Глава девятая Развитие у млекопитающих и у человека
Существует около 5300 видов современных млекопитающих. Они представляют собой лишь небольшую часть от общего количества видов, когда-либо существовавших начиная с обособления ведущей к ним эволюционной линии как минимум 315 миллионов лет назад. Затем родословные ветви рептилий и млекопитающих разделились. По оценкам, сделанным на основе ископаемых остатков, прошло ещё около 100 миллионов лет, когда в юрский период появился последний общий предок всех ныне живущих млекопитающих.[88] Хотя цифра в 5300 современных видов выглядит внушительно, она не столь уж внушительна по сравнению с другими группами: существует вдвое больше видов птиц и в пять раз больше видов костистых рыб. Но млекопитающие выглядят весьма особенными, когда мы рассматриваем их значительное экоморфологическое разнообразие — способы локомоции у людей, кенгуру, китов и летучих мышей; размер, варьирующий от землеройки до синего кита. Что у всех млекопитающих есть общего — это уникальные для позвоночных история жизни и физиология, в том числе энергозатратная эндотермия, родительская забота, выделение молока и детерминированный рост. Эти и многие другие особенности должны были характеризовать последнего общего предка ныне живущих млекопитающих. Когда же, начиная с момента отделения от рептилий, возникли эти особенности? Появились ли они одновременно? Если нет, то в какой последовательности? Неважно, как обстояли дела у наших предков, но во время двух критических отрезков геологической истории их выручали удача и хорошие гены. Я уже говорил выше, как Lystrosaurus и его родственники преодолели границу перми и триаса, время, когда множество групп организмов было полностью стёрто с лица земли. Млекопитающие также пережили событие конца мелового периода, когда вымерло большинство{13} динозавров.
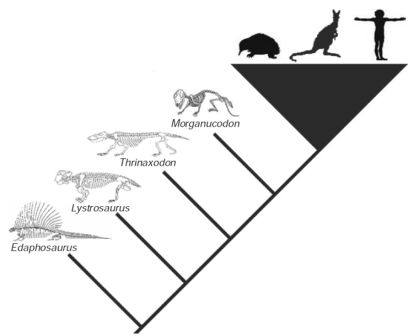
Рисунок 43. После дивергенции с рептилиями не позднее 315 миллионов лет назад началась долгая история «предковых млекопитающих» — вплоть до появления по крайней мере 220 миллионов лет назад последнего общего предка современных видов. Ныне живущих млекопитающих представляют ехидна (однопроходное), кенгуру (сумчатое) и человек (плацентарное). Предковые виды, изображённые здесь — это все ключевые виды, упомянутые в тексте, но они — лишь крохотная часть общего многообразия параллельных эволюционных линий, которые развивались на раннем этапе эволюции млекопитающих.
Несколько уникальных анатомических особенностей млекопитающих не связаны непосредственно с особенностями жизненной истории, например, наличие трёх небольших косточек уха в противоположность только одной у рептилий и птиц. Способы, которыми эмбриологические и палеонтологические данные документируют происхождение слуховых косточек млекопитающих, прекрасно соотносятся друг с другом, и их открытие является одним из величайших триумфов сравнительной анатомии. Как я уже рассказывал в восьмой главе, Гольдшмидт сделал упор на отсутствие промежуточных форм в эволюции и важность эпигенетики, и, в дополнение к глазам камбалы, среднее ухо млекопитающих было его основным примером. Оба они оказались плохими примерами в остальном хорошей идеи. Фактически, Гольдшмидт должен бы приветствовать новейшее свидетельство того, что происхождение среднего уха млекопитающих было связано в процессе развития с увеличением размера мозга и, вероятнее всего, со многими другими особенностями, в том числе с изменениями в жевательном аппарате. Сопутствующие друг другу изменения в процессе развития показывают, что эти особенности не могут быть поняты отдельно друг от друга. В ходе онтогенеза современных млекопитающих рост неокортекса связан с постепенным отделением элементов, связанных с челюстями. Некоторые из старых элементов челюсти становятся относительно меньше и меньше по мере того, как они обособляются и начинают формировать будущие элементы среднего уха — это пример отрицательного аллометрического роста. Постепенные анатомические изменения, сходные с изменениями у эмбрионов — это то, что ископаемые остатки документируют в основной линии, ведущей к современным млекопитающим. Эти изменения зафиксировал Тим Роу из Техасского Университета на основании изучения полученных методом компьютерной томографии изображений окаменелостей и реконструкции развития эмбрионов сумчатых млекопитающих современных видов.[89]
Наличие взаимосвязи, или отсутствие независимости в ходе эволюционного возникновения млекопитающих также характеризует особенности истории жизни. Документация этих эволюционных изменений ограничена скелетом, поскольку мягкие ткани, включая органы вроде сердца или желудка сохраняются редко, если сохраняются вообще. Не удивительно, что большинство свидетельств получено при изучении зубов и гистологии костей.
Сложная картина зубной окклюзии у млекопитающих, когда верхние и нижние коренные зубы, обладающие несколькими буграми, смыкаются друг с другом особым образом, несомненно, коррелирует с высокой скоростью обмена веществ, который, в свою очередь, связан с особенностями истории жизни. Когда большинство рептилий, рацион которых включает других животных, просто ловит свою добычу, и, самое большее, режет её зубами перед тем, как проглотить, млекопитающие имеют тенденцию жевать то, что едят. Выделения слюнных желёз смешиваются с пищей, таким образом частично обрабатывая её во рту перед тем, как она будет переварена в желудке и кишечнике.
Развитие молочного вскармливания у млекопитающих связано с особенностями зубов, такими, как позднее прорезание первых функциональных зубов. У млекопитающих образуется только две смены зубов — это состояние, называемое «дифиодонтия» и включающее наборы зубов молодой и взрослой особи. Кормление молоком, вероятнее всего, возникло, когда характер смены зубов стал дифиодонтным.[90] С этим, в свою очередь, был связан детерминированный рост — это означает, что быстрое развитие ювенильной особи завершается по достижении взрослого размера. У рептилий, напротив, рост главным образом недетерминированный, и это означает, что крокодил, например, никогда не останавливается в росте и у него никогда не прекращается замена зубов, хотя скорость роста может замедлиться с возрастом.
Каждый из существующих в наше время видов млекопитающих имеет определённую зубную формулу, означающую определённое число резцов, клыков, предкоренных и коренных зубов. Эта дифференциация зубной системы специфична для млекопитающих, как и тот факт, что самая задняя группа зубов, коренные зубы, не сменяется. Ископаемые остатки документируют момент, когда установились эта дифференциация зубов и характер их смены.
Серия образцов, демонстрирующих процесс роста триасового «стволового» млекопитающего Thrinaxodon{14}, показывает, что у этого животного было восемь заклыковых зубов на одной из ювенильных стадий, и ещё четыре добавлялось сзади по мере продолжения роста. Sinoconodon, который жил в позднем триасе и считается, как и Morganucodon, одним из «первых млекопитающих», явно обладал недетерминированным ростом и многократно сменявшимися резцами. У мелового млекопитающего Gobiconodon «коренные зубы» сменялись.
Благодаря комбинированию данных о характере прорезывания зубов и палеогистологических данных о росте становится возможным проследить происхождение особенностей истории жизни, характерных для млекопитающих. В палеогистологических исследованиях важным моментом является выявление наличия фиброламеллярной кости, поскольку она указывает на быстрый остеогенез и вместе с тем быстрый общий рост, связанный также с эндотермией (см. главу 4). Этот вид кости обнаружен у многих, хотя и не у самых ранних члены родословной ветви млекопитающих. Самые ранние предки млекопитающих по физиологии и по особенностям индивидуального развития были больше похожи на современных рептилий, чем на млекопитающих. Пример такого рода — Dimetrodon, который обладал недетерминированым ростом, характерным для ранних амниот, и особенности его гистологии показывают отсутствие фиброламеллярной кости.
Помимо прямого указания со стороны гистологии костей, существует косвенный способ, позволяющий выявить детерминированный рост — путём популяционных исследований ископаемых форм при возможности хорошего стратиграфического контроля. Он требует большого количества окаменелостей одного и того же вида из одного и того же слоя отложений. Если в выборке все взрослые особи обладают одинаковым размером, это дало бы основание предполагать наличие детерминированного роста. Как раз это отмечалось по итогам популяционного исследования Morganucodon из позднего триаса — ранней юры, которого многие палеонтологи рассматривают как одно из «первых млекопитающих» из-за набора анатомических особенностей, присутствующих у этого животного. У млекопитающих размер — это в целом хорошее внешнее проявление возраста неполовозрелых особей, поскольку факторы окружающей среды вроде температуры оказывают не настолько сильное воздействие на их размер, как это бывает у рептилий. В отличие от Morganucodon, у Dimetrodon, а также у его близкого родственника Ophiacodon, взрослые животные одного и того же вида имеют различные размеры.[91] У этих самых ранних членов нашей родословной линии, как у рептилий, размер является ненадёжным признаком стадии развития. Поскольку информация об анатомии подтверждает принадлежность особей различных размеров к одному и тому же виду, это наблюдение подразумевает то, что эти самые ранние представители родословной линии млекопитающих не обладали характерным для млекопитающих способом роста и потому для них размер не является хорошим внешним признаком возраста.
На время развития эмбрионов у рыб, амфибий и рептилий оказывает значительное воздействие температура, при которой происходит их инкубация, и у многих видов ею определяется даже пол. В зависимости от температуры, при которой происходила инкубация у черепахи, и которая оказывала значительное влияние на темп обмена веществ и тем самым на рост, размеры на такого рода «стадиях» развития могут быть весьма различными. Млекопитающие, в свою очередь, характеризуются более стабильной внутренней температурной средой, так называемой гомеостатической способностью, достигающейся путём поддержания высокой скорости обмена веществ в постоянной внутренней среде.
Для первых юрских млекопитающих была характерна меньшая пластичность индивидуального развития, с более стабильной картиной роста по сравнению с часто регистрируемыми случаями ускоренного и замедленного роста, типичными для рептилий и самых ранних представителей ветви млекопитающих. Способность приостанавливать или замедлять рост при неблагоприятных условиях окружающей среды должна была характеризовать, например, Dimetrodon, что подтверждается палеогистологическими данными.
Том Кемп из Оксфордского Университета назвал интегрированную эволюцию признаков, описанных выше, «коррелированной прогрессией». Он думает, что эта плавная и почти синергическая картина эволюции, напоминающая сеть взаимоотношений, меняющихся со временем, характеризует не только многое в ранней эволюции млекопитающих, но также происхождение других крупных групп, таких, как черепахи и первые позвоночные с конечностями, или четвероногие. Идея во многом выглядит привлекательной. Конечно, координация изменений в различных частях тела, вероятно, вызвана скорее последовательностью небольших, нарастающих изменений, чем одним единственным шагом. Согласно Кемпу, изменения признаков «эволюционируют аналогично цепочке людей, идущих вперёд, взявшись за руки: любой из них может двигаться в своём собственном темпе, впереди или позади тех, кто находится рядом, но лишь разорвав цепочку». Экспериментальные свидетельства, полученные для таких разных организмов, как лабораторные мыши, растения, насекомые, черепахи и даже люди, служат напоминанием того, что признаки не эволюционируют сами по себе; они являются частью целого организма. Известны эксперименты, в которых исследователи пробовали оценить степень, в которой изменение формы определяется генетической основой. В ходе управляемого размножения или при исследовании информации о чистокровном разведении исследователи изучают эффект селекционных экспериментов, связанных с определённой особенностью. В этих случаях обычно имеет место ответ на отбор со стороны иных аспектов облика сверх того, на который был изначально направлен отбор.
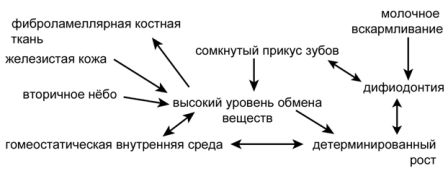
Рисунок 44. Иллюстрация взаимосвязей между несколькими признаками, характерными для млекопитающих, которые, несомненно, эволюционировали параллельно и по принципу «коррелированной прогрессии».
Что касается происхождения картины истории жизни млекопитающего, было бы неправильно предполагать, что первостепенным был какой-то один конкретный меняющийся признак, поскольку они все интегрированы. Изменение в одной переменной величине должно было сопровождаться изменениями в других переменных организма, чтобы они могли функционировать, соответствовать «условиям для поддержания жизни». Рисунок 44 изображает эту идею о сети связей. Картина корреляции и взаимодействий в эволюции, представленная на рисунке, имеет в своей основе эмпирические свидетельства. По мере того, как летопись ранней эволюции родословной линии млекопитающих всё лучше наполняется находками, а исследования истории жизни становятся более определёнными в количественном выражении, у нас появится возможность выверить и дополнить деталями эту картину. Умная идея о коррелированной прогрессии остаётся лишь идеей, если она не подкреплена эмпирическими свидетельствами, а единственный источник прямых свидетельств — это летопись окаменелостей.
История жизни и физиология млекопитающих коррелятивно связаны с несколькими анатомическими особенностями, и некоторые из них зарегистрированы на ископаемых образцах. Они показывают нам, что диагностические особенности млекопитающих не возникали одновременно, а некоторые из них эволюционировали конвергентно в линиях, вымерших в дальнейшем. Пример особенности строения, которая независимо возникала в процессе эволюции несколько раз — вторичное нёбо. Вторичное нёбо — это состоящая из кости и мягкой ткани горизонтальная перегородка, которой мы можем коснуться языком, когда он движется в верхней части ротовой полости. Эта особенность, разделение между ротовой и носовой полостями, позволяет нам, млекопитающим, дышать, пока мы едим, и таким образом способствует более эффективной обработке пищи и выработке энергии для поддержания высокой скорости нашего обмена веществ.[92]
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК