Глава пятая Пропорции, рости таксономия
Во времена, когда было сделано очень мало анатомических исследований микроскопических структур, многие люди думали, что внутри яиц содержатся полностью сформированные очень маленькие индивидуумы, или гомункулюсы — это теория, известная как преформизм (рисунок 23). Для других видов предполагалось существование аналогичных «анималькулюсов». В наше время эта идея кажется нам смешной, но в то время не было никаких микроскопов, а о наличии эволюционных преобразований организмов ещё не догадывались. Но что действительно замечательно и несёт гораздо меньше смысла без знакомства с современным положением дел в биологии, так это то, как яйцеклетка и сперматозоид могут совместно давать информацию и способность выживать и развиваться целому организму. Сейчас мы знаем, что ранний этап онтогенеза сложных организмов включает простое деление клеток снова и снова, и в конечном счёте происходит развитие в организм узнаваемого облика. Открытие этого процесса и механизмов, лежащих в его основе на различных уровнях организации — это великолепное научное достижение.

Рисунок 23. «Гомункулюс», каким его изображал Николас Хартсокер (1694); эмбрион находится в сперматозоиде уже в готовом виде. Рисунок Мадлен Гейгер.
Есть два аспекта онтогенеза, периода от зачатия до смерти, которые рассматриваются отдельно: развитие и рост. Развитие относится к процессам дифференциации клеток и формирования основного плана строения тела, с фундаментальными структурными изменениями и первым появлением основных деталей строения. Оно начинается с зачатия и завершается приблизительно в момент формированием основных тканей тела. Рост является более поздней стадией онтогенеза, в ходе которой увеличивается размер. В основе этой стадии лежит эмбриональный план строения, который уже сформировался в ходе процесса развития. Из-за связи фоссилизации с процессами сохранения многое из того, что палеонтология может сказать об онтогенезе, относится к процессу роста. Косвенно она может сказать намного больше, о чём я расскажу в следующих главах.
Рост как у ископаемых, так и у ныне живущих форм включает изменения размеров и формы. Значительная область исследования связана с выявлением математических законов, которые управляют этими изменениями. Одна из самых ранних попыток сделать это была предпринята многоуважаемым учёным Д’Арси Томпсоном (1860–1948), профессором зоологии в Cент-Эндрюсском Университете в Шотландии. Он был также экспертом в области математики и греческого языка, и именно он, в числе своих прочих заслуг, подготовил стандартный перевод «Истории животных» Аристотеля. Томпсон описал свой подход в книге «О росте и форме», впервые изданной в 1917 году. Он предложил координатные сетки, с помощью которых было возможно перевести сложные различия в формах в простые геометрические понятия. Их предыдущую и более простую версию, касающуюся различий в пропорциях человеческого лица, можно найти в записках Альбрехта Дюрера. Несколько примеров, которые использовал Томпсон в анализе с помощью координатной сетки, относятся к ископаемым формам, среди которых серия родов ископаемых лошадей, Archaeopteryx, вымершие носороги и плечевой пояс молодой и взрослой особей плезиозавра.

Рисунок 24. Плечевой пояс плезиозавра Cryptocleidus в молодом (a) и взрослом (b) возрасте. Из книги Д’Арси Томпсона «О росте и форме» (1917).
Геометрический подход Томпсона заслужил много внимания и похвал, и повлиял на другие области знаний.[57] Но его широкое применение надолго задержалось. Методом стали интенсивно пользоваться лишь в последние десятилетия, когда были разработаны подходящие алгоритмы для работы с комплексной информацией геометрического плана. Начиная с 1980-х годов пережила свой расцвет дисциплина геометрической морфометрии, и были разработаны изощрённые методы работы и компьютерные программы, позволяющие сравнивать разные виды и прослеживать изменения, происходящие в процессе роста; в случае позвоночных большинство из них касалось их черепов. При геометрическом подходе фиксируется много точек-ориентиров и проводится количественная оценка того, как весь их массив различается от вида к виду или между стадиями в серии образцов, иллюстрирующих рост. Существуют различные подходы, основанные на использовании точек-ориентиров; у каждого из них своя собственная математическая структура и они включают методы, основанные на измерении деформации, суперпозиции и линейных расстояний.[58]
Альтернативный и более простой математический подход по сравнению с геометрическим подходом Томпсона был разработан примерно в то же самое время. Этот подход также оказал значительное влияние на исследования роста. В 1936 году Джулиан Хаксли и Жорж Тессье опубликовали одновременно на английском и французском языках работу, которая ввела в оборот простое, но изящное уравнение, которое суммирует отношение между двумя измеряемыми величинами. Это отношение было выражено как
y = xa
или в логарифмической форме,
log y = a log x,
где a — величина, вычисляемая с помощью отношения.
Математический график роста, или онтогенетическая траектория, после логарифмического преобразования обычно представляется в виде прямой линии. Направление или наклон и положение или пересечение с вертикальной осью графика — переменные величины, которые нужно вычислять.
Параметры, которые относятся к исследованию роста — это скорость, продолжительность, начало и завершение. Эволюция включает прерывание, изменения и перестановки этих параметров. Рост может порождать изменение формы, и это вызвано изменениями размеров, которые не находятся в отношении 1:1 с формой. Если отношение размеров к форме постоянно, мы имеем дело с изометрией. Это довольно редкий случай, поскольку мы гораздо чаще сталкиваемся с изменением формы, зависящим от изменения размеров, или аллометрией; этот термин Хаксли и Тессье использовали в своей статье 1936 года.[59] Из-за аллометрических отношений пропорций в процессе роста особи одного и того же вида могут иметь различную форму тела на разных стадиях роста. Из этого также следует, что внешний облик видов может меняться, если они обладают различными видами аллометрических отношений в процессе роста, или же если в процессе роста имеет место один и тот же вид отношений, но рост тормозится, или, напротив, продолжается дольше. В онтогенетическом шкалировании различия между двумя видами могут быть целиком отнесены к различиям в размерах, если они попадают на одну общую для обоих видов траекторию роста. Здесь различия являются следствием продления времени роста, или же его остановки. И наоборот, можно сделать вывод об изменениях в характере роста, если изменчивость не вписывается в одну общую траекторию.
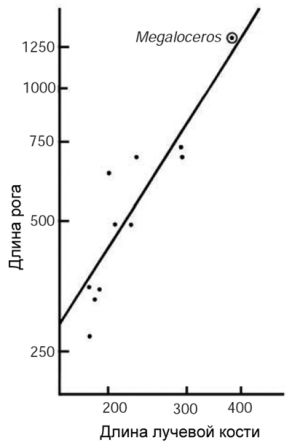
Рисунок 25. Классический пример исследования аллометрического роста, призванного помочь понять различные пропорции тела у вида — это исследование вымершего большерогого оленя Megaloceros giganteus, обладателя рогов, которые для нас выглядят огромными. Построив график отношения размеров тела к размерам рогов у разных видов оленей, Гоулд (1974) продемонстрировал, что большерогий олень вписывается в рамки той же самой онтогенетической траектории, что и другие виды. Таким образом, размер рогов этого «гиганта» фактически является ожидаемым для оленя такого размер. Длина рогов соотносится здесь с длиной одной из костей предплечья.
У математического подхода, разработанного Хаксли и Тессье, было несколько предшественников в конце девятнадцатого века, но лишь в двадцатом веке появилось сообщество биологов, готовых освоить и использовать новый числовой инструмент.[60] Сразу же после публикации уравнения роста, которое изящно выражало отношение между двумя размерными величинами, в этом ключе была исследована эволюция вымерших видов. Аллометрический подход был с энтузиазмом принят сообществом палеонтологов, и результатом этого стали многочисленные исследования эволюционных рядов тех таксонов, у которых приобретение какой-то специфической особенности выглядело результатом аллометрического роста — например, рога у ореодонтов (вымерших родственников верблюда), длина ростральной области у лошадей с эоцена до наших дней, гребень на голове у позднемелового динозавра Protoceratops и удлинённые верхние остистые отростки позвонков, образующие «парус» у пеликозавров, очень дальних, ранних родственников млекопитающих из позднего карбона и ранней перми Техаса. По итогам тех исследований было объявлено, что этими особенностями управляют простые взаимосвязи и прямые и постоянные аллометрические соотношения. Проведённые позже ревизии многих из этих исследований показали иное. Когда было обнаружено больше ископаемых остатков и были построены надёжно документированные эволюционные древа для этих групп организмов, стало понятно, что сама аллометрия также эволюционирует, что организмы представляли собой мозаику особенностей строения, растущих с различной скоростью, и что эти изменения находились под управлением экологических факторов.
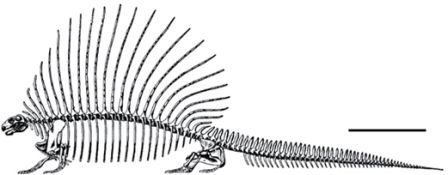
Рисунок 26. «Пеликозавр» Edaphosaurus boanerges. По Romer и Price, 1940 и Sander, 1994, с изменениями.
Хороший пример такого пересмотра прежних взглядов — эволюция рогов в филогенетическом ряду бронтотериев, вымерших дальних родственников лошадей и тапиров. Наросты на черепе у бронтотериев демонстрируют положительную аллометрию, как и в случаях других групп вымерших позвоночных. Эта разница в росте приводит к изменению формы; отклонения траекторий роста от изометрии ведут к значительным изменениям размеров, что может давать начало новым типам морфологии. Постепенное увеличение рогов с течением геологического времени изначально интерпретировалось как простое продолжение обычной траектории роста.
Более новые исследования показали, что в этом процессе были задействованы и другие изменения, такие, как более раннее начало развития и увеличение скорости роста. Эти изменения в траекториях роста могут быть ответом на более массивное телосложение и увеличение усилий, прикладывавшихся к рогам.
Классический пример эволюции аллометрических пропорций — это лошади, безрогие родственники бронтотериев. Эволюция черепа у лошадей включала появление новых типов процесса роста, связанных с новым рационом. Они хорошо документированы в виде сложного древа с многочисленными «боковыми ветвями», большинство из которых ведёт к вымершим видам, а другие ведут к видам, близкородственным современной лошади. Единственный современный род Equus — последний уцелевший представитель весьма разнообразной адаптивной радиации, протекавшей на протяжении последних 55 миллионов лет, итогом которой были примерно три дюжины вымерших родов и нескольких сотен вымерших видов. Около 20 миллионов лет назад лошади претерпели взрывоподобный рост разнообразия морфологии зубов. В процессе эволюции у многих родословных ветвей образовались высококоронковые зубы, подходящие для поедания травы на обширных травянистых равнинах большого количества открытых местообитаний, которые распространились в то время. К тому моменту геологического времени оцениваемый размер тела лошади находился в промежутке между ~5 и 50 килограммами. Напротив, в последние 20 миллионов лет лошади были более разнообразными в плане размеров тела. Некоторые группы стали крупными, другие сохранили относительно постоянный размере тела, а третьи со временем стали мельче. Из-за этих отклоняющихся эволюционных путей картина изменений выявленных закономерностей роста в эволюции лошадей выглядит не линейной, а разнообразной и ветвистой.[61] Полагают, что изменения в пропорциях черепа лошади вызваны изменениями аллометрического роста. Эти пропорции у взрослых особей ископаемых видов различны и связаны с функциями: в частности, это изменения в сторону высококоронковых зубов, сильнее сдвинутых вперёд относительно челюстного сустава, и более крупные челюсти для увеличения площади прикрепления мускулов. Аллометрические отношения между изменчивыми характеристиками черепа, наблюдаемые в ходе индивидуального роста ныне живущей лошади не идентичны тем, которые были выявлены путём сравнения черепов взрослых экземпляров различных ископаемых видов.
Вне зависимости от конкретной тенденции и от изучаемой группы позвоночных, будь то лошади, бронтотерии, бегемоты, рептилии, амфибии или рыбы, у них у всех имеются некоторые общие особенности роста. Та часть головы, где расположены мозг и органы чувств, в процессе роста демонстрирует отрицательную аллометрию: они меньше увеличиваются в размерах по сравнению с телом по мере роста животного. Морда, часть анатомии, связанная с челюстями, растёт, демонстрируя положительную аллометрию и становясь крупнее по отношению к увеличению размеров тела в процессе роста. Эта общая особенность справедлива также в отношении людей; например, младенцы имеют относительно большие глаза и маленький рот.
На уровне популяций в пределах вида аллометрический рост также может приводить к появлению морфологического разнообразия, как среди пород собак. Псовые в целом, группа, к которой принадлежат не только собаки, но также лисы и волки, характеризуются явно выраженным аллометрическим ростом черепа. В противоположность им кошачьи демонстрируют изометрию черепа и, соответственно, породы кошек морфологически менее разнообразны. То, что изменчиво у одной группы, может быть постоянным у другой.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК