Информативные срезы ископаемых костей
Причина того, что гистология ископаемой кости является богатым источником информации, состоит в том, что кость — это живая ткань, которая претерпевает много последовательных процессов резорбции и реконструкции. Этот факт приносит нам пользу после того, как с нами приключается несчастный случай, при котором мы ломаем кость; мы страдаем от этого в пожилом возрасте из-за остеопороза. Для палеонтологов изменение кости в течение жизни означает, что многое из этой записи жизни позвоночного животного можно увидеть. Ключевой момент здесь — умение различать следы первичного роста кости и процессов вторичной перестройки среди отметок, образовавшихся в процессе роста.
Самую ненарушенную информацию о росте скелета позвоночного обычно получают из района средней части длинных костей, таких, как в руках (плечевая кость) или в ногах (бедренная кость). Перед изготовлением срезов кости погружаются в синтетическую смолу, затем они шлифуются и полируются, пока не достигают соответствующей толщины, обычно 0,6–0,8 см. Далее эти срезы изучаются под микроскопом при различном освещении и с разными светофильтрами. Обычно предметом исследований палеонтологов являются отдельные обломки костей, поскольку музейные кураторы обычно не любят агрессивного способа исследований, который включает разрезание ископаемых образцов на тонкие кусочки. Это приводит к проблемам, поскольку зачастую нет уверенности в том, что известно точное с позиции анатомии или таксономии место взятия образца. Несмотря на эту трудность, количество новых открытий, сделанных путём исследований тонких срезов окаменелостей, увеличивается экспоненциально.
Моё первое знакомство с палеогистологией произошло благодаря моему более молодому коллеге Торстену Шейеру, который работал в моей лаборатории вскоре после моего прибытия в Цюрихский Университет, и благодаря его бывшему научному руководителю работы на соискание степени доктора философии Мартину Сандеру из Бонна, мировому лидеру в этой области. Я посетил лекцию Сандера в Тюбингене, посвящённую зауроподам, крупнейшим среди динозавров, на примере которых он показал, как много может дать палеогистология в понимании характера роста и устройства костей у этих животных. Мне пришло в голову, что этот подход можно было бы использовать для исследования Stupendemys geographicus, самой крупной водной черепахи из когда-либо существовавших, представителя гондванской группы, остатки которой мы собрали в ходе моего полевого проекта в Венесуэле. Сандер направил меня к Шейеру, и мы исследовали Stupendemys и других животных, главным образом млекопитающих и морских рептилий, о чём я и расскажу далее.
Панцирь Stupendemys принадлежит к числу самых толстых и самых крупных среди известных панцирей черепах; у одного экземпляра он достигает 3,3 м в длину. С точки зрения гистологии он демонстрирует хорошо выраженную слоистую, словно бутерброд, структуру, где внешние слои, включающие плотную кость, устойчивую к напряжению, окружают ячеистую внутреннюю губчатую кость. Такой тип структуры был важен для уменьшения веса панциря при сохранении прочности в процессе роста. Совокупность отметок, характеризующих рост, была неполной из-за эрозии, которой, несомненно, подверглись образцы, изучавшиеся нами, и которая характеризует местонахождение окаменелостей Урумако в Венесуэле.[40] Однако, вид ткани и просветы между отметками, образовавшимися в процессе роста, сопоставимы с таковыми у ныне живущих черепах с «нормальным» медленным темпом роста. Если предположить темп роста подобным таковому у большинства современных морских черепах с сопоставимыми гистологическими характеристиками, то экземпляр, который мы изучили, должно быть, рос примерно от 60 до 110 лет, прежде чем достиг гигантского размера панциря.
Организация ткани, наблюдаемая у современных животных, служит ключом для понимания особенностей гистологии, которые можно наблюдать в ископаемых костях. Остеобласты — это клетки, создающие кость. Также существуют пустоты, называемые лакунами, которые связаны друг с другом посредством канальцев, ответственных за распределение питательных веществ и кислорода. Эти и другие основные особенности строения кости могут наблюдаться в окаменелостях. Остеобласты и остеоциты, кровеносные сосуды или васкулярные каналы и коллагеновые волокна в большинстве случаев разрушаются в ходе фоссилизации, но их положение и форма могут быть зафиксированы в полной мере.[41] Некоторые исследования ныне живущих позвоночных показали, что костные ткани у животных, которые растут быстро, обладают иными особенностями в сравнении с тканями тех, кто растё медленно. Быстрый рост связан с костной тканью, богатой кровеносными сосудами, означающей усиленное снабжение кости кислородом и питательными веществами. Медленный рост коррелирует с менее васкуляризированной костью, часто прерываемой концентрическими линиями, сходными с годичными кольцами дерева. Это открытие — то, что скорость роста влияет на тип откладываемой костной ткани — было опубликовано Родолфо Амприно из Туринского Университета в 1947 году и сейчас известно как правило Амприно. Изучение гистологии костей было продолжено после выхода статьи Амприно, и палеонтологи следовали ходу его мыслей, пытаясь реконструировать аспекты физиологии и истории жизни ископаемых видов.

Рисунок 17. Тонкий срез фрагмента щитка Stupendemys geographicus из позднемиоценовой формации Урумако, Венесуэла. Образец находится в коллекции Университета Франсиско де Миранда в Коро (UNEFM-CIAPP-2002-01).
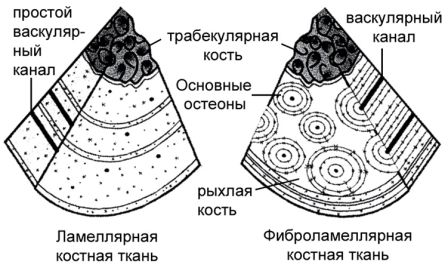
Рисунок 18. Схематическое представление двух основных видов гистологии костей. Они обычно отмечаются для эктотермных «рептилий» (слева) и эндотермных птиц и млекопитающих (справа). По Chiappe 2007, с изменениями.
В микроструктуре кости также существуют следы, которые предоставляют нам информацию о продолжительности жизни, возрасте и зрелости; это предмет изучения для скелетохронологии. Оценка возраста основывается на идентификации отметок, возникающих в процессе роста, таких, как кольца, образующиеся при замедлении роста, и линии задержек роста (ЛЗР) между зонами более быстрого роста кости. В большинстве случаев отметка, свидетельствующая об одном замедлении роста (кольцо) или его остановке, откладывается каждый год, во время более холодного, или же более сухого сезона, и это вызвано характером питания и физиологией животного наряду с переменными составляющими вроде болезни и репродуктивной цикличности. Из-за этих событий, происходящих в жизни отдельной особи, мы знаем, что образование маркеров роста может быть нерегулярным. Типичная картина отклонения — наличие короткого периода роста между годичным кольцом и ЛЗР, или когда за один цикл роста откладываются два кольца (плюс одна ЛЗР). Кроме минимального возраста, по микроструктуре костей можно дать оценку половой зрелости.
Уменьшение ширины зон быстрого роста и обычно изменение вида откладывающейся костной ткани может подразумевать достижение половой зрелости. В наилучших случаях может быть изучена ростовая серия образцов, и данные, полученные из различных элементов скелета, в противоположность единственному образцу, являются намного лучшим материалом для того, чтобы делать достоверные выводы о характере роста вымерших позвоночных. Для оценки минимального возраста животного в каждой кости подсчитываются циклы роста; для заключения о возрасте половой зрелости анализируется размер годичных слоёв. Когда в разных костях одной и той же особи наблюдается резорбция костной ткани, для оценки минимального возраста индивида используется кость с минимальным количеством резорбции и максимальным количеством циклов роста.
Разрушение и перестройка кости осуществляется клетками, которые носят название остеокласты.[42] Это интенсивный и энергоёмкий процесс; поэтому развитие кости тесно связано с метаболизмом. Это представляет непосредственный интерес для людей: самые последние исследования в области молекулярной медицины показали, что ожирение, диабет и остеопороз — это взаимосвязанные состояния.[43] В этом контексте были важны эксперименты с лабораторными мышами. Многое из того, что мы знаем о том, как растут и изменяются кости, получено прежде всего в ходе контрольных исследований, когда изучаются кости отдельно взятых животных, для которых исследователи знают пол, историю жизни, условия жизни и рацион. Например, у многих видов позвоночных беременность может привести к значительной перестройке костных тканей.[44] Это было зарегистрировано для многих видов рептилий, таких, как черепахи и крокодилы. В зависимости от группы источником кальция являются строго определённые кости, но не какие-то другие. Одной из них является малая берцовая кость, более тонкая из двух длинных костей, которая также присутствует в задней конечности и у нас, у людей. У некоторых видов крокодилов остеодермы у откладывающих яйца самок функционируют как хранилища кальция в ходе оогенеза. Эта стратегия, вероятно, характеризовала, по крайней мере, некоторых ископаемых рептилий, поэтому, как правило, использование остеодерм для целей скелетохронологии не является оптимальным решением, потому что в них происходит перестройка костного вещества. В случае с черепахами можно было бы подумать, что панцирь представляет собой превосходный источник материала, но пока изученные виды не демонстрируют использование самками этого источника. Возможно, панцирь просто слишком важен для обеспечения биомеханической целостности животного. Вместо него самки используют длинные кости вроде бедренной. Это явление в ходе оогенеза аналогично тому, что происходит у людей во время беременности и лактации, в периоды, характеризующиеся уменьшением плотности минеральной составляющей костей у женщин.
Птицы производят твёрдую скорлупу яиц, состоящую больше чем на 90 процентов из кристаллов карбоната кальция. У птиц перед овуляцией в полости костного мозга образуется особый вид эфемерной костной ткани, служащий источником кальция, который позволяет матери образовывать скорлупу яиц. Этот тип кости называется медуллярной костью, и теперь он описан для Tyrannosaurus rex и для других видов, представляющих основные группы динозавров. Это открытие стало важным не только для того, чтобы вывести происхождение птиц из динозавров, но также (что даже ещё важнее) для более точной реконструкции кривых роста для многих видов динозавров с использованием надёжного маркера состояния репродуктивной зрелости. У динозавров этот момент совпал с переходом от стадии ускорения роста к стадии замедления.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК