов индивидуального развития. Но могут данные из области эволюции индивидуального развития сообщить что-либо палеонтологам? Могут ли открытия из области молекулярной эволюционной биологии развития внести свой вклад в понимание разнообразия вымерших таксонов? Поскольку нуклеиновые кислоты и белки очень редко сохраняются с течением геологического времени, реконструкция молекулярной основы морфологических изменений у ископаемых форм должна основываться на умозаключениях.
Здесь я исследую основания для таких выводов, рассматривая их на примере ископаемой рыбы.
Среди богатых коллекций среднетриасовых позвоночных в Цюрихе есть замечательная рыба. Внешне она выглядит похожей на современную щуку, с удлинённой и обтекаемой формой тела, которая выдаёт в ней хищника. Её название — Saurichthys, «рыба-ящерица». Этот род располагается довольно близко к основанию родословного древа костных рыб, близко к осетрам, веслоносам и ильной рыбе, своим ныне живущим родственникам. Существуют четыре вида, известных из местонахождения в Монте-Сан-Джорджио в Швейцарии, и они выставлены в Цюрихе, но род имеет всесветное распространение и включает более тридцати видов. Те виды, которые имеются в нашем музее, наиболее очевидно характеризуются различным рисунком и количеством чешуй, как было установлено в 1980-х годах Оливье Риппелом, когда он работал в Цюрихе. Гипотетический последний общий предок, давший начало радиации видов Saurichthys, может быть уверенно реконструирован на основании нескольких других окаменелостей как полностью покрытый однородной ромбовидной чешуёй, с многочисленными, сильно сегментированными плавниковыми лучами и с большим количеством дермальных костей в черепе. Виды рода Saurichthys, происхождение которых можно отследить назад к этому гипотетическому предку, характеризуются различной степенью редукции чешуйного покрова (ряды и вытянутая форма чешуй), редукцией количества плавниковых лучей и их сегментов, сокращением количества дермальных костей в черепе и их формой, отражающейся в форме жаберной крышки, а также снижением степени массивности зубов и челюстей. Фактически, новизна в эволюции во многих случаях ассоциируется с утратой структур. Хорошие примеры этого явления — общая редукция экзоскелета во многих родословных линиях и утрата тяжёлой брони у силурийских и девонских рыб.[80] Но нам пока нет необходимости возвращаться во времени так далеко. Утрата или редукция чешуй у других рыб не представляет собой ничего необычного. Например, у осетров и веслоносов (отряд Осетрообразные) полный чешуйный покров, зарегистрированный у образцов из позднего мела, редуцировался до пяти рядов щитков у ныне живущих видов{7}. Утрата чешуй независимо происходила в процессе эволюции, по крайней мере, тринадцать раз у гольянов и карпов (карпообразные) и несколько раз у галаксиевых, саланксовых (корюшкообразные) и икромечущих карпозубых (карпозубообразные). В пределах некоторых видов возможна значительная изменчивость. Одомашненный карп Cyprinus carpio существует в виде различных разновидностей: кожистый карп с фактически отсутствующей чешуёй; зеркальный карп с немногочисленными увеличенными чешуями и зеркальный линейный карп с единственным рядом крупных чешуй, тянущимся вдоль боков. Эти примеры изменчивости и эволюционных изменений вызвали интерес к расшифровке генетических механизмов, лежащих в основе явления редукции чешуи.[81]
Известны сотни образцов Saurichthys, и изучение некоторых из них привело к открытию живорождения у этого животного. Я всегда думал, что когда-нибудь я бы хотел изучить эту рыбу и поработать с ней, но это случилось довольно неожиданным образом. Генетика развития и любезный и вежливый отставной доктор медицины из Базеля Лиени Шмид являются ключевыми фигурами в этой истории.
В последние годы Лиени Шмид присоединился к Рудольфу Стокару и его коллегам в Лугано, собирая Saurichthys в триасовых породах в южной Швейцарии. Он также посещал курсы эво-дево, на которых обсуждались самые новые работы, связывающие морфологическую эволюцию с генетикой развития. Среди них были исследования некоторых видов рыб, которые хорошо известны с точки зрения генетики и служат моделями для понимания эволюционных изменений. Шмид заметил сходство в характере морфологических изменений, представленных в исследованиях генетики развития видов рыб, с изменчивостью у Saurichthys.
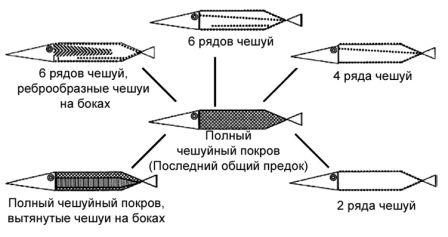
Рисунок 35. Схемы, показывающие разнообразие картины чешуйного покрова у Saurichthys. Последний общий предок должен представлять собой рыбу, полностью покрытую однородной ромбической чешуёй. По Schmid и S?nchez-Villagra 2010, с изменениями.
Он рассказал мне о своих идеях, и мы исследовали вопрос глубже и написали об этом работу, которую я вкратце излагаю здесь.
Серия изящных исследований, которые включены в учебники, иллюстрирует генетические и фенотипические изменения на популяционном уровне в экологическом контексте. Лучше всего известный пример — это поразительная изменчивость в строении чешуйчатого покрова у колюшек.[82] У современных видов был зарегистрирован быстрый рост морфологического разнообразия, порождённый изменением регуляторных функций и взаимодействием между генами, экспрессия которых происходила посредством изменяющихся экологических условий. Трёхиглая колюшка (Gasterosteus aculeatus) несёт на своих боках ряд из примерно 36 пластинок — видоизменённых чешуй. В пресной воде последовательно отбиралась мутация эктодисплазинового сигнального пути (совокупности взаимодействий на молекулярном уровне), которая уменьшала количество этих видоизменённых чешуй с 36 до 0–7 в гомозиготном состоянии и до промежуточного количества в гетерозиготном. Молекулярные генетические факторы, ведущие к утрате чешуйчатого покрова, были изучены в лаборатории и в полевых условиях на колюшках, а также на мутантах нескольких видов рыб.
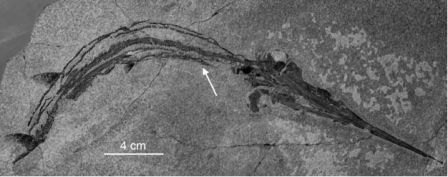
Рисунок 36. Образец Saurichthys curionii из триаса Монте-Сан-Джорджио, Швейцария (Палеонтологический институт и музей, Цюрихский Университет).
Существуют другие мутанты по эктодисплазиновому сигнальному пути. На примере японской медаки (Oryzias latipes) было продемонстрировано, что мутант с утратой чешуи возник из-за мутации в рецепторе этого пути. У данио-рерио (Danio rerio) было идентифицировано несколько мутаций, как в сигнальной молекуле самого эктодисплазина, так и в рецепторе, которые приводили к появлению фенотипов со сходными типами утраты чешуи. Другой молекулярный механизм, приводящий к утрате чешуи, был обнаружен у мутанта данио-рерио и у домашних карпов. Причиной этого является мутация утраты функционирования в рецепторе-1 фактора роста фибробластов. Известна важность факторов роста фибробластов на всех стадиях развития. Жизнеспособный мутант с мутацией этого пути возможен только благодаря дублированию гена, поскольку летальные побочные эффекты мутации в одном гене компенсируются немутированной версией этого же гена, которая в терминологии генетиков носит название «паралог».
Мы с Шмидом постулировали, что подобные генетические процессы лежат в основе фенотипических изменений у Saurichthys и у мутантов этих рыб. Молекулярные основы морфологического разнообразия Saurichthys включали мутацию или регуляторные изменения сигнального пути в качестве ключевого механизма. Существуют серьёзные основания полагать, что эти механизмы действовали у триасовых рыб так же, как в наши дни.
Анализ последовательности генов, кодирующих компоненты эктодисплазинового сигнального пути, показывает, что эти гены у позвоночных очень консервативны. Известно, что эктодисплазин контролирует формирование зубов и дермальных костей у челюстноротых позвоночных, чешуи у рыб, перьев у птиц и волос и кожных желёз у млекопитающих. Что же касается факторов роста фибробластов, то их работа существенна на очень ранних стадиях развития и они явно унаследованы по меньшей мере всеми позвоночными животными. Эти сигнальные пути представляют собой пример того, что названо «инструментальными белками»{8} и хорошо сохраняется в ходе эволюции.
В дополнение к редукции количества чешуй, вышеупомянутые виды рыб благодаря мутации в эктодисплазиновом пути или в пути фактора роста фибробластов демонстрируют тенденцию к выстраиванию оставшихся чешуй в виде рисунка, схожего с тем, который наблюдается у отдельных рядов чешуй у Saurichthys. Кроме того, сохраняющиеся чешуи на боках мутантов вытянуты в дорсовентральном направлении так, что их можно сравнить с удлинёнными боковыми чешуями, как у видов Saurichthys с реброобразными боковыми чешуями. Мутации в компонентах эктодисплазинового пути также ведут, по крайней мере, у некоторых мутантов данио-рерио, к сокращению количества и снижению сложности строения плавниковых лучей, как было отмечено для некоторых морфологических типов Saurichthys. Кроме того, были сообщения об утрате зубов и изменениях размеров дермальных костей у мутантов данио-рерио, опять-таки изменения органов, которые менялись в процессе роста видового многообразия у Saurichthys.

Рисунок 37. Схематичные изображения рыб дикого типа и мутантов различных модельных видов, используемых в исследованиях в области генетики развития. Эктодисплазиновый путь: данио-рерио и японская медака, по Harris и др. 2008; трёхиглая колюшка, по Colosimo и др. 2005. Путь фактора роста фибробластов, по Rohner и др. 2009. Рисунки сделаны Л. Шмидом (Базель), из Schmid и S?nchez-Villagra 2010, с изменениями.
Особенности строения, контролируемые инструментальными белками, в том числе чешуйчатый покров, строение плавниковых лучей и особенности зубной системы, являются особенно важными для целей самозащиты, хищничества и питания, и таким образом для адаптации в целом. Изменение в важном гене, определяющем ход развития, даёт хороший шанс для симпатрического видообразования. Чем больше будет фенотипический эффект единственной мутации, тем больше шанс немедленной репродуктивной изоляции мутантов, если они окажутся жизнеспособными и успешными. Изменения в сигнальных путях, наличие которых мы предполагаем в случае увеличения видового многообразия у Saurichthys, могут, таким образом, дать объяснение явлению очень быстрого видообразования. В противоположность градуалистической модели эволюции, изменение в важном гене, контролирующем развитие, представляет собой реалистичное объяснение существенных различий между видами рода Saurichthys в палеоэкологическом контексте, в котором они возникли. Тот факт, что не были обнаружены промежуточные, плавно отличающиеся друг от друга ископаемые формы Saurichthys и большинства других видов, хорошо вписывается в общую картину.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК