Учимся летать: от множества жабр к паре крыльев
Многофункциональность разветвленных конечностей водных членистоногих является ключом к пониманию двух важнейших новых способностей, приобретенных членистоногими, — ходить по земле и летать по воздуху. У водных ракообразных жаберные придатки, расположенные на верхних ветвях конечностей, служат для выполнения дыхательной функции, тогда как нижние ветви нужны для ходьбы или плавания. У всех наземных членистоногих ходильные ноги неразветвленные. Это упрощенное строение появилось в результате редуцирования сложных разветвленных конечностей предков к единственной нижней ветви.
Таким образом, мы разобрались, как у насекомых появились ноги, но откуда взялись крылья? Происхождение крыльев насекомых долгое время оставалось загадкой. Некоторые биологи считали, что крылья появились как независимые выросты стенки грудного отдела бескрылых насекомых. Другая гипотеза заключалась в том, что крылья могли эволюционировать из ветвей конечностей, в частности, из жабр обитавших в воде предков. Специалисты в области сравнительной анатомии долгое время бились над этими двумя гипотезами и никак не могли прийти к единому мнению.
И опять новые данные были получены с помощью эво-дево. Изучение развития крыльев насекомых, особенно дрозофил, позволило идентифицировать белки, необходимые для построения крыла. Два таких белка, являющихся продуктами генов развития, получили названия Apterous ("бескрылый"; при мутации соответствующего гена крылья отсутствуют) и Nubbin ("комочек"; при мутации соответствующего гена вместо крыльев появляются лишь небольшие утолщения ткани). Чтобы проверить гипотезу о происхождении крыльев из жаберных придатков конечностей ракообразных, Михалис Авероф и Стивен Коэн занялись анализом экспрессии Apterous и Nubbin в конечностях других членистоногих, особенно ракообразных. К большому удивлению, они обнаружили, что гены apterous и nubbin избирательно экспрессируются в жаберном придатке верхней ветви конечностей ракообразных. Лучшее объяснение этого открытия заключается в том, что жаберный придаток ракообразных и крыло насекомых являются гомологами —т.е. вариантами одной и той же части тела у разных видов животных. Единственным альтернативным объяснением могло быть невероятное совпадение, в результате которого для создания жабр и крыльев насекомые и ракообразные из сотен генов развития выбрали именно эти два. Гораздо более вероятно, что Apterous и Nubbin использовались для создания дыхательного аппарата у водных ракообразных, являвшихся предками насекомых, и продолжали свою работу даже после того, как у насекомых эти конечности превратились в крылья (и, как мы увидим позднее, в другие структуры у других животных). Затем у насекомых верхние и нижние ветви конечностей разделились: часть верхней ветви поднялась на спину и превратилась в крыло, а нижняя часть превратилась в неразветвленную ходильную ногу.
У теории превращения жабр в крылья всегда были доказательства (но, вероятно, они были недостаточно весомы, чтобы раз и навсегда решить этот вопрос). Если крылья насекомых образовались из жабр ракообразных, означает ли это, что какие-то раки или креветки просто выползли на землю и взлетели? Нет, совсем не так. Животных, приобретших дыхательный аппарат, и насекомых с пригодными для полета крыльями разделяет множество эволюционных стадий. Лучше всего понять, как произошел этот переход, помогают ископаемые остатки давно исчезнувших насекомых и открытия эво-дево. Наша лаборатория участвовала в изучении этого вопроса, и это еще один пример того, как сочетание новых знаний об ископаемых животных, генах и эмбрионах позволяет выстроить цельную картину событий прошлого.
Ископаемые, ставшие важными источниками информации о структуре древних насекомых, не похожи ни на одно современное животное. Меня в свое время очень удивил рисунок нимфы[8] примитивного водного насекомого" (рис. 7.5. слева). Это существо, жившее более 300 млн лет назад, независимо изучали палеонтологи Робин Вуттон и Ярмила Кукалова-Пек. Важнейшая особенность этого животного — наличие на всех грудных и брюшных сегментах структур, напоминающих крылья. Сходство с крыльями им придают канальцы, похожие на жилки на крыльях насекомых. Однако это существо было водным животным, и эти структуры — не крылья, а жабры, напоминающие жабры нимф стрекоз и поденок.
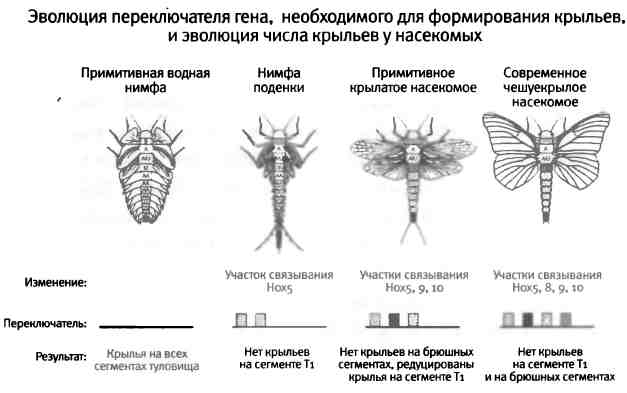
Рис. 7.5. Эволюция числа и формы крыльев. В соответствии с законом Уилли-стона, крылья возникли из ряда жаброподобных конечностей на всех сегментах нимфы вымершего членистоногого. Число этих выростов постепенно уменьшалось (нимфа поденки), и в какой-то момент они превратились в две пары крыльев, как у большинства современных крылатых насекомых. Постепенное сокращение числа крыльев связано с постепенным увеличением числа участков для связывания Hox-белков в переключателях генов, способствующих развитию крыла. Рисунок Лианн Олдс.
Скорее всего, крылья появились у тех наземных насекомых, которые на личиночной стадии имели жабры. В таком случае крыльями могли стать структуры взрослого животного, модифицированные из жабр, но это не означало отсутствия жабр у личинок. Поденки и стрекозы, самые примитивные из всех крылатых насекомых, проходят в своем жизненном цикле стадию водной нимфы с жабрами на брюшке. Наличие в жизненном цикле животных дискретных стадий открывает большие возможности для эволюции. Подумайте, ведь взрослая стрекоза или взрослая поденка имеют мало общего со своей водной молодью, обитающей в совершенно других условиях. Адаптация к этим разным условиям существования произошла одновременно, на базе одного генома, только за счет разделения программы развития нимфы и программы развития взрослого животного. Эволюция совершенно разной морфологии личиночной и взрослой формы — очень распространенная тенденция в мире животных (гусеницы и бабочки, билатерально симметричные личинки иглокожих и радиально симметричные взрослые формы).
Этот сценарий помогает объяснить превращение жабр в крылья, но не объясняет, почему изменилось число крыльев. Аэродинамические исследования показывают, что две пары крыльев, расположенные на втором и третьем грудных сегментах, обеспечивают наилучшие летные характеристики. Как насекомые пришли к этому оптимальному решению?
Возвращаемся к закону Уиллистона, Hox-генам и переключателям. Специализация часто сопровождается сокращением числа сериально гомологичных структур, помните? Именно это и произошло в процессе эволюции крыльев насекомых. В палеонтологической летописи присутствуют вымершие формы с меньшим количеством или меньшими по размеру крыльями на брюшке и на первом грудном сегменте (T1). Эти формы являются промежуточными между примитивными водными формами и современными крылатыми насекомыми (рис. 7.5). В процессе развития формирование крыльев на всех участках, кроме второго и третьего грудных сегментов, сокращалось, что привело к уменьшению количества крыльев.
За счет чего подавление развития крыльев осуществлялось столь избирательно? Сегменты, на которых крылья исчезли, у всех насекомых совпадают с зонами экспрессии определенных Hox-белков. Более того, Скотт Уэзерби и Джим Ленжленд из нашей лаборатории показали, что в этих сегментах у дрозофилы образование крыла подавляется за счет экспрессии соответствующих Hox-генов. Это говорит о том, что современные насекомые с двумя парами крыльев получились в результате подавления развития крыла Hox-белками, действующими в первом грудном и во всех брюшных сегментах. Этот механизм должен был эволюционировать постепенно в разных группах древних насекомых, поскольку палеонтологическая летопись содержит такие виды, у которых формирование "лишних" крыльев подавлено не полностью. Первопричина подавления формирования крыльев заключается в том, что в переключателях генов, вовлеченных в формирование крыльев, появились сигнатурные последовательности для Hox-белков, которые привели к отключению этих генов в соответствующих сегментах.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК